
Effect of L-Glutamine on Chylomicron Formation and Fat-Induced Activation of Intestinal Mucosal Mast Cells in Sprague-Dawley Rats


Abstract
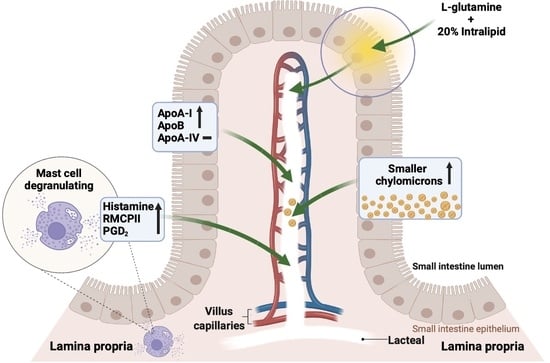
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Lymph Fistula Surgery with Duodenal Cannulation
2.3. Fat Absorption Study and Lymph Collection
2.4. Measurement of Lymphatic Lipid Outputs
2.5. Measurement of Apolipoproteins
2.6. Lipoprotein Particle Size Analysis by Negative Staining Electron Microscopy
2.7. Measurement of Rat Mucosal Mast Cell Protease II
2.8. Measurement of Lymphatic Histamine and Prostaglandin D2
2.9. Statistical Analysis
3. Results
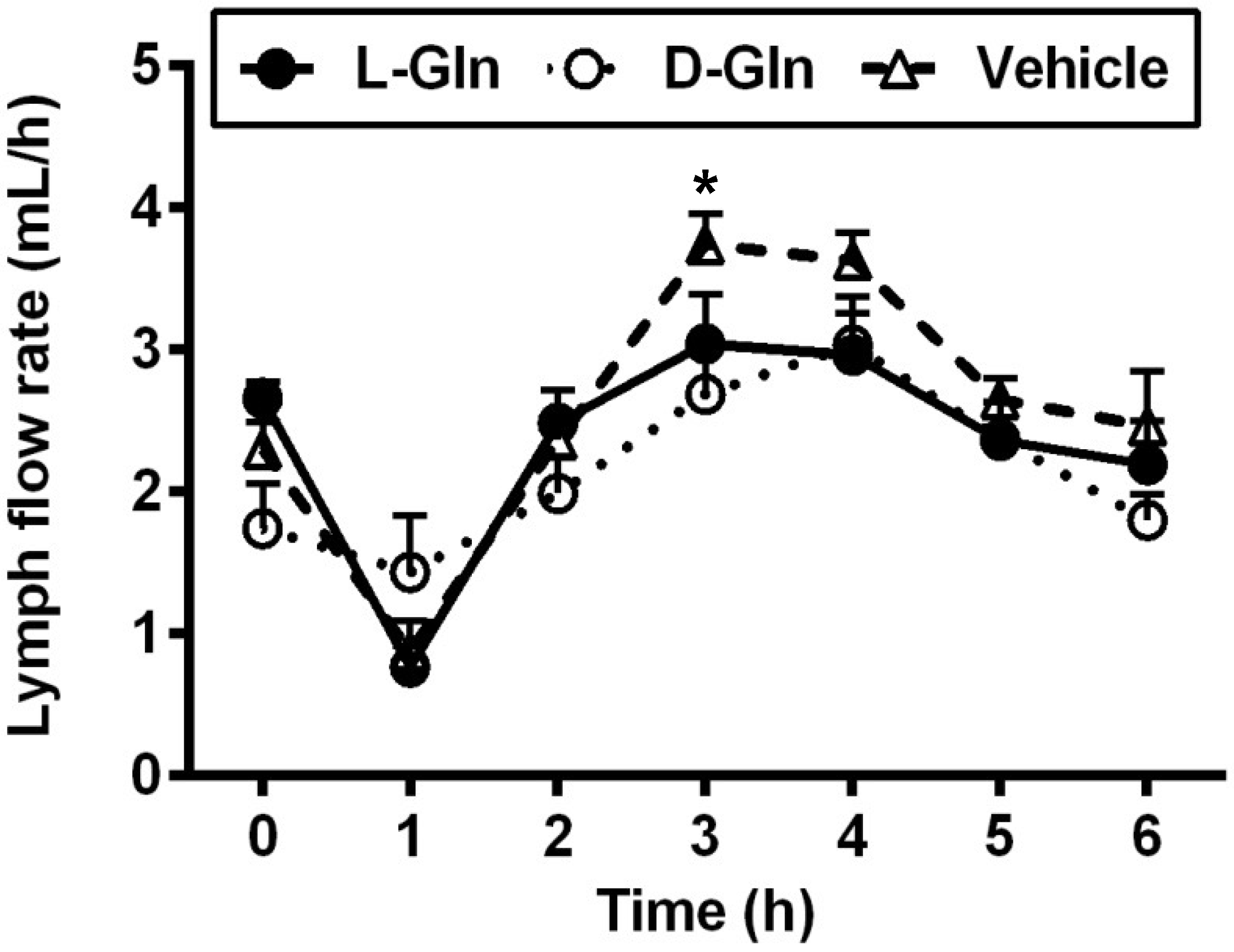
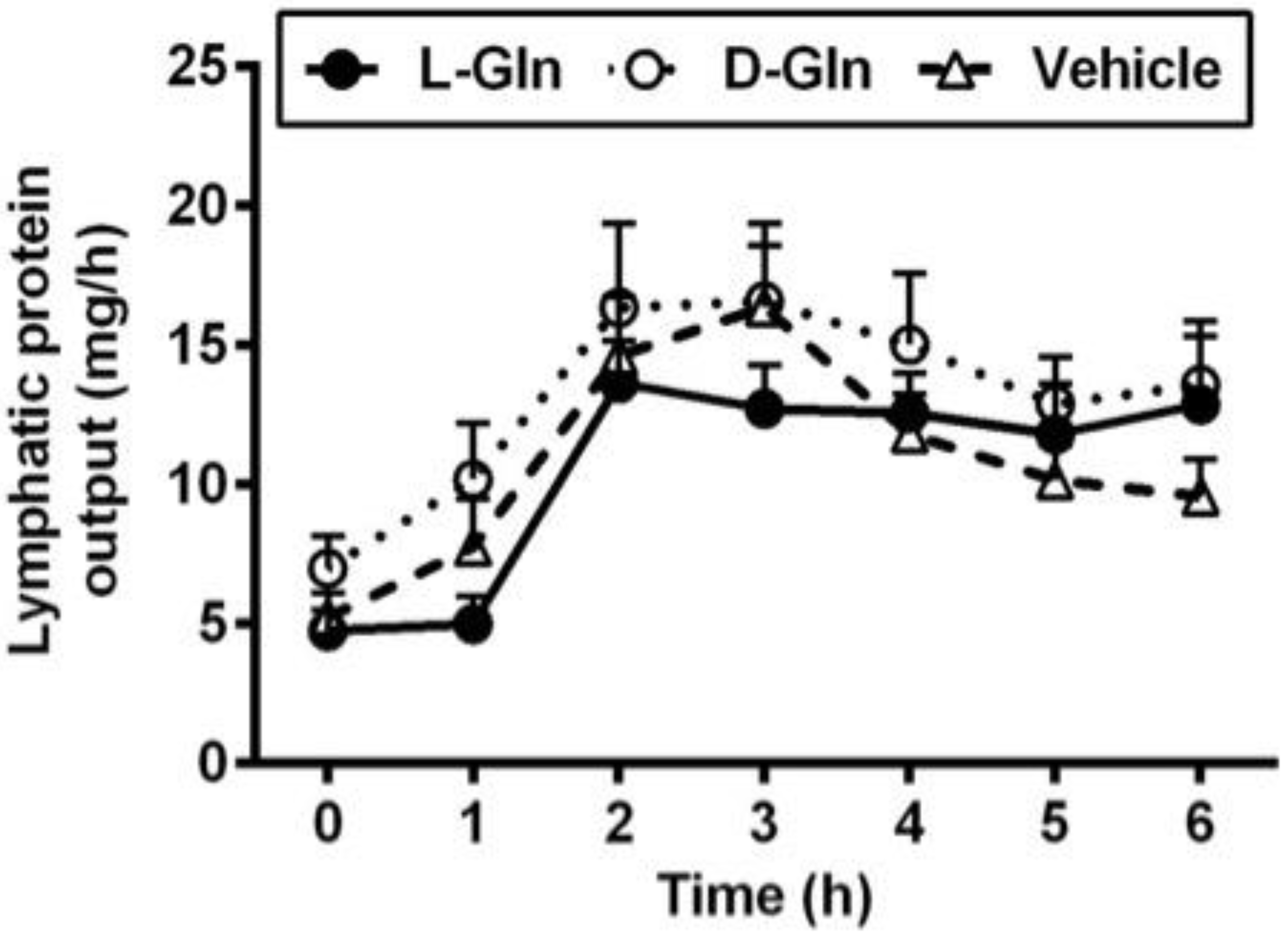
3.1. Lymph Flow Rate and Protein Output
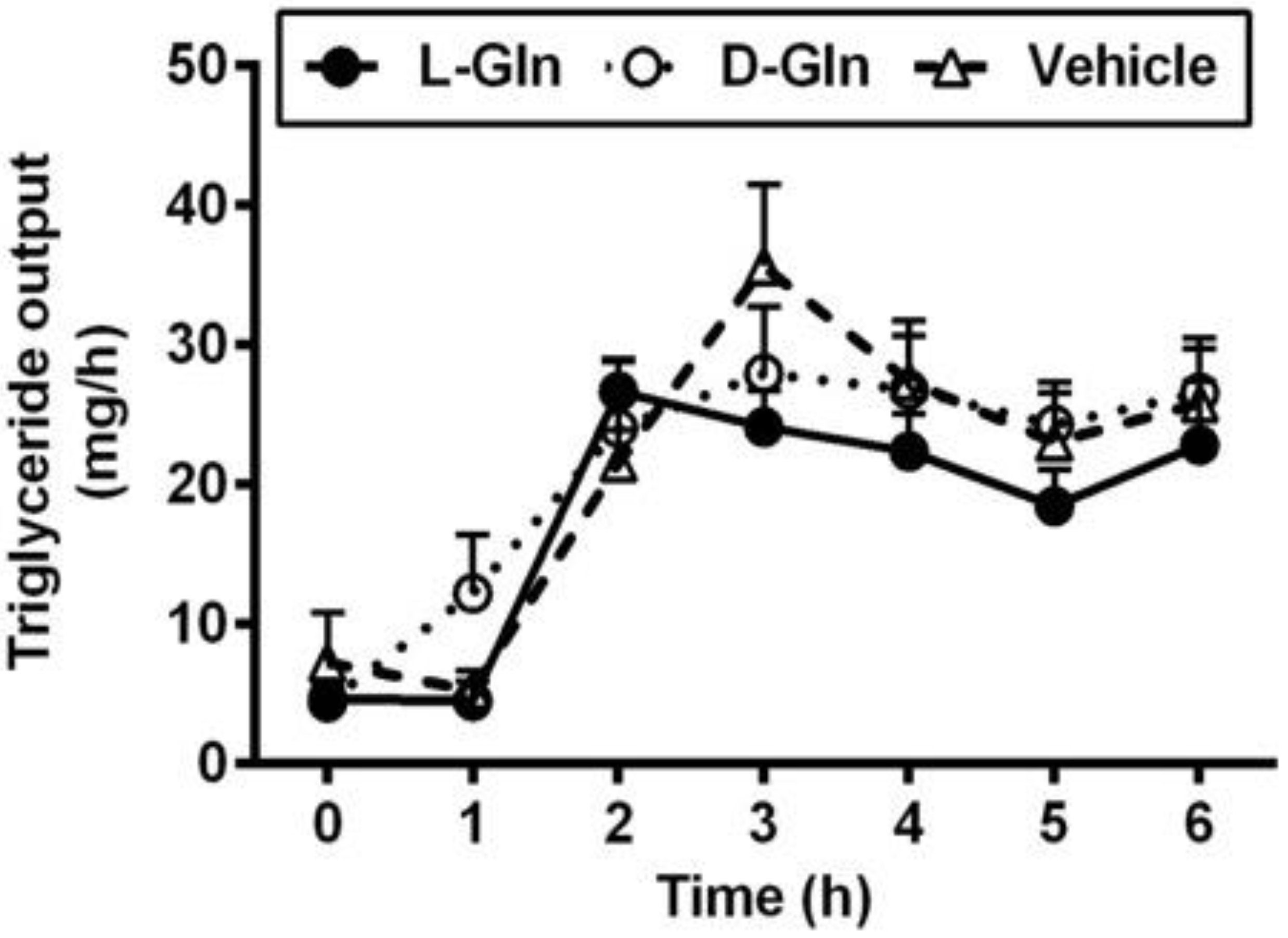
3.2. Lymphatic Outputs of Triglyceride and Phospholipid
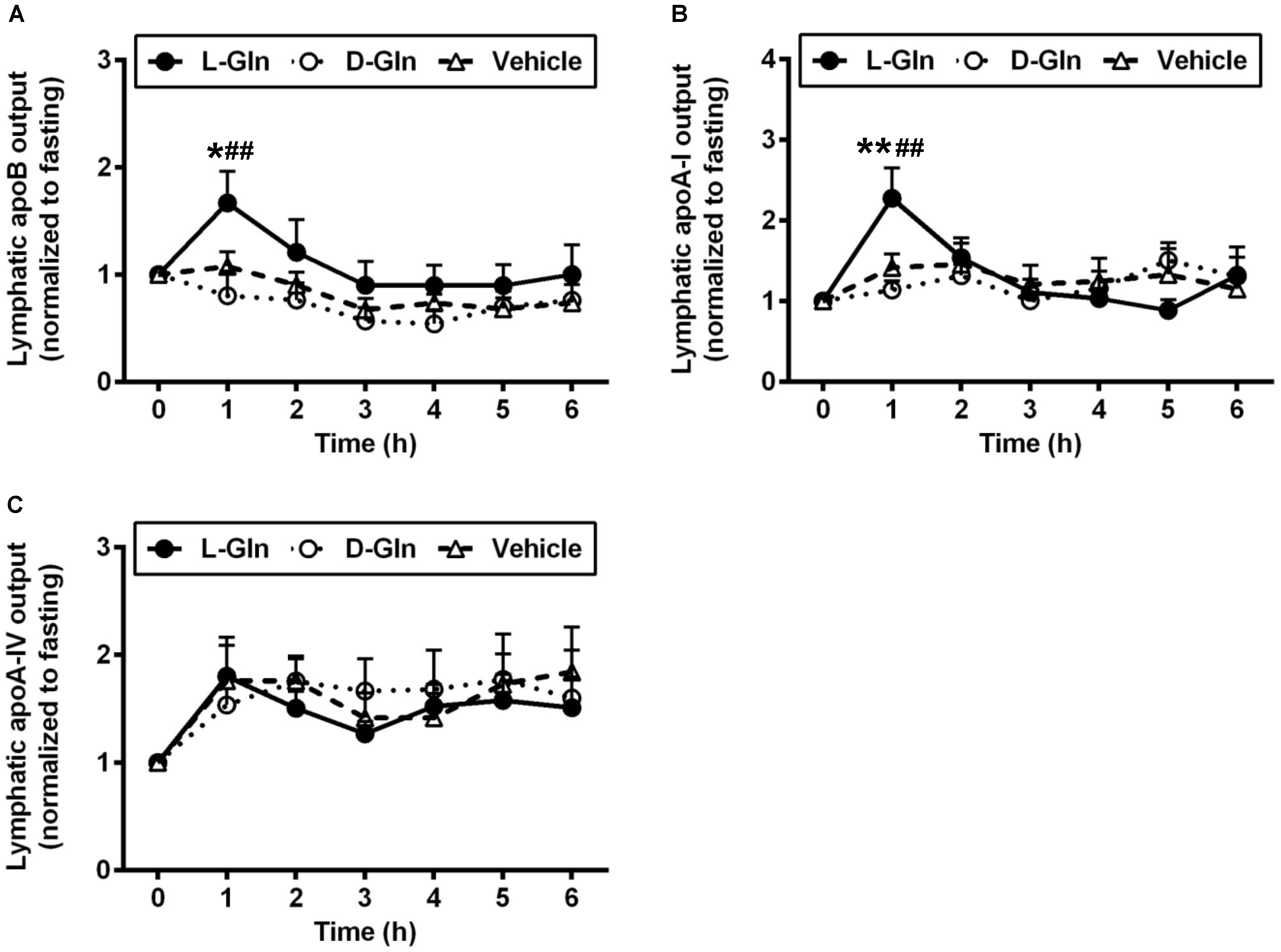
3.3. Lymphatic Outputs of Apolipoproteins
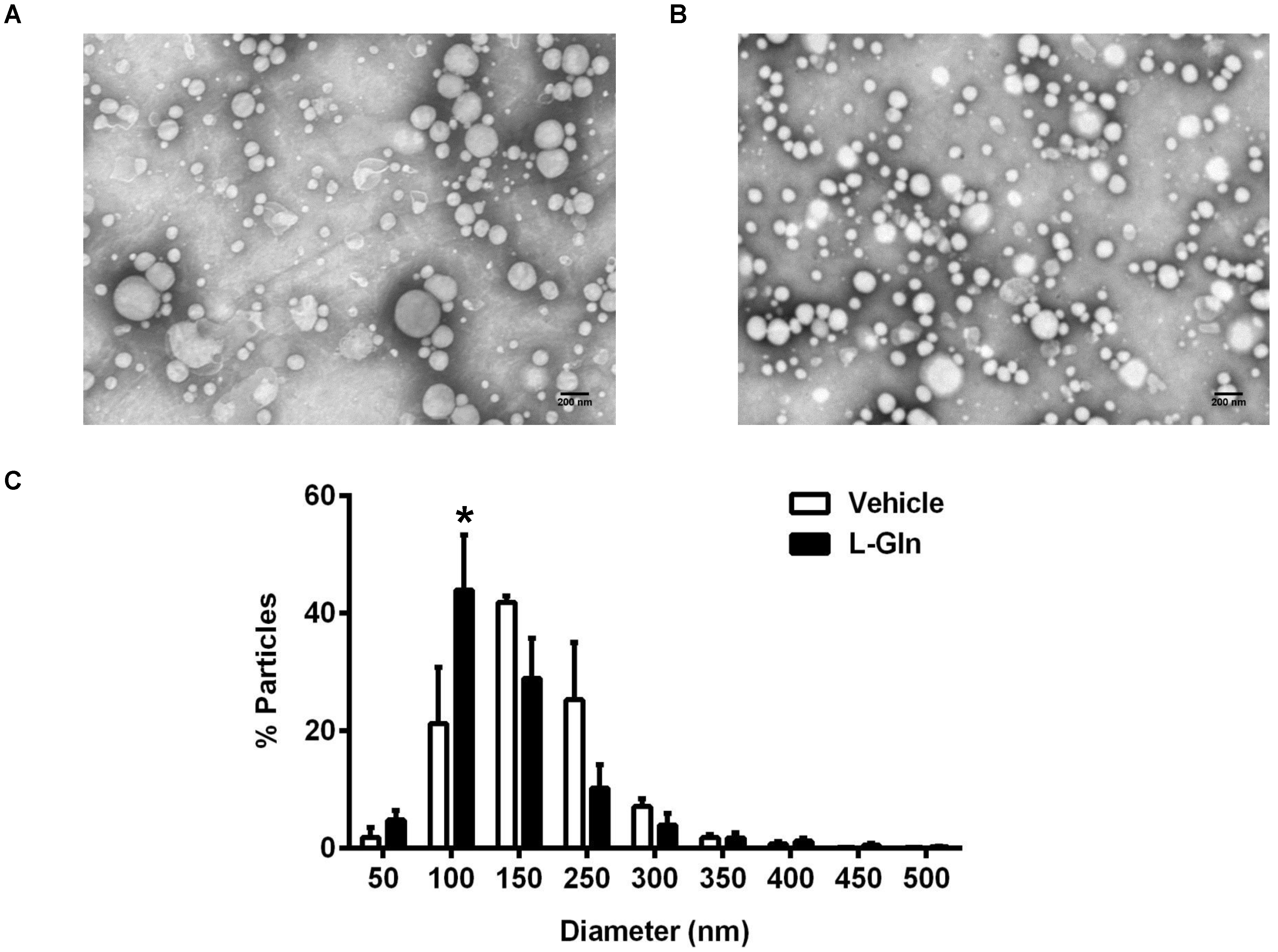
3.4. Size Distribution of Lipoproteins as Analyzed by Negative Staining Electron Microscopy
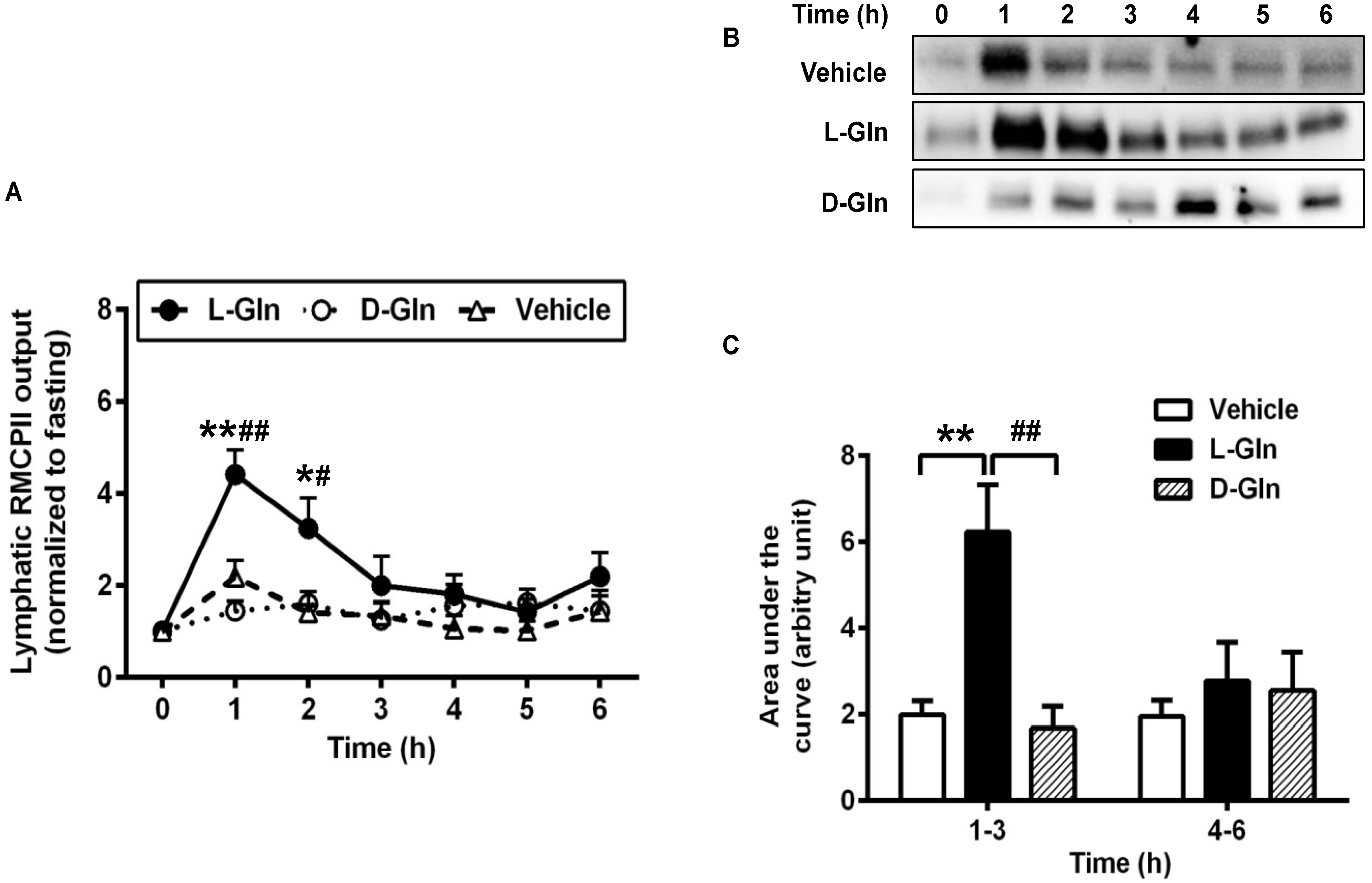
3.5. Activation of Intestinal Mucosal Mast Cells
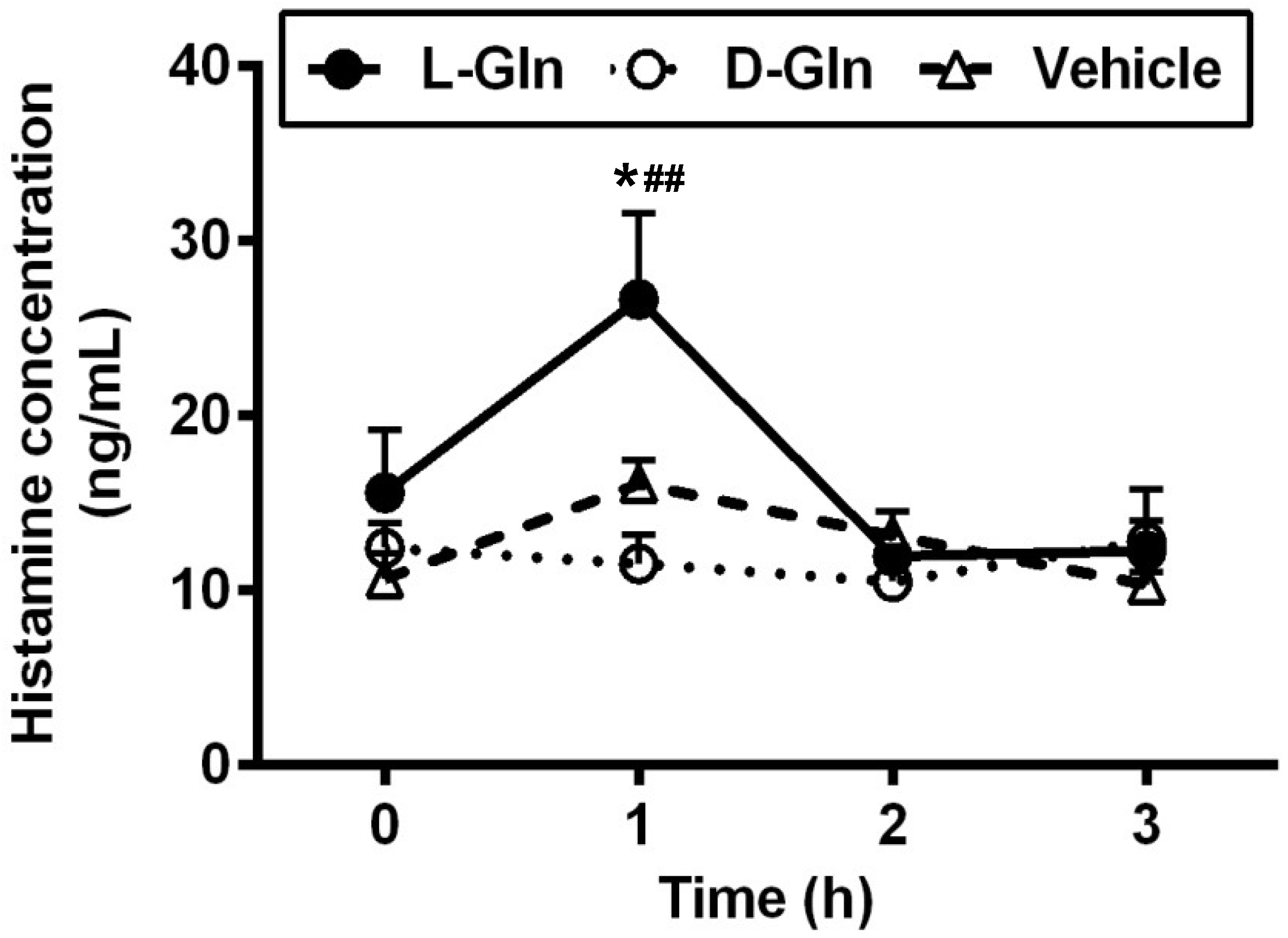
3.6. Lymphatic Histamine Secretion
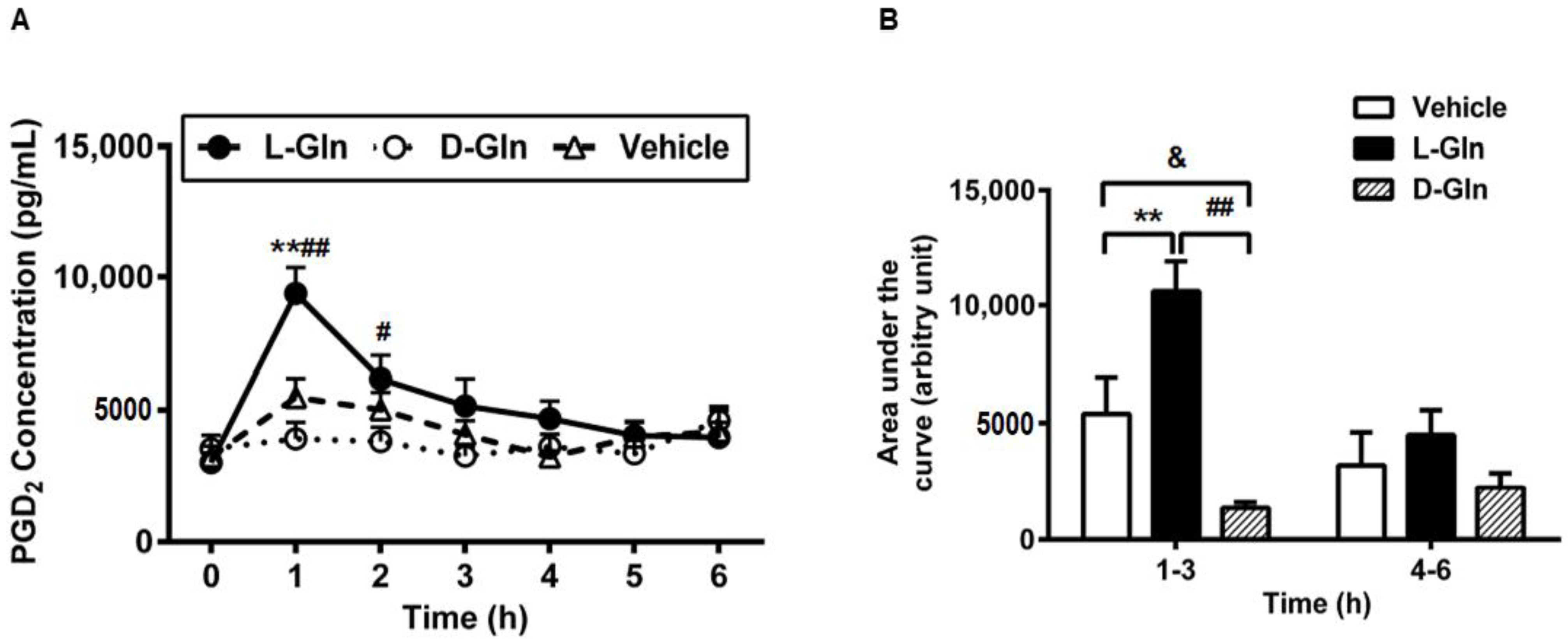
3.7. Lymphatic Prostaglandin D2 (PGD2) Secretion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Camilleri, M.; Madsen, K.; Spiller, R.; Greenwood-Van Meerveld, B.; Verne, G.N. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol. Motil. 2012, 24, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Y. Intestinal mucosal amino acid catabolism. J. Nutr. 1998, 128, 1249–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rescigno, M. The intestinal epithelial barrier in the control of homeostasis and immunity. Trends Immunol. 2011, 32, 256–264. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Albert-Bayo, M.; Paracuellos, I.; Gonzalez-Castro, A.M.; Rodriguez-Urrutia, A.; Rodriguez-Lagunas, M.J.; Alonso-Cotoner, C.; Santos, J.; Vicario, M. Intestinal Mucosal Mast Cells: Key Modulators of Barrier Function and Homeostasis. Cells 2019, 8, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, S.C. Physiological and pathophysiological functions of intestinal mast cells. Semin. Immunopathol. 2009, 31, 185–205. [Google Scholar] [CrossRef]
- Reber, L.L.; Sibilano, R.; Mukai, K.; Galli, S.J. Potential effector and immunoregulatory functions of mast cells in mucosal immunity. Mucosal Immunol. 2015, 8, 444–463. [Google Scholar] [CrossRef] [Green Version]
- Boyce, J.A. Mast cells and eicosanoid mediators: A system of reciprocal paracrine and autocrine regulation. Immunol. Rev. 2007, 217, 168–185. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Valenta, R.; Hochwallner, H.; Linhart, B.; Pahr, S. Food allergies: The basics. Gastroenterology 2015, 148, 1120–1131.e4. [Google Scholar] [CrossRef] [Green Version]
- Khodoun, M.; Strait, R.; Orekov, T.; Hogan, S.; Karasuyama, H.; Herbert, D.R.; Kohl, J.; Finkelman, F.D. Peanuts can contribute to anaphylactic shock by activating complement. J. Allergy Clin. Immunol. 2009, 123, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, H.A.; Munoz-Furlong, A.; Campbell, R.L.; Adkinson, N.F.; Bock, S.A.; Branum, A.; Brown, S.G.A.; Camargo, C.A.; Cydulka, R.; Galli, S.J.; et al. Second Symposium on the Definition and Management of Anaphylaxis: Summary report—Second National Institute of Allergy and Infectious Disease/Food Allergy and Anaphylaxis Network Symposium. Ann. Emerg. Med. 2006, 47, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Fields, J.Z.; Keshavarzian, A. Mucosal mast cells are pivotal elements in inflammatory bowel disease that connect the dots: Stress, intestinal hyperpermeability and inflammation. World J. Gastroenterol. 2007, 13, 3027–3030. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sharma, A. Mast cells: Emerging sentinel innate immune cells with diverse role in immunity. Mol. Immunol. 2010, 48, 14–25. [Google Scholar] [CrossRef]
- Ji, Y.; Sakata, Y.; Yang, Q.; Li, X.; Xu, M.; Yoder, S.; Langhans, W.; Tso, P. Activation of rat intestinal mucosal mast cells by fat absorption. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 302, G1292–G1300. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Zhang, L.S.; Martinez, K.; Chang, E.B.; Yang, Q.; Wang, F.; Howles, P.N.; Hokari, R.; Miura, S.; Tso, P. Antibiotics Suppress Activation of Intestinal Mucosal Mast Cells and Reduce Dietary Lipid Absorption in Sprague-Dawley Rats. Gastroenterology 2016, 151, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Knabe, D.A. Free and protein-bound amino acids in sow’s colostrum and milk. J. Nutr. 1994, 124, 415–424. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Knabe, D.A.; Burghardt, R.C.; Spencer, T.E.; Li, X.L.; Wang, J.J. Triennial Growth Symposium: Important roles for L-glutamine in swine nutrition and production. J. Anim. Sci. 2011, 89, 2017–2030. [Google Scholar] [CrossRef] [Green Version]
- Windmueller, H.G.; Spaeth, A.E. Uptake and metabolism of plasma glutamine by the small intestine. J. Biol. Chem. 1974, 249, 5070–5079. [Google Scholar] [CrossRef]
- Benjamin, J.; Makharia, G.; Ahuja, V.; Anand Rajan, K.D.; Kalaivani, M.; Gupta, S.D.; Joshi, Y.K. Glutamine and whey protein improve intestinal permeability and morphology in patients with Crohn’s disease: A randomized controlled trial. Dig. Dis. Sci. 2012, 57, 1000–1012. [Google Scholar] [CrossRef]
- Lechowski, S.; Feilhauer, K.; Staib, L.; Coeffier, M.; Bischoff, S.C.; Lorentz, A. Combined arginine and glutamine decrease release of de novo synthesized leukotrienes and expression of proinflammatory cytokines in activated human intestinal mast cells. Eur. J. Nutr. 2013, 52, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Reeds, P.J.; Burrin, D.G. Glutamine and the bowel. J. Nutr. 2001, 131, 2505S–2508S; discussion 2523S–2504S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, T.A.; Morrissey, T.B.; Nattakom, T.V.; Ziegler, T.R.; Wilmore, D.W. Growth hormone, glutamine, and a modified diet enhance nutrient absorption in patients with severe short bowel syndrome. J. Parenter. Enter. Nutr. 1995, 19, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Hatami, B.; Saffaei, A.; Jamali, F.; Abbasinazari, M. Glutamine powder-induced hepatotoxicity: It is time to understand the side effects of sports nutritional supplements. Gastroenterol. Hepatol. Bed Bench 2020, 13, 86–89. [Google Scholar]
- Holecek, M. Side effects of long-term glutamine supplementation. J. Parenter. Enter. Nutr. 2013, 37, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Holecek, M. Adverse effects of chronic intake of glutamine-supplemented diet on amino acid concentrations and protein metabolism in rat: Effect of short-term starvation. e-SPEN 2011, 6, e190–e196. [Google Scholar] [CrossRef] [Green Version]
- Schwimmer, J.B.; Ee, L.; Zheng, S.; Tso, P. Glutamine promotes triglyceride absorption in a dose-dependent manner. Am. J. Physiol.-Gastrointest. Liver Physiol. 2002, 282, G317–G323. [Google Scholar] [CrossRef]
- D’Alessio, D.; Lu, W.; Sun, W.; Zheng, S.; Yang, Q.; Seeley, R.; Woods, S.C.; Tso, P. Fasting and postprandial concentrations of GLP-1 in intestinal lymph and portal plasma: Evidence for selective release of GLP-1 in the lymph system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R2163–R2169. [Google Scholar] [CrossRef]
- Bollman, J.L.; Cain, J.C.; Grindlay, J.H. Techniques for the collection of lymph from the liver, small intestine, or thoracic duct of the rat. J. Lab. Clin. Med. 1948, 33, 1349–1352. [Google Scholar]
- Austgen, T.R.; Dudrick, P.S.; Sitren, H.; Bland, K.I.; Copeland, E.; Souba, W.W. The effects of glutamine-enriched total parenteral nutrition on tumor growth and host tissues. Ann. Surg. 1992, 215, 107–113. [Google Scholar] [CrossRef]
- Bruckner, H.; Westhauser, T. Chromatographic Determination of D-Amino Acids as Native Constituents of Vegetables and Fruits. Chromatographia 1994, 39, 419–426. [Google Scholar] [CrossRef]
- Hayashi, H.; Nutting, D.F.; Fujimoto, K.; Cardelli, J.A.; Black, D.; Tso, P. Transport of lipid and apolipoproteins A-I and A-IV in intestinal lymph of the rat. J. Lipid Res. 1990, 31, 1613–1625. [Google Scholar] [CrossRef]
- Tso, P.; Drake, D.S.; Black, D.D.; Sabesin, S.M. Evidence for Separate Pathways of Chylomicron and Very Low-Density Lipoprotein Assembly and Transport by Rat Small-Intestine. Am. J. Physiol. 1984, 247, G599–G610. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.M.; Nordskog, B.K.; Nauli, A.M.; Zheng, S.Q.; Vonlehmden, S.B.; Yang, Q.; Lee, D.; Swift, L.L.; Davidson, N.O.; Tso, P. Why does the gut choose apolipoprotein B48 but not B100 for chylomicron formation? Am. J. Physiol.-Gastrointest. Liver Physiol. 2008, 294, G344–G352. [Google Scholar] [CrossRef] [Green Version]
- Aberdeen, V.; Shepherd, P.A.; Simmonds, W.J. Concurrent measurement, in unanaesthetized rats, of intestinal transport and fat absorption from the lumen. Q. J. Exp. Physiol. Cogn. Med. Sci. Transl. Integr. 1960, 45, 265–274. [Google Scholar] [CrossRef]
- Tso, P.; Balint, J.A. Formation and transport of chylomicrons by enterocytes to the lymphatics. Am. J. Physiol. 1986, 250, G715–G726. [Google Scholar] [CrossRef]
- Ockner, R.K.; Hughes, F.B.; Isselbacher, K.J. Very Low Density Lipoproteins in Intestinal Lymph—Origin, Composition, and Role in Lipid Transport in Fasting State. J. Clin. Investig. 1969, 48, 2079–2088. [Google Scholar] [CrossRef]
- Gibson, S.; Mackeller, A.; Newlands, G.F.; Miller, H.R. Phenotypic expression of mast cell granule proteinases. Distribution of mast cell proteinases I and II in the rat digestive system. Immunology 1987, 62, 621–627. [Google Scholar]
- Beaugerie, L.; Carbonnel, F.; Hecketsweiler, B.; Dechelotte, P.; Gendre, J.P.; Cosnes, J. Effects of an isotonic oral rehydration solution, enriched with glutamine, on fluid and sodium absorption in patients with a short-bowel. Aliment. Pharm. Therap. 1997, 11, 741–746. [Google Scholar] [CrossRef]
- Scolapio, J.S.; Camilleri, M.; Fleming, C.R.; Oenning, L.V.; Burton, D.D.; Sebo, T.J.; Batts, K.P.; Kelly, D.G. Effect of growth hormone, glutamine, and diet on adaptation in short-bowel syndrome: A randomized, controlled study. Gastroenterology 1997, 113, 1074–1081. [Google Scholar] [CrossRef]
- Zhou, Q.; Verne, M.L.; Fields, J.Z.; Lefante, J.J.; Basra, S.; Salameh, H.; Verne, G.N. Randomised placebo-controlled trial of dietary glutamine supplements for postinfectious irritable bowel syndrome. Gut 2019, 68, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Jin, M.; Lee, E.; Moon, T.C.; Quan, Z.; Yang, J.H.; Son, K.H.; Kim, K.U.; Son, J.K.; Chang, H.W. Effects of methyl gallate on arachidonic acid metabolizing enzymes: Cyclooxygenase-2 and 5-lipoxygenase in mouse bone marrow-derived mast cells. Arch. Pharmacal. Res. 2006, 29, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Sniderman, A.D.; Thanassoulis, G.; Glavinovic, T.; Navar, A.M.; Pencina, M.; Catapano, A.; Ference, B.A. Apolipoprotein B Particles and Cardiovascular Disease: A Narrative Review. JAMA Cardiol. 2019, 4, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.W.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Kohan, A.B.; Howles, P.N.; Tso, P. Methods for studying rodent intestinal lipoprotein production and metabolism. Curr. Protoc. Mouse Biol. 2012, 2, 219–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Fujimoto, K.; Cardelli, J.A.; Nutting, D.F.; Bergstedt, S.; Tso, P. Fat feeding increases size, but not number, of chylomicrons produced by small intestine. Am. J. Physiol. 1990, 259, G709–G719. [Google Scholar] [CrossRef]
- Kindel, T.; Lee, D.M.; Tso, P. The mechanism of the formation and secretion of chylomicrons. Atheroscler. Suppl. 2010, 11, 11–16. [Google Scholar] [CrossRef]
- Ji, Y.; Sakata, Y.; Li, X.; Zhang, C.; Yang, Q.; Xu, M.; Wollin, A.; Langhans, W.; Tso, P. Lymphatic diamine oxidase secretion stimulated by fat absorption is linked with histamine release. Am. J. Physiol.-Gastrointest. Liver Physiol. 2013, 304, G732–G740. [Google Scholar] [CrossRef] [Green Version]
- Julve, J.; Martin-Campos, J.M.; Escola-Gil, J.C.; Blanco-Vaca, F. Chylomicrons: Advances in biology, pathology, laboratory testing, and therapeutics. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 455, 134–148. [Google Scholar] [CrossRef]
- Weinberg, R.B.; Gallagher, J.W.; Fabritius, M.A.; Shelness, G.S. ApoA-IV modulates the secretory trafficking of apoB and the size of triglyceride-rich lipoproteins. J. Lipid Res. 2012, 53, 736–743. [Google Scholar] [CrossRef] [Green Version]
- Albers, J.J.; Kennedy, H.; Marcovina, S.M. Evidence that Lp[a] contains one molecule of apo[a] and one molecule of apoB: Evaluation of amino acid analysis data. J. Lipid Res. 1996, 37, 192–196. [Google Scholar] [CrossRef]
- Uarfordt, S.H.; Goodman, D.S. Chylomicron cholesteryl ester metabolism in the perfused rat liver. Biochim. Biophys. Acta 1969, 176, 863–872. [Google Scholar] [CrossRef]
- Higashiguchi, T.; Hasselgren, P.O.; Wagner, K.; Fischer, J.E. Effect of glutamine on protein synthesis in isolated intestinal epithelial cells. J. Parenter. Enteral. Nutr. 1993, 17, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Curi, R.; Lagranha, C.J.; Doi, S.Q.; Sellitti, D.F.; Procopio, J.; Pithon-Curi, T.C.; Corless, M.; Newsholme, P. Molecular mechanisms of glutamine action. J. Cell Physiol. 2005, 204, 392–401. [Google Scholar] [CrossRef]
- Keita, A.V.; Soderholm, J.D. The intestinal barrier and its regulation by neuroimmune factors. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2010, 22, 718–733. [Google Scholar] [CrossRef]
- Scudamore, C.L.; Jepson, M.A.; Hirst, B.H.; Miller, H.R. The rat mucosal mast cell chymase, RMCP-II, alters epithelial cell monolayer permeability in association with altered distribution of the tight junction proteins ZO-1 and occludin. Eur. J. Cell Biol. 1998, 75, 321–330. [Google Scholar] [CrossRef]
- Patrick, M.K.; Dunn, I.J.; Buret, A.; Miller, H.R.; Huntley, J.F.; Gibson, S.; Gall, D.G. Mast cell protease release and mucosal ultrastructure during intestinal anaphylaxis in the rat. Gastroenterology 1988, 94, 1–9. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.N.; Qu, J.; Liu, M.; Tso, P. The Role of Interstitial Matrix and the Lymphatic System in Gastrointestinal Lipid and Lipoprotein Metabolism. Front. Physiol. 2020, 11, 4–13. [Google Scholar] [CrossRef]
- Ghoshal, S.; Witta, J.; Zhong, J.; de Villiers, W.; Eckhardt, E. Chylomicrons promote intestinal absorption of lipopolysaccharides. J. Lipid Res. 2009, 50, 90–97. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Fan, X.X.; Liu, N.; Song, Q.Q.; Kou, J.; Shi, Y.H.; Luo, X.; Dai, Z.L.; Yang, Y.; Wu, Z.L.; et al. L-Glutamine Represses the Unfolded Protein Response in the Small Intestine of Weanling Piglets. J. Nutr. 2019, 149, 1904–1910. [Google Scholar] [CrossRef] [PubMed]
- Ockenga, J.; Borchert, K.; Stuber, E.; Lochs, H.; Manns, M.P.; Bischoff, S.C. Glutamine-enriched total parenteral nutrition in patients with inflammatory bowel disease. Eur. J. Clin. Nutr. 2005, 59, 1302–1309. [Google Scholar] [CrossRef] [Green Version]
- Akobeng, A.K.; Miller, V.; Stanton, J.; Elbadri, A.M.; Thomas, A.G. Double-blind randomized controlled trial of glutamine-enriched polymeric diet in the treatment of active Crohn’s disease. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 78–84. [Google Scholar] [CrossRef] [PubMed]
- MKaK, M. Modulation of mast cell function by amino acids in vitro: A potential mechanism of immunonutrition for wound healing. J. Nutr. Health Food Sci. 2013, 1, 6–12. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Qu, J.; Yang, Q.; Wu, Z.; Liu, M.; Tso, P. Effect of L-Glutamine on Chylomicron Formation and Fat-Induced Activation of Intestinal Mucosal Mast Cells in Sprague-Dawley Rats. Nutrients 2022, 14, 1777. https://doi.org/10.3390/nu14091777
He Y, Qu J, Yang Q, Wu Z, Liu M, Tso P. Effect of L-Glutamine on Chylomicron Formation and Fat-Induced Activation of Intestinal Mucosal Mast Cells in Sprague-Dawley Rats. Nutrients. 2022; 14(9):1777. https://doi.org/10.3390/nu14091777
Chicago/Turabian StyleHe, Yu, Jie Qu, Qing Yang, Zhenlong Wu, Min Liu, and Patrick Tso. 2022. "Effect of L-Glutamine on Chylomicron Formation and Fat-Induced Activation of Intestinal Mucosal Mast Cells in Sprague-Dawley Rats" Nutrients 14, no. 9: 1777. https://doi.org/10.3390/nu14091777