
Fiber Consumption Mediates Differences in Several Gut Microbes in a Subpopulation of Young Mexican Adults


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Subjects
2.2. Variables and Data Collection
2.2.1. Dietary Variables
2.2.2. Anthropometric Variables
2.3. Microbial Determination
2.3.1. Stool Collection
2.3.2. DNA Extraction
2.3.3. Identification of the Intestinal Microbiota
2.4. Statistical Analysis
3. Results
Participants
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iversen, K.N.; Carlsson, F.; Andersson, A.; Michaelsson, K.; Langton, M.; Riserus, U.; Hellstrom, P.M.; Landberg, R. A hypocaloric diet rich in high fiber rye foods causes greater reduction in body weight and body fat than a diet rich in refined wheat: A parallel randomized controlled trial in adults with overweight and obesity (the RyeWeight study). Clin. Nutr. ESPEN 2021, 45, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kelly, A.S. Review of childhood obesity: From epidemiology, etiology, and comorbidities to clinical assessment and treatment. Mayo Clin. Proc. 2017, 92, 251–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varkevisser, R.D.M.; van Stralen, M.M.; Kroeze, W.; Ket, J.C.F.; Steenhuis, I.H.M. Determinants of weight loss maintenance: A systematic review. Obes. Rev. 2019, 20, 171–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, L.M.Y.; Fong, S.S.M.; Law, Q.P.S. Younger Adults Are More Likely to Increase Fruit and Vegetable Consumption and Decrease Sugar Intake with the Application of Dietary Monitoring. Nutrients 2021, 13, 333. [Google Scholar] [CrossRef] [PubMed]
- Sprake, E.; Russell, J.; Cecil, J.; Cooper, R.; Grabowski, P.; Pourshahidi, L.K.; Barker, M. Dietary patterns of university students in the UK: A cross-sectional study. Nutr. J. 2018, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ramón-Arbués, E.; Granada-López, J.-M.; Martínez-Abadía, B.; Echániz-Serrano, E.; Antón-Solanas, I.; Jerue, B.A. Factors Related to Diet Quality: A Cross-Sectional Study of 1055 University Students. Nutrients 2021, 13, 3512. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, A.; Ali, B.; Cameron, A.; Armstrong, M.E.; Isaacs, P.; Thomas, K.S.; Gadbois, E.A.; Willis, P. ‘It’s not just about the dinner; it’s about everything else that we do’: A qualitative study exploring how Meals on Wheels meet the needs of self-isolating adults during COVID-19. Health Soc. Care Community 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Mitri, R.; Boulos, C.; Ziade, F. Mediterranean diet adherence among adolescents in North Lebanon: The role of skipping meals, meals with the family, physical activity, and physical wellbeing. Br. J. Nutr. 2021, 1–8, in press. [Google Scholar] [CrossRef] [PubMed]
- Nuss, T.; Morley, B.; Scully, M.; Wakefield, M. Energy drink consumption among Australian adolescents associated with a cluster of unhealthy dietary behaviours and short sleep duration. Nutr. J. 2021, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Granata, I.; Nardelli, C.; D’Argenio, V.; Tramontano, S.; Compare, D.; Guarracino, M.R.; Nardone, G.; Pilone, V.; Sacchetti, L. Duodenal Metatranscriptomics to Define Human and Microbial Functional Alterations Associated with Severe Obesity: A Pilot Study. Microorganisms 2020, 8, 1811. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Zhang, J. Role of intestinal microbiota and metabolites on gut homeostasis and human diseases. BMC Immunol. 2017, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevas-Sierra, A.; Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, and Obesity: Links with Host Genetics and Epigenetics and Potential Applications. Adv. Nutr. 2019, 10, S17–S30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornejo-Pareja, I.; Munoz-Garach, A.; Clemente-Postigo, M.; Tinahones, F.J. Importance of gut microbiota in obesity. Eur. J. Clin. Nutr. 2019, 72, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myhrstad, M.C.W.; Tunsjo, H.; Charnock, C.; Telle-Hansen, V.H. Dietary Fiber, Gut Microbiota, and Metabolic Regulation-Current Status in Human Randomized Trials. Nutrients 2020, 12, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Rajoka, M.S.R.; Shi, J.; Mehwish, H.M.; Zhu, J.; Li, Q.; Shao, D.; Huang, Q.; Yang, H. Interaction between diet composition and gut microbiota and its impact on gastrointestinal tract health. Food Sci. Hum. Wellness 2017, 6, 121–130. [Google Scholar] [CrossRef]
- Martinez, T.M.; Meyer, R.K.; Duca, F.A. Therapeutic Potential of Various Plant-Based Fibers to Improve Energy Homeostasis via the Gut Microbiota. Nutrients 2021, 13, 3470. [Google Scholar] [CrossRef] [PubMed]
- Poobalan, A.S.; Aucott, L.S.; Clarke, A.; Smith, W.C. Diet behaviour among young people in transition to adulthood (18–25 year olds): A mixed method study. Health Psychol. Behav. Med. 2014, 2, 909–928. [Google Scholar] [CrossRef]
- Hypponen, E.; Carslake, D.; Berry, D.J.; Power, C.; Davey Smith, G. Estimating the influence of body mass index (BMI) on mortality using offspring BMI as an instrumental variable. Int. J. Obes. 2022, 46, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Macedo-Ojeda, G.; Vizmanos-Lamotte, B.; Marquez-Sandoval, Y.F.; Rodriguez-Rocha, N.P.; Lopez-Uriarte, P.J.; Fernandez-Ballart, J.D. Validation of a semi-quantitative food frequency questionnaire to assess food groups and nutrient intake. Nutr. Hosp. 2013, 28, 2212–2220. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Díaz, J.; Molina-Montes, E.; Soto-Méndez, M.J.; Madrigal, C.; Hernández-Ruiz, Á.; Valero, T.; Lara Villoslada, F.; Leis, R.; Martínez de Victoria, E.; Moreno, J.M. Clustering of dietary patterns and lifestyles among spanish children in the EsNuPi study. Nutrients 2020, 12, 2536. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, C.; Soto-Méndez, M.J.; Hernández-Ruiz, Á.; Valero, T.; Ávila, J.M.; Ruiz, E.; Lara Villoslada, F.; Leis, R.; Martínez de Victoria, E.; Moreno, J.M. Energy intake, macronutrient profile and food sources of spanish children aged one to <10 years—Results from the EsNuPI Study. Nutrients 2020, 12, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tester, J.M.; Leak, T.M. Fiber-rich foods delivered to Low-Income Households: A feasibility study of children with prediabetes and spillover effect on their caregivers. Prev. Med. Rep. 2021, 24, 101511. [Google Scholar] [CrossRef] [PubMed]
- United States. Dietary Guidelines Advisory Committee. Dietary Guidelines for Americans, 2010; US Department of Health and Human Services, US Department of Agriculture: Washington, DC, USA, 2010.
- Pérez-Lizaur, A.; Palacios-González, B.; Castro-Becerra, A.; Flores-Galicia, I. Sistema Mexicano de Alimentos Equivalentes (Mexican Equivalent Food System); Fomento de Nutrición y Salud, AC: Mexico City, Mexico, 2014; pp. 31–36. [Google Scholar]
- Procuraduría Federal del Consumidor. Gobierno de Mexico, PROFECO. Available online: https://www.gob.mx/profeco (accessed on 12 February 2022).
- Plaza-Diaz, J.; Fernandez-Caballero, J.A.; Chueca, N.; Garcia, F.; Gomez-Llorente, C.; Saez-Lara, M.J.; Fontana, L.; Gil, A. Pyrosequencing analysis reveals changes in intestinal microbiota of healthy adults who received a daily dose of immunomodulatory probiotic strains. Nutrients 2015, 7, 3999–4015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinttila, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, T.; Watanabe, K.; Fujimoto, J.; Takada, T.; Tanaka, R. Use of 16S rRNA gene-targeted group-specific primers for real-time PCR analysis of predominant bacteria in human feces. Appl. Environ. Microbiol. 2004, 70, 7220–7228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.; Tannock, G.W.; Tilsala-Timisjarvi, A.; Rodtong, S.; Loach, D.M.; Munro, K.; Alatossava, T. Detection and identification of gastrointestinal Lactobacillus species by using denaturing gradient gel electrophoresis and species-specific PCR primers. Appl. Environ. Microbiol. 2000, 66, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- McGuire, S. US department of agriculture and US department of health and human services, dietary guidelines for Americans, 2010. Washington, DC: US government printing office, January 2011. Adv. Nutr. 2011, 2, 293–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astrup, A. Healthy lifestyles in Europe: Prevention of obesity and type II diabetes by diet and physical activity. Public Health Nutr. 2001, 4, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Asif, M. The prevention and control the type-2 diabetes by changing lifestyle and dietary pattern. J. Educ. Health Promot. 2014, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallus, S.J.; Brandt, L.J. The intestinal microbiota and obesity. J. Clin. Gastroenterol. 2012, 46, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Dore, J.; Blottiere, H. The influence of diet on the gut microbiota and its consequences for health. Curr. Opin. Biotechnol. 2015, 32, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Yokomichi, H.; Matsubara, H.; Ishikuro, M.; Kikuya, M.; Isojima, T.; Yokoya, S.; Tanaka, T.; Kato, N.; Chida, S.; et al. Longitudinal changes in body mass index of children affected by the Great East Japan Earthquake. Int. J. Obes. 2017, 41, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.; Threlkeld, R.J. Nutritional Recommendations for Individuals with Diabetes. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2019. [Google Scholar] [PubMed]
- Koo, S.H.; Chu, C.W.; Khoo, J.J.C.; Cheong, M.; Soon, G.H.; Ho, E.X.P.; Law, N.M.; De Sessions, P.F.; Fock, K.M.; Ang, T.L.; et al. A pilot study to examine the association between human gut microbiota and the host’s central obesity. JGH Open 2019, 3, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Sogari, G.; Velez-Argumedo, C.; Gomez, M.I.; Mora, C. College Students and Eating Habits: A Study Using An Ecological Model for Healthy Behavior. Nutrients 2018, 10, 1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.L.; Heaver, S.L.; Walters, W.A.; Ley, R.E. Microbiome and metabolic disease: Revisiting the bacterial phylum Bacteroidetes. J. Mol. Med. 2017, 95, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhang, B.; Hu, J.; Nie, S.; Xiong, T.; Xie, M. Intervention of five strains of Lactobacillus on obesity in mice induced by high-fat diet. J. Funct. Foods 2020, 72, 104078. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Backhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frugé, A.D.; Van der Pol, W.; Rogers, L.Q.; Morrow, C.D.; Tsuruta, Y.; Demark-Wahnefried, W. Fecal Akkermansia muciniphila is associated with body composition and microbiota diversity in overweight and obese women with breast cancer participating in a presurgical weight loss trial. J. Acad. Nutr. Diet. 2020, 120, 650–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Q.; Li, D.; He, Y.; Li, Y.; Yang, Z.; Zhao, X.; Liu, Y.; Wang, Y.; Sun, J.; Feng, X.; et al. Discrepant gut microbiota markers for the classification of obesity-related metabolic abnormalities. Sci. Rep. 2019, 9, 13424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, A.; Sasaki, T.; Itoh, K.; Kitahara, T.; Takema, Y.; Hiramatsu, K.; Ishikawa, D.; Shibuya, T.; Kobayashi, O.; Osada, T.; et al. A Soluble Fiber Diet Increases Bacteroides fragilis Group Abundance and Immunoglobulin A Production in the Gut. Appl. Environ. Microbiol. 2020, 86, e00405-20. [Google Scholar] [CrossRef] [PubMed]
- Menni, C.; Jackson, M.A.; Pallister, T.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int. J. Obes. 2017, 41, 1099–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, C.T.; Yeh, Y.T.; Lin, C.C.; Yang, L.Y.; Chiang, C.P. Akkermansia muciniphila is Negatively Correlated with Hemoglobin A1c in Refractory Diabetes. Microorganisms 2020, 8, 1360. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; de Vos, W.M. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, S.L.; Hervey, T.K.; Mackenbach, J.P.; McKee, M. Health law and policy in the European Union. Lancet 2013, 381, 1135–1144. [Google Scholar] [CrossRef]
- Burris, S.; Anderson, E. Legal regulation of health-related behavior: A half century of public health law research. Annu. Rev. Law Soc. Sci. 2013, 9, 95–117. [Google Scholar] [CrossRef]




{kind=link}
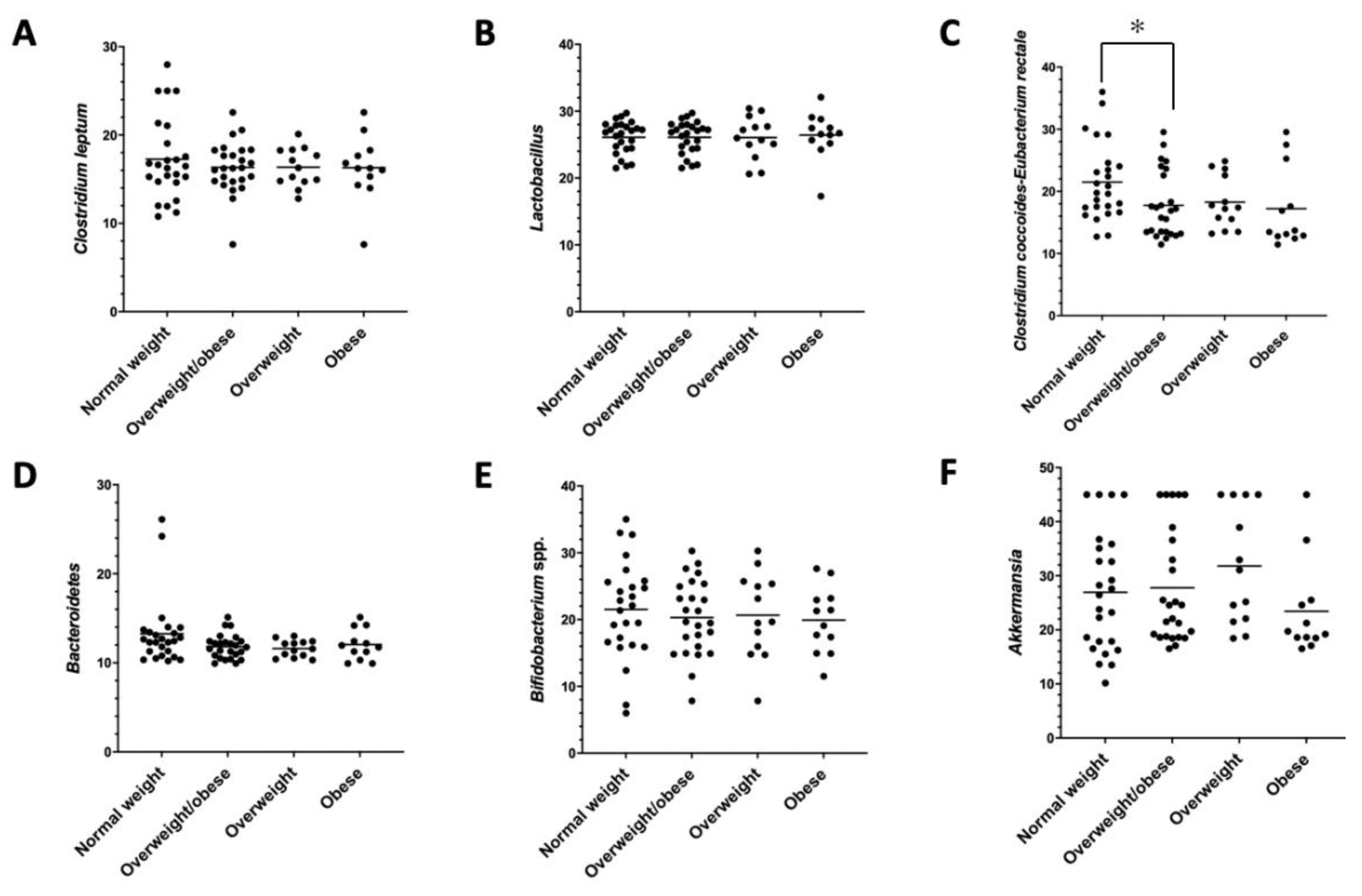{kind=link}
{kind=link}
| PCR Assay | Sequence | Target Species | Size (bp) | Cycles and Tm (°C) | Reference |
|---|---|---|---|---|---|
| Bacteroidetes (Bacteroides, Prevotella and Porphyromonas) | F. 5′-GGTGTCGGCTTAAGTGCCAT-3′, R. 5′-CGGA(C/T)GTAAGGGCCGTGC-3′ | Bacteroides fragilis, B. stercoris, B. vulgatus, B. eggerthii, B. acidofaciens, B. caccae, B. ovatus, B. uniformis, B. thetaiotaomicron, B. distasonis, B. merdae, B. forsythus, Prevotella tannerae, P. bryantii, P. ruminicola, P. heparinolytica, P. zoogleoformans, P. brevis, P. loescheii, P. buccae, P. oralis, P. enoeca, P. melaninogenica, P. veroralis, P. intermedia P. albensis, P. nigrescens, P. corporis, P. disiens, P. bivia, P. pallens, P. denticola, Porphyromonas canoris, P. gingivalis, P. asaccharolytica, P. levii, P. cangingivalis, P. endodontalis, P. macacae, P. circumdentaria, P. catoniae | 140 | Polymerase activation at 95 °C for 10 min, and 45 cycles of denaturation (95 °C/10 s), then annealing (68 °C/8 s), and extension (72 °C/6 s); 68 °C | Rinttilä, (2004) [30] |
| Actinobacteria (Bifidobacterium spp.) | F. 5′-TCGCGTC(C/T)GGTGTGAAAG-3′, R. 5′-CCACATCCAGC(A/G)TCCAC-3′ | Bifidobacterium longum, B. minimum, B. angulatum, B. catenulatum, B. pseudocatenulatum, B. dentium, B. ruminantium, B. thermophilum, B. subtile, B. bifidum, B. boum, B. lactis, B. animalis, B. choerinum, B. gallicum, B. pseudolongum subsp. globosum, B. pseudolongum subsp. pseudolongum, B. magnum, B. infantis, B. indicum, B. gallinarum, B. pullorum, B. saeculare, B. suis | 243 | Polymerase activation at 95 °C for 10 min, and 45 cycles of denaturation (95 °C/10 s), then annealing (58 °C/8 s), and extension (72 °C/10 s); 58 °C | Rinttilä, (2004) [30] |
| Firmicutes (Clostridium coccoides-Eubacterium rectale) | F. 5′-CGGTACCTGACTAAGAAGC-3′, R. 5′-AGTTT(C/T)ATTCTTGCGAACG-3′ | Clostridium coccoides, C. proteoclasticum, C. aminophilum, C. symbiosum, C. sphenoides, C. celerecrescens, C. aerotolerans, C. xylanolyticum, C. clostridiiforme, C. fusiformis, C. nexile, C. oroticum, C. populeti, C. aminovalericum, C. indolis, C. herbivorans, C. polysaccharolyticum, Eubacterium xylanophilum, E. ruminantium, E. saburreum, E. fissicatena, E. hadrum, E. rectale, E. ramulus, E. contortum, E. eligens, E. hallii, E. formicigenerans, E. cellulosolvens, Ruminococcus productus, R. obeum, R. schinkii, R. hydrogenotrophicus, R. hansenii, R. torques, R. lactaris, R. gnavus, Butyrivibrio fibrisolvens, B. crossotus, B. fibrisolvens, Desulfotomaculum guttoideum, Roseburia cecicola, Pseudobutyrivibrio ruminis, Lachnospira multipara, L. pectinoschiza, Acetitomaculum ruminis, Catonella morbi | 429 | Polymerase activation at 95 °C for 10 min, and 45 cycles of denaturation (95 °C/10 s), then annealing (58 °C/8 s), and extension (72 °C/10 s); 58 °C | Rinttilä, (2004) [30] |
| Firmicutes (Clostridium leptum) | F. 5′-GCA CAA GCA GTG GAGT-3′, R. 5′-CTT CCT CCG TTT TGT CAA-3′ | Clostridium leptum, C. viride, Eubacterium siraeum, Ruminococcus bromii, R. callidus, R. albus | 239 | Polymerase activation at 95 °C for 10 min, and 45 cycles of denaturation (95 °C/10 s), then annealing (58 °C/8 s), and extension (72 °C/14 s); 50 °C | Matsuki, (2004) [31] |
| Akkermansia | F.5′ CAGCACGTGAAGGTGGGAC-3′, R. 5′-CCTTGCGGTTG GCTTCAGAT-3′ | Akkermansia muciniphila | Polymerase activation at 95 °C for 10 min, and 45 cycles of denaturation (95 °C/10 s), then annealing (58 °C/8 s), and extension (72 °C/30 s); 62 °C | Dao, (2016) [32] | |
| Lactobacillus | F.5′-AGCAGTAGGGAATCTTCCA-3′, R. 5′-CACCGCTACACATGGAG-3′ | Lactobacillus acidophilus, L. amylovorus, L. delbrueckii subsp. bulgaricus, L. delbrueckii subsp. delbrueckii, L. delbrueckii subsp. lactis, L. amylolyticus, L. acetotolerans, L. crispatus, L. amylophilus, L. johnsonii, L. gasseri, L. fermentum, L. pontis, L. reuteri, L. mucosae, L. vaginalis, L. panis, L. oris, L. pentosus, L. plantarum, L. collinoides, L. alimentarius, L. farciminis, L. brevis, L. buchneri, L. kefiri, L. fructivorans, L. mali, L. animalis, L. murinus, L. ruminis, L. agilis, L. salivarius subsp. salicinius, L. aviarius subsp. aviarius, L. sharpeae, L. manihotivorans, L. rhamnosus, L. casei subsp. casei, L. zeae, L. paracasei subsp. paracasei, L. paracasei subsp. tolerans, L. coryniformis subsp. coryniformis, L. bifermentans, L. perolens, L. sakei subsp. sakei, L. casei subsp. fusiformis, Pediococcus pentosaceus, P. parvulus, P. acidilactici, P. dextrinicus, Weissella halotolerans, W. confusus, W. Paramesenteroides, W. hellenica, W. viridescens, W. kandleri, W. minor, Leuconostoc lactis | Amplification program was 92 °C for 2 min, followed by 40 cycles of 95 °C for 30 s, 30 s at the appropriate annealing temperature, and 72 °C for 30 s; 56 °C | Walter et al., (2001) [33] |
| Characteristics | Normal-Weight n = 25 | Overweight/Obese n = 25 | Overweight n = 13 | Obese n = 12 | p-Value |
|---|---|---|---|---|---|
| Sociodemographic Data | |||||
| Age, years | 20.5 ± 1.7 | 20.7 ± 1.7 | 21 ± 1.6 | 21 ± 1.8 | NS |
| Sex, F/M | 15/10 | 12/13 | 6/7 | 6/6 | NS |
| Anthropometric Data | |||||
| Weight (Kg) | 59.0 ± 8.0 | 85.0 ± 12.0 | 80.0 ± 9.0 | 89.0 ± 14.0 | 0.0001 |
| Height (cm) | 165.0 ± 9.0 | 169.0 ± 9.0 | 170.0 ± 9.0 | 167.0 ± 10.0 | NS |
| BMI, kg/m2 | 21.9 ± 1.9 | 29.6 ± 3.7 | 27.7 ± 2.1 | 31.8 ± 4.0 | 0.0001 |
| Body fat, % | 23.3 ± 6.7 | 32.5 ± 7.1 | 31.5 ± 7.3 | 33.6 ± 7.0 | 0.0001 |
| Body water, % | 56.9 ± 4.8 | 51.7 ± 4.8 | 51.3 ± 4.5 | 52.1 ± 5.3 | 0.003 |
| Visceral fat, % | 1.6 ± 0.8 | 5.4 ± 2.7 | 4.8 ± 1.9 | 6.0 ± 3.4 | 0.0001 |
| Muscle | 43.5 ± 8.3 | 53.8 ± 11.1 | 52.3 ± 11.5 | 55.5 ± 10.9 | 0.0007 |
| Basal metabolic rate | 1432.0 ± 226.0 | 1704.0 ± 330.0 | 1676.0 ± 297.0 | 1735.0 ± 374.0 | 0.002 |
| Metabolic age | 17.4 ± 6.3 | 44.8 ± 13.7 | 42.9 ± 10.2 | 46.7 ± 16.9 | 0.0001 |
| Bone mass | 2.36 ± 0.4 | 3.1 ± 1.1 | 3.2 ± 1.5 | 3.1 ± 0.5 | 0.0001 |
| Portions/Day | Normal-Weight n = 25 | Overweight/Obese n = 25 | p-Value |
|---|---|---|---|
| Dairy products | 3.8 (0.4–13.8) | 3.8 (1.0–10.9) | 0.915 |
| Fruits | 7.5 (3.0–14.8) | 5.2 (1.5–10.5) | 0.043 |
| Vegetables | 9.5 (3.5–15.3) | 6.3 (2.8–13.0) | 0.066 |
| Cereal with fat | 4.9 (2.6–8.7) | 3.9 (0.8–9.8) | 0.132 |
| Cereal without fat | 6.0 (3.1–11.2) | 5.0 (2.3–6.4) | 0.005 |
| Animal protein foods | 2.2 (1.4–4.9) | 2.1 (0.5–4.9) | 0.455 |
| Vegetable protein foods | 0.4 (0.1–1.4) | 0.7 (0.1–1.9) | 0.331 |
| Oils and fats with protein | 1.2 (0.3–5.6) | 1.0 (0.0–7.7) | 0.414 |
| Oils and fats without protein | 4.4 (1.6–8.9) | 2.9 (1.0–5.8) | 0.022 |
| Sugars | 6.4 (2.9–10.8) | 6.5 (2.1–8.7) | 0.472 |
| Alcoholic beverages | 1.4 (0.0–2.5) | 1.0 (0.0–2.5) | 0.682 |
| Fiber (g per day) | 19.9 (9.5–37.2) | 19.0 (7.4–35.8) | 0.366 |
| Fermented dairy foods (CFU × 109 per day) | 18.3 (0.0–240.4) | 29.6 (0–177.4) | 0.661 |
| Variables | Normal Weight n = 25 | Overweight/Obesity n = 25 | p-Value |
|---|---|---|---|
| Total energy | 3122 (1600–6843) | 2825 (1002–4953) | 0.232 |
| kcal/day | |||
| Carbohydrates | 464 (240–1005) | 394 (151–755) | 0.063 |
| g/day a | |||
| Kcal/day | 1886 (969–4031) | 1576 (604–3044) | 0.066 |
| % daily b | 60.4 (86–337) | 55.7(44–210) | 0.005 |
| Proteins | 95 (53–273) | 101 (33–190) | 0.763 |
| g/day a | |||
| Kcal/day | 382 (215–1093) | 404 (133–761) | 0.763 |
| % daily b | 12.2 (76–349) | 14.3 (41–221) | 0.037 |
| Lipids | 86 (44–204) | 80 (28–138) | 0.377 |
| g/day a | |||
| Kcal/day | 774 (398–1841) | 724 (252–1216) | 0.377 |
| % daily b | 24.7 (76–311) | 25.6 (40–191) | 0.017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Lara, A.; Plaza-Díaz, J.; López-Uriarte, P.; Vázquez-Aguilar, A.; Reyes-Castillo, Z.; Álvarez-Mercado, A.I. Fiber Consumption Mediates Differences in Several Gut Microbes in a Subpopulation of Young Mexican Adults. Nutrients 2022, 14, 1214. https://doi.org/10.3390/nu14061214
Rodríguez-Lara A, Plaza-Díaz J, López-Uriarte P, Vázquez-Aguilar A, Reyes-Castillo Z, Álvarez-Mercado AI. Fiber Consumption Mediates Differences in Several Gut Microbes in a Subpopulation of Young Mexican Adults. Nutrients. 2022; 14(6):1214. https://doi.org/10.3390/nu14061214
Chicago/Turabian StyleRodríguez-Lara, Avilene, Julio Plaza-Díaz, Patricia López-Uriarte, Alejandra Vázquez-Aguilar, Zyanya Reyes-Castillo, and Ana I. Álvarez-Mercado. 2022. "Fiber Consumption Mediates Differences in Several Gut Microbes in a Subpopulation of Young Mexican Adults" Nutrients 14, no. 6: 1214. https://doi.org/10.3390/nu14061214