
Exploring the Chemical Space of Kawakawa Leaf (Piper excelsum)
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Solvents and Chemicals
2.2. Preparation of Standards
2.3. Kawakawa Leaf Collection
2.4. Extraction and Sample Preparation for LC-MS/MS
2.4.1. Aqueous Extraction
2.4.2. Solvent Extraction
2.5. LC-MS/MS Instrument and Conditions
2.6. Data Processing, Visualisation and Statistical Analysis
3. Results
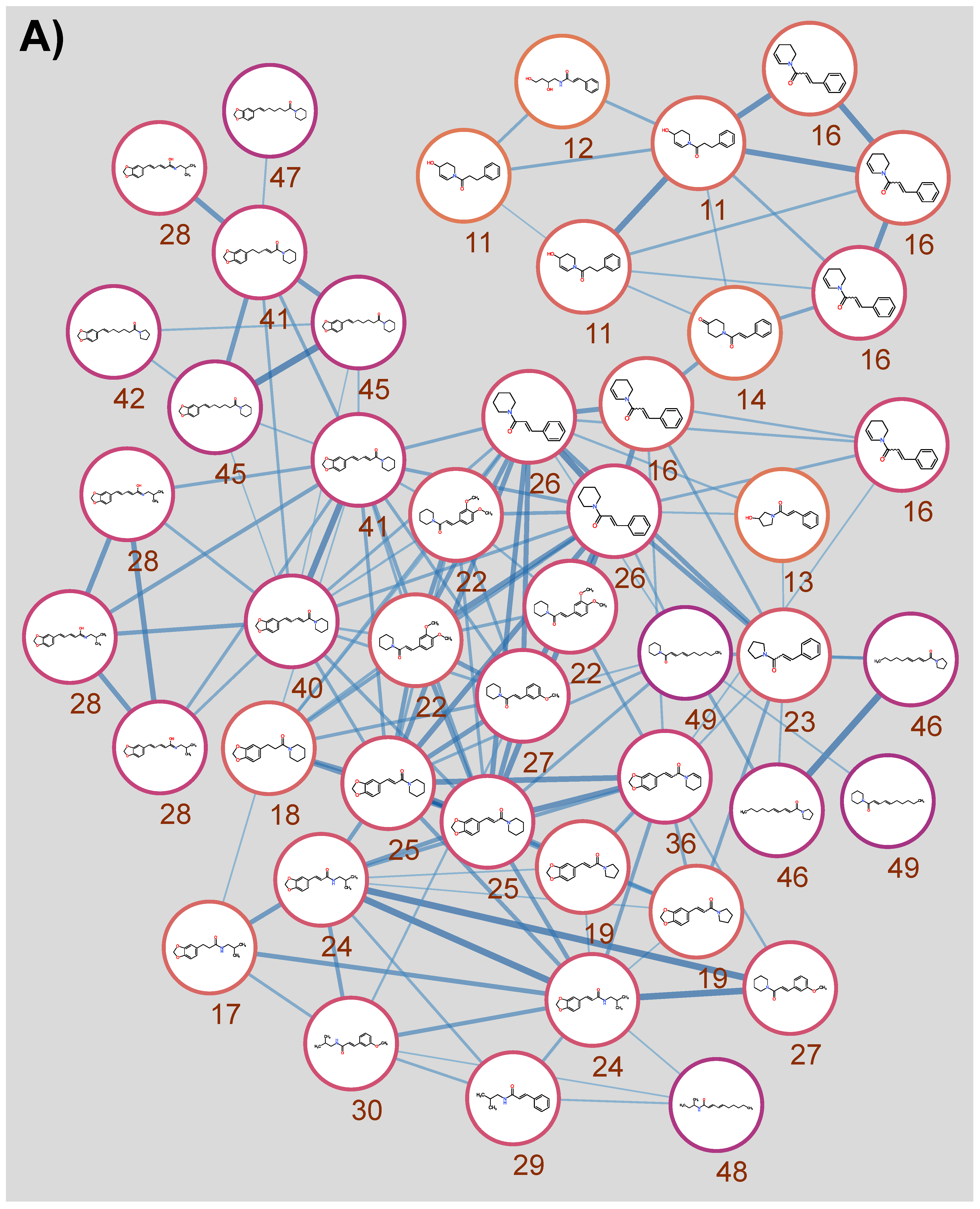
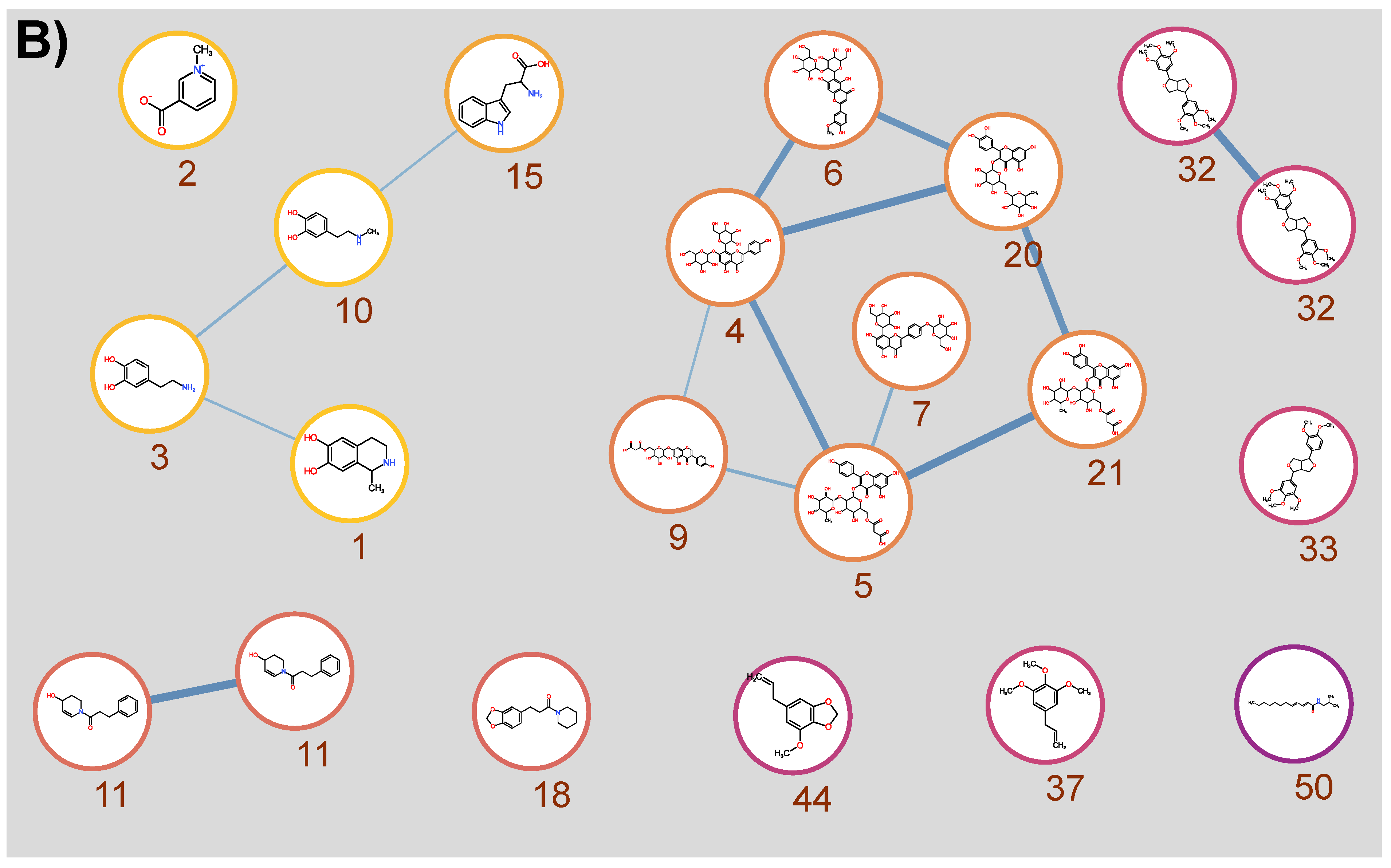
3.1. Qualitative Metabolic Profiling of Kawakawa Aqueous Extracts
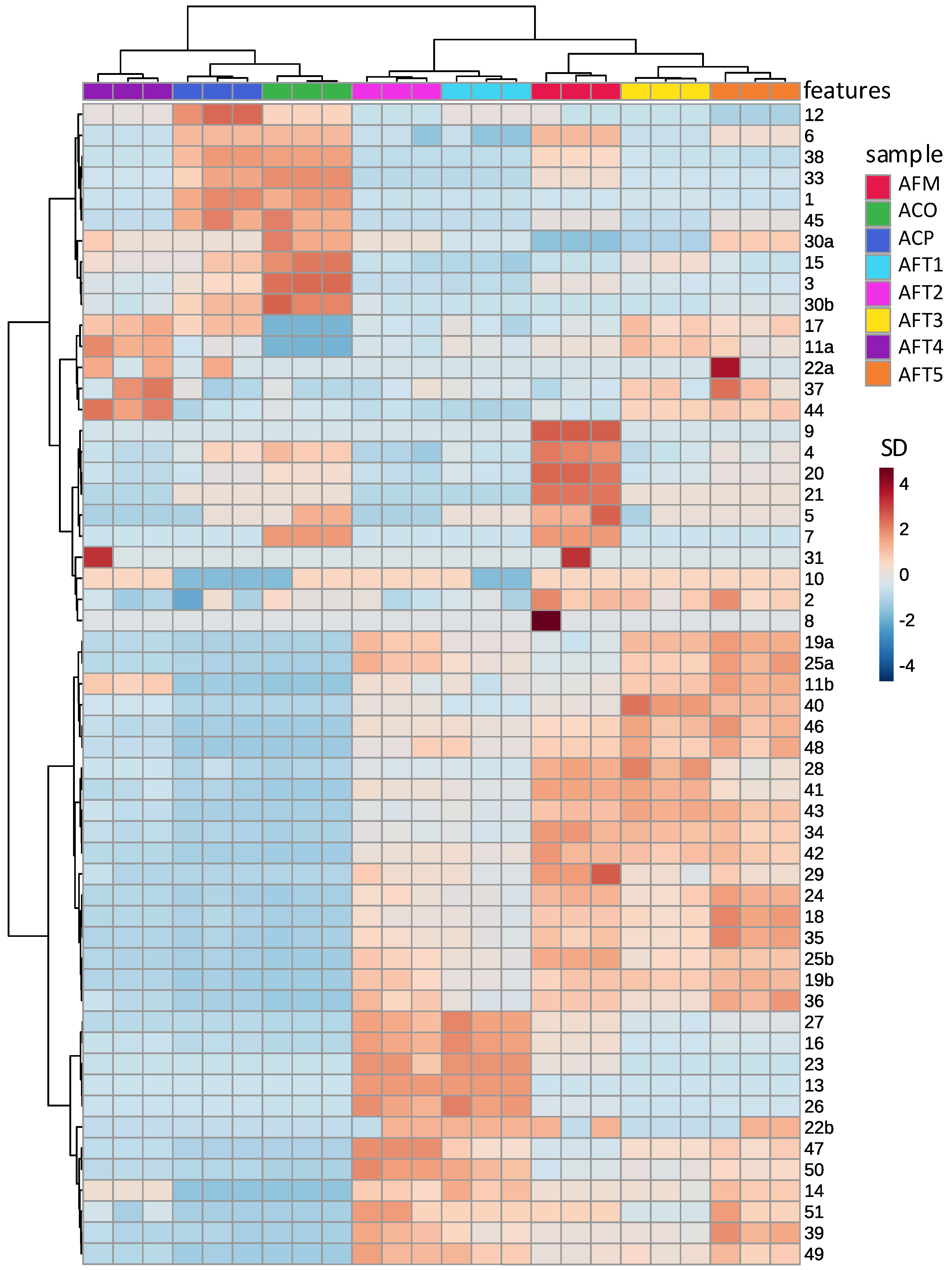
3.2. Differences in Kawakawa Chemistry
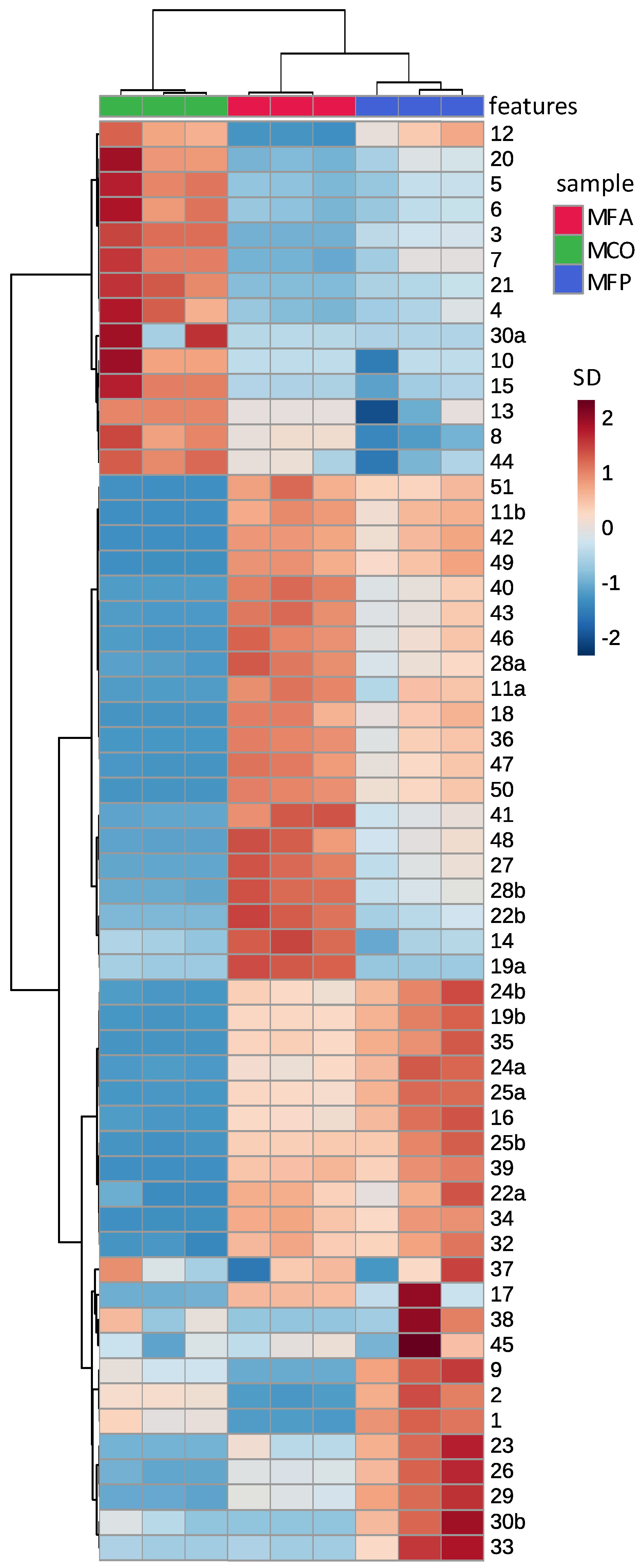
3.3. The Comparison of Aqueous and Methanolic Kawakawa Extracts
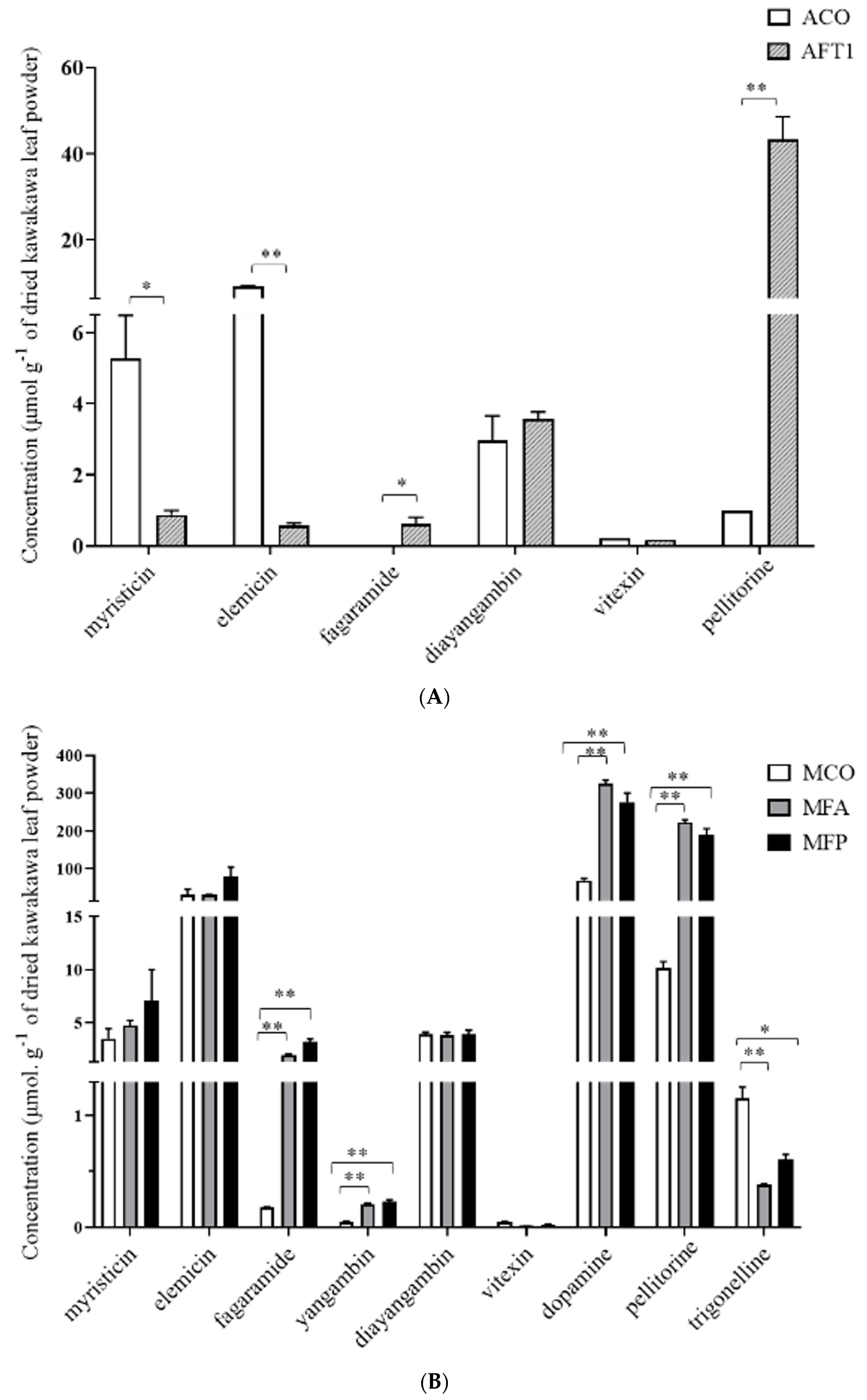
3.4. Absolute Quantitative Analysis of Aqueous and Methanolic Kawakawa Extracts
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brooker, S.G.; Cooper, R.C. New Zealand Medicinal Plants. Econ. Bot. 1961, 15, 1–10. [Google Scholar] [CrossRef]
- Xu, H. Anti-Inflammatory and Anti-Cancer Effects of Kawakawa (Macropiper excelsum) Leaves. Master’s Thesis, The University of Auckland, Auckland, New Zealand, 2018. [Google Scholar]
- Ramzan, F.; Jayaprakash, R.; Pook, C.; Foster, M.; Miles-Chan, J.L.; Mithen, R. Acute Effects of Kawakawa (Piper excelsum) Intake on Postprandial Glycemic and Insulinaemic Response in a Healthy Population. Nutrients 2022, 14, 1638. [Google Scholar] [CrossRef] [PubMed]
- Obst, K.; Lieder, B.; Reichelt, K.V.; Backes, M.; Paetz, S.; Geißler, K.; Krammer, G.; Somoza, V.; Ley, J.P.; Engel, K.H. Sensory Active Piperine Analogues from Macropiper excelsum and Their Effects on Intestinal Nutrient Uptake in Caco-2 Cells. Phytochemistry 2017, 135, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Butts, C.A.; van Klink, J.W.; Joyce, N.I.; Paturi, G.; Hedderley, D.I.; Martell, S.; Harvey, D. Composition and Safety Evaluation of Tea from New Zealand Kawakawa (Piper excelsum). J. Ethnopharmacol. 2019, 232, 110–118. [Google Scholar] [CrossRef]
- Takooree, H.; Aumeeruddy, M.Z.; Rengasamy, K.R.R.; Venugopala, K.N.; Jeewon, R.; Zengin, G.; Mahomoodally, M.F. A Systematic Review on Black Pepper (Piper nigrum L.): From Folk Uses to Pharmacological Applications. Crit. Rev. Food Sci. Nutr. 2019, 59, S210–S243. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Jaiswal, A.; Subhashini; Singh, R. Combination Therapy with Curcumin Alone Plus Piperine Ameliorates Ovalbumin-Induced Chronic Asthma in Mice. Inflammation 2018, 41, 1922–1933. [Google Scholar] [CrossRef]
- Okumura, Y.; Narukawa, M.; Iwasaki, Y.; Ishikawa, A.; Matsuda, H.; Yoshikawa, M.; Watanabe, T. Activation of TRPV1 and TRPA1 by Black Pepper Components. Biosci. Biotechnol. Biochem. 2010, 74, 1068–1072. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Nam, M.; Kang, M.S.; Lee, J.O.; Lee, Y.W.; Hwang, G.S.; Kim, H.S. Piperine Regulates UCP1 through the AMPK Pathway by Generating Intracellular Lactate Production in Muscle Cells. Sci. Rep. 2017, 7, 41066. [Google Scholar] [CrossRef]
- Shulgin, A.T. Possible Implication of Myristicin as a Psychotropic Substance. Nature 1966, 210, 380–384. [Google Scholar] [CrossRef]
- Beyer, J.; Ehlers, D.; Maurer, H.H. Abuse of Nutmeg (Myristica fragrans Houtt.): Studies on the Metabolism and the Toxicologic Detection of Its Ingredients Elemicin, Myristicin, and Safrole in Rat and Human Urine Using Gas Chromatography/Mass Spectrometry. Ther. Drug Monit. 2006, 28, 568–575. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vandergheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen Rearrangement Rules: Computational MS/MS Fragmentation and Structure Elucidation Using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying Metabolites by Integrating Metabolome Databases with Mass Spectrometry Cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Otasek, D.; Morris, J.H.; Bouças, J.; Pico, A.R.; Demchak, B. Cytoscape Automation: Empowering Workflow-Based Network Analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive Linear Step-up Procedures That Control the False Discovery Rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Burgess, E.J.; Richardson, A.T.B.; Hawkins, B.C.; Baird, S.K.; Smallfield, B.M.; Van Klink, J.W.; Perry, N.B. Cytotoxic Amides from Fruits of Kawakawa, Macropiper excelsum. Planta Med. 2015, 81, 1163–1168. [Google Scholar] [CrossRef] [Green Version]
- Russell, G.B.; Fenemore, P.G. New Lignans from Leaves of Macropiper excelsum. Phytochemistry 1973, 12, 1799–1803. [Google Scholar] [CrossRef]
- Briggs, L.H. The Essential Oil of Macropiper excelsum (Kawakawa). J. Soc. Chem. Ind. 1941, 60, 210–212. [Google Scholar] [CrossRef]
- Martha Perez Gutierrez, R.; Maria Neira Gonzalez, A.; Hoyo-Vadillo, C. Alkaloids from Piper: A Review of Its Phytochemistry and Pharmacology. Mini Rev. Med. Chem. 2013, 13, 163–193. [Google Scholar]
- Dragull, K.; Yoshida, W.Y.; Tang, C.S. Piperidine Alkaloids from Piper methysticum. Phytochemistry 2003, 63, 193–198. [Google Scholar] [CrossRef]
- Schnabel, A.; Athmer, B.; Manke, K.; Schumacher, F.; Cotinguiba, F.; Vogt, T. Identification and Characterization of Piperine Synthase from Black Pepper, Piper nigrum L. Commun. Biol. 2021, 4, 445. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Qian, D.; Du, Q. Preparation of Bioactive Amide Compounds from Black Pepper by Countercurrent Chromatography and Preparative HPLC. Ind. Crops Prod. 2013, 44, 258–262. [Google Scholar] [CrossRef]
- Gómez-Calvario, V.; Rios, M.Y. 1H and 13C NMR Data, Occurrence, Biosynthesis, and Biological Activity of Piper Amides. Magn. Reson. Chem. 2019, 57, 994–1070. [Google Scholar] [CrossRef]
- Schnabel, A.; Cotinguiba, F.; Athmer, B.; Vogt, T. Piper nigrum Cyp719a37 Catalyzes the Decisive Methylenedioxy Bridge Formation in Piperine Biosynthesis. Plants 2021, 10, 128. [Google Scholar] [CrossRef]
- Vassão, D.G.; Davin, L.B.; Lewis, N.G. Metabolic Engineering of Plant Allyl/Propenyl Phenol and Lignin Pathways: Future Potential for Biofuels/Bioenergy, Polymer Intermediates, and Specialty Chemicals? Adv. Plant Biochem. Mol. Biol. 2008, 1, 385–428. [Google Scholar]
- Zeng, Y.; Qin, B.; Shi, Y.W.; Long, Y.S.; Deng, W.Y.; Li, B.M.; Tang, B.; Zhao, Q.H.; Gao, M.M.; He, N.; et al. Ilepcimide Inhibited Sodium Channel Activity in Mouse Hippocampal Neurons. Epilepsy Res. 2021, 170, 106533. [Google Scholar] [CrossRef]
- Lee, S.W.; Kim, Y.K.; Kim, K.; Lee, H.S.; Choi, J.H.; Lee, W.S.; Jun, C.D.; Park, J.H.; Lee, J.M.; Rho, M.C. Alkamides from the Fruits of Piper longum and Piper nigrum Displaying Potent Cell Adhesion Inhibition. Bioorganic. Med. Chem. Lett. 2008, 18, 4544–4546. [Google Scholar] [CrossRef]
- Rohm, B.; Riedel, A.; Ley, J.P.; Widder, S.; Krammer, G.E.; Somoza, V. Capsaicin, Nonivamide and Trans-Pellitorine Decrease Free Fatty Acid Uptake without TRPV1 Activation and Increase Acetyl-Coenzyme A Synthetase Activity in Caco-2 Cells. Food Funct. 2015, 6, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Lieder, B.; Zaunschirm, M.; Holik, A.K.; Ley, J.P.; Hans, J.; Krammer, G.E.; Somoza, V. The Alkamide Trans-Pellitorine Targets PPARγ via TRPV1 and TRPA1 to Reduce Lipid Accumulation in Developing 3T3-L1 Adipocytes. Front. Pharmacol. 2017, 8, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Alstyne, K.L.; Nelson, A.V.; Vyvyan, J.R.; Cancilla, D.A. Dopamine Functions as an Antiherbivore Defense in the Temperate Green Alga Ulvaria obscura. Oecologia 2006, 148, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.I.; Ozawa, R.; Kugimiya, S.; Takabayashi, J.; Bohlmann, J. Herbivore-Induced Defense Response in a Model Legume. Two-Spotted Spider Mites Induce Emission of (E)-β-Ocimene and Transcript Accumulation of (E)-β-Ocimene Synthase in Lotus japonicus. Plant Physiol. 2004, 135, 1976–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of Secondary Metabolites in Plant Defense against Pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Hodge, S.; Keesing, V.; Wratten, S.D.; Lövei, G.L.; Palmer, J.; Çilgi, T. Herbivore Damage and Leaf Loss in the New Zealand Pepper Tree (“kawakawa”; Macropiper excelsum; Piperaceae). N. Z. J. Ecol. 1998, 22, 173–180. [Google Scholar]
- FSANZ. Novel Foods—Record of Views Formed in Response to Inquiries; Recommended by the Advisory Committee on Novel Foods; FSANZ: Canberra, Australia, 2022.
- Hallström, H.; Thuvander, A. Toxicological Evaluation of Myristicin. Nat. Toxins 1997, 5, 186–192. [Google Scholar] [CrossRef]
- Yang, X.N.; Wang, Y.K.; Zhu, X.; Xiao, X.R.; Dai, M.Y.; Zhang, T.; Qu, Y.; Yang, X.W.; Qin, H.B.; Gonzalez, F.J.; et al. Metabolic Activation of Elemicin Leads to the Inhibition of Stearoyl-CoA Desaturase 1. Chem. Res. Toxicol. 2019, 32, 1965–1976. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Rossi, P.G.; Bao, L.; Luciani, A.; Panighi, J.; Desjobert, J.M.; Costa, J.; Casanova, J.; Bolla, J.M.; Berti, L. (E)-Methylisoeugenol and Elemicin: Antibacterial Components of Daucus carota L. Essential Oil against Campylobacter jejuni. J. Agric. Food Chem. 2007, 55, 7332–7336. [Google Scholar] [CrossRef]
- Flamini, G.; Cosimi, E.; Cioni, P.L.; Molfetta, I.; Braca, A. Essential-Oil Composition of Daucus carota Ssp. Major (Pastinocello Carrot) and Nine Different Commercial Varieties of Daucus carota Ssp. Sativus Fruits. Chem. Biodivers. 2014, 11, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Veryser, L.; Bracke, N.; Wynendaele, E.; Joshi, T.; Tatke, P.; Taevernier, L.; De Spiegeleer, B. Quantitative in Vitro and in Vivo Evaluation of Intestinal and Blood-Brain Barrier Transport Kinetics of the Plant N -Alkylamide Pellitorine. BioMed Res. Int. 2016, 2016, 5497402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farsi, E.; Shafaei, A.; Hor, S.Y.; Ahamed, M.B.K.; Yam, M.F.; Asmawi, M.Z.; Ismail, Z. Genotoxicity and Acute and Subchronic Toxicity Studies of a Standardized Methanolic Extract of Ficus deltoidea Leaves. Clinics 2013, 68, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Castro-Faria-Neto, H.C.; Bozza, P.T.; Cruz, H.N.; Silva, C.L.M.; Violante, F.A.; Barbosa-Filho, J.M.; Thomas, G.; Martins, M.A.; Tibirica, E.V.; Noel, F.; et al. Yangambin: A New Naturally-Occurring Platelet-Activating Factor Receptor Antagonist: Binding and in Vitro Functional Studies. Planta Med. 1995, 61, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, D.Y.; Zhang, Z.H.; Zuo, C.H.; Zhang, Y.; Pei, Y.Q.; Lo, Y.Q. Trial of Antiepilepsirine (AES) in Children with Epilepsy. Brain Dev. 1999, 21, 36–40. [Google Scholar] [CrossRef]
- Ley, J.P.; Hilmer, J.M.; Weber, B.; Krammer, G.; Gatfield, I.L.; Bertram, H.J. Stereoselective Enzymatic Synthesis of Cis-Pellitorine, a Taste Active Alkamide Naturally Occurring in Tarragon. Eur. J. Org. Chem. 2004, 2004, 5135–5140. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Date Collected/Acquired | Kawakawa Source | Extraction | Provenance |
|---|---|---|---|---|
| MCO | 2020 | ŌKU | 80% methanol | commercial |
| MFA | Late 2021 | Arapaoa Island | 80% methanol | fresh |
| MFP | Late 2021 | Pōhara | 80% methanol | fresh |
| ACO | 2020 | ŌKU | aqueous | commercial |
| ACP | 2020 | Phytomed | aqueous | commercial |
| AFT1 | Early January 2021 | Te Tauihu | aqueous | fresh |
| AFT2 | Late January 2021 | Te Tauihu | aqueous | fresh |
| AFT3 | Early March 2021 | Te Tauihu | aqueous | fresh |
| AFT4 | Late March 2021 | Te Tauihu | aqueous | fresh |
| AFT5 | April 2021 | Te Tauihu | aqueous | fresh |
| AFM | October 2020 | Mohua, Golden Bay | aqueous | fresh |
| ID | Annotations [Empirical Observations] | Chemical Formula | RT (min) | Observed m/z | Mass Accuracy (ppm) | Major MS/MS Ions |
|---|---|---|---|---|---|---|
| 1 | salsolinol | C10H13NO2 | 3.28 | 180.1022 | −1.28 | 85.0283, 145.0647, 163.0753, 180.1022 |
| 2 | trigonelline | C7H7NO2 | 1.65 | 138.0550 | −3.62 | 81.0698, 95.0854, 138.0555 |
| 3 | dopamine | C8H11NO2 | 2.15 | 154.0868 | 0.00 | 91.0542, 137.0596, 154.0868 |
| 4 | vitexin-O-glucoside [4] | C27H30O15 | 5.08 | 595.1666 | 0.55 | 271.0597, 313.0705, 337.0709, 397.0927, 415.1015, 433.1126, 595.1666 |
| 5 | kaempferol malonyl neohesperidoside | C30H32O18 | 5.23 | 681.1676 | 0.56 | 271.0603, 313.0706, 415.1021, 433.1125, 561.1249, 681.1676 |
| 6 | isorhamnetin galactoside rhamnoside | C28H32O16 | 5.28 | 625.1776 | 1.47 | 301.0703, 343.0812, 427.1017, 445.1124, 463.1235, 625.1776 |
| 7 | vitexin | C21H20O10 | 5.32 | 433.1129 | 1.20 | 271.0606, 313.0712, 337.0712, 397.0908, 415.1024, 433.1129 |
| 8 | hydroxycinnamoyl pyrrolidinol | C13H15NO3 | 5.41 | 234.1134 | −1.27 | 131.0491, 234.1134 |
| 9 | malonylgenistin | C24H22O13 | 5.60 | 519.1140 | 1.67 | 283.0595, 295.0602, 313.0702, 519.1140 |
| 10 | N-methyldopamine | C9H13NO2 | 5.82 | 168.1020 | 0.40 | 91.0541, 119.0490, 151.0752, 170.0961 |
| 11 | awaine [23] | C14H17NO2 | 6.62, 6.94, 7.08 | 232.1342 | −2.32 | 82.0654, 105.0702, 133.0652, 150.0918, 232.1331 |
| 12 | piperlotine-K | C13H17NO3 | 5.93 | 236.1291 | 2.11 | 86.0600, 105.0698, 133.0645, 235.1691 |
| 13 | cinnamoyl hydroxypyrrolidine | C13H15NO2 | 6.13 | 218.1185 | 1.83 | 131.0490, 218.1174 |
| 14 | cinnamoyl piperidone (annotated as piperchabamide A) [19] | C14H15NO2 | 6.22, 7.04 | 230.1184 | 2.12 | 82.0653, 100.0706, 131.0494 |
| 15 | tryptophan | C11H12N2O2 | 6.27 | 205.0976 | 1.83 | 146.0598, 159.0914, 188.0704 |
| 16 | cinnamoyl piperideine | C14H15NO | 8.91, 9.44 | 214.1228 | 1.39 | 84.0807, 131.0490, 214.1223 |
| 17 | saturated fagaramide | C14H19NO3 | 7.17 | 250.1443 | −0.04 | 82.0651, 93.0366, 155.0853, 177.0545, 229.1034, 250.1443 |
| 18 | saturated ilepcimide | C15H19NO3 | 7.36 | 262.1442 | −0.59 | 82.0651, 145.0283, 177.0546, 262.1438 |
| 19 | methylenedioxycinnamoyl pyrrolidine | C14H15NO3 | 7.41, 7.82 | 246.1130 | −1.40 | 82.0651, 145.0282, 175.0388, 246.1130 |
| 20 | orientin-O-glucoside [4] | C27H30O16 | 7.55 | 611.1619 | −0.04 | 329.0663, 431.0981, 449.1084 |
| 21 | kaempferol malonylglucoside glucoside | C30H32O19 | 7.64 | 697.1620 | −0.27 | 127.0389, 287.0548, 329.0654, 431.0967, 554.4190 |
| 22 | dimethoxycinnamoyl piperidine | C16H21NO3 | 7.74, 8.16 | 276.1604 | −0.04 | 84.0807, 112.0755, 191.0700, 216.1378, 276.1599 |
| 23 | cinnamoyl pyrrolidine | C13H15NO | 7.89 | 202.1232 | 1.15 | 131.0491, 161.0959, 181.0107, 202.1225 |
| 24 | fagaramide [4] | C14H17NO3 | 8.15 (cis), 8.40 (trans) | 248.1287 | 0.24 | 175.0395, 248.1289 |
| 25 | ilepcimide [4] | C15H17NO3 | 8.30, 8.70 | 260.1289 | 0.56 | 175.0394, 260.1288 |
| 26 | cinnamoyl piperidine | C14H17NO | 8.43, 8.82 | 216.1387 | 1.70 | 131.0494, 216.1389 |
| 27 | methoxycinnamoyl piperidine | C15H19NO2 | 8.48, 8.86 | 246.1494 | 0.35 | 82.0650, 112.0755, 161.0596, 246.1494 |
| 28 | piperlonguminine | C16H19NO3 | 8.75 | 274.1444 | 0.24 | 115.0534, 135.0439, 201.0544, 274.1443 |
| 29 | cinnamoyl isobutylamine | C13H17NO | 8.51 | 204.1387 | −0.49 | 116.0529, 131.0491, 148.0755, 204.1388 |
| 30 | dihydrocinnamoyl hydroxypiperidine | C14H19NO2 | 8.55, 8.76 | 234.1499 | 1.08 | 82.0652, 135.0802, 161.0596, 234.1499 |
| 31 | dicinnamoyl putrescine | C22H24N2O2 | 8.75 | 349.1931 | 4.3 | 72.0807, 131.0490, 202.1225, 219.1492, 349.1931 |
| 32 | yangambin, diayangambin, epiyangambin | C24H30O8 | 9.11, 9.32, 9.40 | 447.2023 | −0.60 | 181.0864, 204.0787, 219.1021, 231.1022, 358.1422 |
| 33 | magnolin | C23H28O7 | 9.17 | 417.1911 | −0.12 | 135.0349, 151.0800, 189.0900, 193.0864, 328.13, 399.1797 |
| 34 | dihydropiperlonguminine | C16H21NO3 | 9.18 | 276.1600 | 0.29 | 135.0444, 276.1603 |
| 35 | methylenedioxycinnamoyl isopentylamine | C15H19NO3 | 9.18 | 262.1441 | 0.20 | 145.0283, 175.0389, 262.1438 |
| 36 | ilepcimide piperideine | C15H15NO3 | 9.30 | 258.1129 | 2.26 | 167.0704, 175.0394, 199.01329, 257.1282, 258.1130 |
| 37 | elemicin [3] | C12H16O3 | 9.33 | 209.1173 | 4.24 | 168.0785, 194.0942, 209.1178 |
| 38 | N-isobutyl octadienamide | C12H21NO | 9.40 | 196.1699 | 1.12 | 123.0804, 181.1222 |
| 39 | piperamide-C7:2 | C18H21NO3 | 9.40 | 300.1592 | −0.48 | 74.0963, 135.0439, 302.1747 |
| 40 | piperine [4] | C17H19NO3 | 9.59, 9.70 | 286.1444 | 0.25 | 201.0550, 286.1444 |
| 41 | piperanine | C17H21NO3 | 9.43 | 288.1599 | −0.76 | 86.0966, 135.0443, 288.1601 |
| 42 | chingchengenamide A [22] | C18H23NO3 | 9.69 | 302.1759 | −0.58 | 135.0444, 302.1761 |
| 43 | episesartemin [4] | C23H26O8 | 9.83 | 431.1709 | −1.96 | 216.1388 |
| 44 | myristicin [4] | C11H12O3 | 10.05 | 193.0863 | 0.88 | 135.0808, 152.0471, 163.0757, 193.0864 |
| 45 | piperdardine | C19H23NO3 | 10.45 | 314.1758 | −0.82 | 135.0443, 314.1758 |
| 46 | sarmentine | C14H23NO | 10.63 | 222.1858 | −2.40 | 98.0600, 133.1010, 151.1116, 168.1381, 222.1857 |
| 47 | piperolein A | C19H25NO3 | 10.71 | 316.1917 | 0.17 | 86.0963, 135.0439, 173.0951, 316.1906 |
| 48 | pellitorine [4] | C14H25NO | 10.92 | 224.2013 | 0.62 | 209.1653, 224.2012, 227.1758 |
| 49 | neopellitorine B (annotated as achilleamide) [4] | C15H25NO | 11.37, 11.49 | 236.2011 | 0.03 | 123.0808, 151.1122, 236.2015 |
| 50 | kalecide [4] | C16H29NO | 12.33 | 252.2327 | 0.99 | 123.0807, 179.1071, 195.1013, 252.2328 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayaprakash, R.; Ramzan, F.; Miles-Chan, J.L.; Foster, M.; Mithen, R.F.; Pook, C. Exploring the Chemical Space of Kawakawa Leaf (Piper excelsum). Nutrients 2022, 14, 5168. https://doi.org/10.3390/nu14235168
Jayaprakash R, Ramzan F, Miles-Chan JL, Foster M, Mithen RF, Pook C. Exploring the Chemical Space of Kawakawa Leaf (Piper excelsum). Nutrients. 2022; 14(23):5168. https://doi.org/10.3390/nu14235168
Chicago/Turabian StyleJayaprakash, Ramya, Farha Ramzan, Jennifer L. Miles-Chan, Meika Foster, Richard F. Mithen, and Chris Pook. 2022. "Exploring the Chemical Space of Kawakawa Leaf (Piper excelsum)" Nutrients 14, no. 23: 5168. https://doi.org/10.3390/nu14235168