
Benefits and Risks of Early Life Iron Supplementation


Abstract
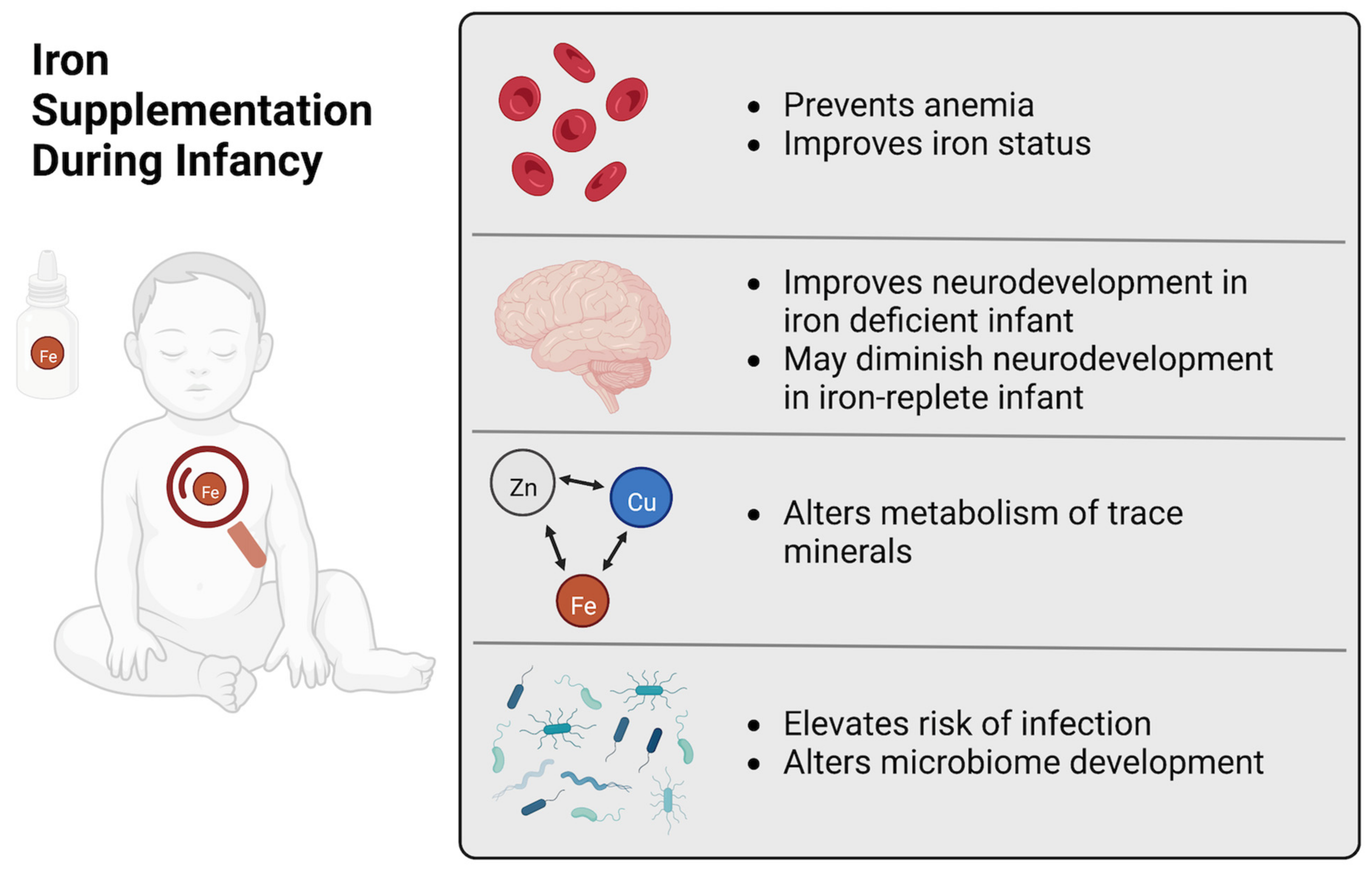
:1. Introduction
2. Deficiency & Toxicity
2.1. Defining Anemia
2.2. Defining Iron Deficiency
2.3. Iron Supplementation, Iron Status, & Hematology
2.4. Developmental Regulation of Iron
2.5. Oxidative Stress Results from Iron Overload
3. Growth & Development
3.1. Growth Effects of Dietary Iron Excess
3.2. Neurodevelopmental Outcomes of Iron Supplementation
3.3. Mechanisms Underlying Neurodevelopmental Effects of Iron Supplementation
4. Trace Mineral Interactions
4.1. Iron Deficiency May Mask Copper or Zinc Deficiency
4.2. Iron Competes with Other Trace Minerals for Absorption & Metabolism
4.3. Trace Minerals and Oxidative Stress
5. Morbidity & Mortality
5.1. Iron Affects Morbidity & Mortality of Infants & Children
5.2. Gut Development & the Gut Microbiota
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, J.L. Iron Deficiency Anemia: A Common and Curable Disease. Cold Spring Harb. Perspect. Med. 2013, 3, a011866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, R.; Leon, J.; Suchdev, P. Identification, Prevention and Treatment of Iron Deficiency during the First 1000 Days. Nutrients 2014, 6, 4093–4114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Lönnerdal, B.; Adu-Afarwuah, S.; Brown, K.H.; Chaparro, C.M.; Cohen, R.J.; Domellöf, M.; Hernell, O.; Lartey, A.; Dewey, K.G. Prevalence and Predictors of Iron Deficiency in Fully Breastfed Infants at 6 Mo of Age: Comparison of Data from 6 Studies. Am. J. Clin. Nutr. 2009, 89, 1433–1440. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Iron Deficiency Anaemia: Assessment, Prevention, and Control. A Guide for Programme Managers; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Shelov, S.P. American Academy of Pediatrics. Caring for Your Baby and Young Child: Birth to Age Five; Bantam: New York, NY, USA, 2009; ISBN 978-0-553-38630-1. [Google Scholar]
- Lozoff, B.; Beard, J.; Connor, J.; Barbara, F.; Georgieff, M.; Schallert, T. Long-Lasting Neural and Behavioral Effects of Iron Deficiency in Infancy. Nutr. Rev. 2006, 64, S34–S43; discussion S72–S91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- East, P.; Doom, J.R.; Blanco, E.; Burrows, R.; Lozoff, B.; Gahagan, S. Iron Deficiency in Infancy and Neurocognitive and Educational Outcomes in Young Adulthood. Dev. Psychol. 2021, 57, 962–975. [Google Scholar] [CrossRef] [PubMed]
- DRI. Dietary. Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc: A Report of the Panel on Micronutrients ... and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes, Food and Nutrition Board, Institute of Medicine; Institute of Medicine (U.S.), Ed.; National Academy Press: Washington, DC, USA, 2001; ISBN 978-0-309-07279-3. [Google Scholar]
- Domellöf, M.; Lönnerdal, B.; Abrams, S.A.; Hernell, O. Iron Absorption in Breast-Fed Infants: Effects of Age, Iron Status, Iron Supplements, and Complementary Foods. Am. J. Clin. Nutr. 2002, 76, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Lönnerdal, B. Development of Iron Homeostasis in Infants and Young Children. Am. J. Clin. Nutr. 2017, 106, 1575S–1580S. [Google Scholar] [CrossRef] [Green Version]
- Lönnerdal, B.; Georgieff, M.K.; Hernell, O. Developmental Physiology of Iron Absorption, Homeostasis, and Metabolism in the Healthy Term Infant. J. Pediatr. 2015, 167, S8–S14. [Google Scholar] [CrossRef] [Green Version]
- Lönnerdal, B. Iron, Zinc, Copper, and Manganese in Infant Formulas. Arch. Pediatr. Adolesc. Med. 1983, 137, 433. [Google Scholar] [CrossRef]
- Lonnerdal, B. Excess Iron Intake as a Factor in Growth, Infections, and Development of Infants and Young Children. Am. J. Clin. Nutr. 2017, 106, 1681S–1687S. [Google Scholar] [CrossRef] [Green Version]
- Wessling-Resnick, M. Excess Iron: Considerations Related to Development and Early Growth. Am. J. Clin. Nutr. 2017, 106, 1600S–1605S. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Berggren, K.L.; Marks, E.; Fox, J.H. Impact of High Iron Intake on Cognition and Neurodegeneration in Humans and in Animal Models: A Systematic Review. Nutr. Rev. 2017, 75, 456–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganini, D.; Zimmermann, M.B. The Effects of Iron Fortification and Supplementation on the Gut Microbiome and Diarrhea in Infants and Children: A Review. Am. J. Clin. Nutr. 2017, 106, 1688S–1693S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanchi, A.; James, P.T.; Cerami, C. Guts, Germs, and Iron: A Systematic Review on Iron Supplementation, Iron Fortification, and Diarrhea in Children Aged 4–59 Months. Curr. Dev. Nutr. 2019, 3, nzz005. [Google Scholar] [CrossRef] [Green Version]
- Allen, L.H. Iron Supplements: Scientific Issues Concerning Efficacy and Implications for Research and Programs. J. Nutr. 2002, 132, 813S–819S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietary Guidelines Advisory Committee. Scientific Report of the 2020 Dietary Guidelines Advisory Committee: Advisory Report to the Secretary of Agriculture and the Secretary of Health and Human Services; U.S. Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 2020; p. 786. [Google Scholar]
- Pasricha, S.-R.; Hayes, E.; Kalumba, K.; Biggs, B.-A. Effect of Daily Iron Supplementation on Health in Children Aged 4–23 Months: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Lancet Glob. Health 2013, 1, e77–e86. [Google Scholar] [CrossRef] [Green Version]
- Guideline: Daily Iron Supplementation in Infants and Children; World Health Organization: Geneva, Switzerland, 2016; ISBN 978-92-4-154952-3.
- Dewey, K.G.; Domellöf, M.; Cohen, R.J.; Landa Rivera, L.; Hernell, O.; Lönnerdal, B. Iron Supplementation Affects Growth and Morbidity of Breast-Fed Infants: Results of a Randomized Trial in Sweden and Honduras. J. Nutr. 2002, 132, 3249–3255. [Google Scholar] [CrossRef] [Green Version]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron Fortification Adversely Affects the Gut Microbiome, Increases Pathogen Abundance and Induces Intestinal Inflammation in Kenyan Infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef]
- Lind, T.; Seswandhana, R.; Persson, L.-A.; Lönnerdal, B. Iron Supplementation of Iron-Replete Indonesian Infants Is Associated with Reduced Weight-for-Age. Acta Paediatr. 2008, 97, 770–775. [Google Scholar] [CrossRef]
- Paganini, D.; Uyoga, M.A.; Kortman, G.A.M.; Cercamondi, C.I.; Winkler, H.C.; Boekhorst, J.; Moretti, D.; Lacroix, C.; Karanja, S.; Zimmermann, M.B. Iron-Containing Micronutrient Powders Modify the Effect of Oral Antibiotics on the Infant Gut Microbiome and Increase Post-Antibiotic Diarrhoea Risk: A Controlled Study in Kenya. Gut 2019, 68, 645–653. [Google Scholar] [CrossRef]
- Domellöf, M.; Dewey, K.G.; Cohen, R.J.; Lönnerdal, B.; Hernell, O. Iron Supplements Reduce Erythrocyte Copper-Zinc Superoxide Dismutase Activity in Term, Breastfed Infants. Acta Paediatr. 2005, 94, 1578–1582. [Google Scholar] [CrossRef]
- World Health Organization. Hemoglobin Concentrations for the Diagnosis of Anemia and Assessment of Severity; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Nutritional Anemias. Report of a WHO Scientific Group. 1968. Available online: https://apps.who.int/iris/handle/10665/40707 (accessed on 26 July 2022).
- Sturgeon, P. Studies of Iron Requirements in Infants and Children. Pediatrics 1954, 13, 107–125. [Google Scholar] [CrossRef]
- Sturgeon, P. Studies of Iron Requirements in Infants III. Influence of Supplemental Iron during Normal Pregnancy on Mother and Infant B. The Infant. Br. J. Haematol 1959, 5, 45–55. [Google Scholar] [CrossRef]
- Center for Disease Control & Prevention. CDC Criteria for Anemia in Children and Childbearing-Aged Women. Morb. Mortal. Wkly. Rep. 1989, 38, 400–404. [Google Scholar]
- Pilch, S.M.; Senti, F.R.; Assessment of the Iron Nutritional Status of the U.S. Population Based on Data Collected in the Second National Health and Nutrition Examination Survey. 1984. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300391414 (accessed on 19 August 2022).
- Baker, R.D.; Greer, F.R. Diagnosis and Prevention of Iron Deficiency and Iron-Deficiency Anemia in Infants and Young Children (0–3 Years of Age). Pediatrics 2010, 126, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Serum Ferritin Concentrations for the Assessment of Iron Status and Iron Deficiency in Populations; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Garcia-Casal, M.N.; Pasricha, S.-R.; Martinez, R.X.; Lopez-Perez, L.; Peña-Rosas, J.P. Serum or Plasma Ferritin Concentration as an Index of Iron Deficiency and Overload. Cochrane Database Syst. Rev. 2021, 2021, CD011817. [Google Scholar] [CrossRef]
- Hare, D.J.; Braat, S.; Cardoso, B.R.; Morgan, C.; Szymlek-Gay, E.A.; Biggs, B.-A. Health Outcomes of Iron Supplementation and/or Food Fortification in Iron-Replete Children Aged 4–24 Months: Protocol for a Systematic Review and Meta-Analysis. Syst. Rev. 2019, 8, 253. [Google Scholar] [CrossRef]
- Nutrient Requirements of Swine, 11th ed.; National Research Council, (U.S.) (Ed.) National Academies Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- Dong, Z.; Wan, D.; Li, G.; Zhang, Y.; Yang, H.; Wu, X.; Yin, Y. Comparison of Oral and Parenteral Iron Administration on Iron Homeostasis, Oxidative and Immune Status in Anemic Neonatal Pigs. Biol. Trace Elem. Res. 2020, 195, 117–124. [Google Scholar] [CrossRef]
- Ji, P.; Lonnerdal, B.; Kim, K.; Jinno, C.N. Iron Oversupplementation Causes Hippocampal Iron Overloading and Impairs Social Novelty Recognition in Nursing Piglets. J. Nutr. 2019, 149, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Egeli, A.K.; Framstad, T. Effect of an Oral Starter Dose of Iron on Haematology and Weight Gain in Piglets Having Voluntary Access to Glutamic Acid-Chelated Iron Solution. Acta Vet. Scand. 1998, 39, 359–365. [Google Scholar] [CrossRef]
- Furugouri, K.; Kawabata, A. Iron Absorption in Nursing Piglets. J. Anim. Sci. 1975, 41, 1348–1354. [Google Scholar] [CrossRef]
- Loh, T.J.; Leong, K.; Too, H.; Mah, C.; Choo, P. The Effects of Iron Supplementation in Preweaning Piglets. Malays. J. Nutr. 2001, 7, 41–49. [Google Scholar]
- Webster, W.R.; Dimmock, C.K.; O’Rourke, P.K.; Lynch, P.J. Evaluation of Oral Iron Galactan as a Method of Iron Supplementation for Intensively Housed Sucking Piglets. Aust. Vet. J. 1978, 54, 345–348. [Google Scholar] [CrossRef]
- Perng, V.; Li, C.; Klocke, C.R.; Navazesh, S.E.; Pinneles, D.K.; Lein, P.J.; Ji, P. Iron Deficiency and Iron Excess Differently Affect Dendritic Architecture of Pyramidal Neurons in the Hippocampus of Piglets. J. Nutr. 2021, 151, 235–244. [Google Scholar] [CrossRef]
- Leong, W.-I.; Bowlus, C.L.; Tallkvist, J.; Lönnerdal, B. DMT1 and FPN1 Expression during Infancy: Developmental Regulation of Iron Absorption. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 285, G1153–G1161. [Google Scholar] [CrossRef] [Green Version]
- Leong, W.-I.; Bowlus, C.L.; Tallkvist, J.; Lönnerdal, B. Iron Supplementation during Infancy—Effects on Expression of Iron Transporters, Iron Absorption, and Iron Utilization in Rat Pups. Am. J. Clin. Nutr. 2003, 78, 1203–1211. [Google Scholar] [CrossRef] [Green Version]
- Alexeev, E.E.; He, X.; Slupsky, C.M.; Lönnerdal, B. Effects of Iron Supplementation on Growth, Gut Microbiota, Metabolomics and Cognitive Development of Rat Pups. PLoS ONE 2017, 12, e0179713. [Google Scholar] [CrossRef] [Green Version]
- McMillen, S.; Lönnerdal, B. Postnatal Iron Supplementation with Ferrous Sulfate vs. Ferrous Bis-Glycinate Chelate: Effects on Iron Metabolism, Growth, and Central Nervous System Development in Sprague Dawley Rat Pups. Nutrients 2021, 13, 1406. [Google Scholar] [CrossRef]
- Kaur, D.; Peng, J.; Chinta, S.J.; Rajagopalan, S.; Di Monte, D.A.; Cherny, R.A.; Andersen, J.K. Increased Murine Neonatal Iron Intake Results in Parkinson-like Neurodegeneration with Age. Neurobiol. Aging 2007, 28, 907–913. [Google Scholar] [CrossRef]
- Fredriksson, A.; Archer, T. Subchronic Administration of Haloperidol Influences the Functional Deficits of Postnatal Iron Administration in Mice. Neurotox. Res. 2006, 10, 123–129. [Google Scholar] [CrossRef]
- Fredriksson, A.; Archer, T. Effect of Postnatal Iron Administration on MPTP-Induced Behavioral Deficits and Neurotoxicity: Behavioral Enhancement by L-Dopa-MK-801 Co-Administration. Behav. Brain Res. 2003, 139, 31–46. [Google Scholar] [CrossRef]
- Fredriksson, A.; Archer, T. Postnatal Iron Overload Destroys NA-DA Functional Interactions. J. Neural Transm. 2007, 114, 195–203. [Google Scholar] [CrossRef]
- Berggren, K.L.; Lu, Z.; Fox, J.A.; Dudenhoeffer, M.; Agrawal, S.; Fox, J.H. Neonatal Iron Supplementation Induces Striatal Atrophy in Female YAC128 Huntington’s Disease Mice. J. Huntingt. Dis. 2016, 5, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Piñero, D.J.; Li, N.-Q.; Connor, J.R.; Beard, J.L. Variations in Dietary Iron Alter Brain Iron Metabolism in Developing Rats. J. Nutr. 2000, 130, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Y.; Babitt, J.L. Liver Iron Sensing and Body Iron Homeostasis. Blood 2019, 133, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Billesbølle, C.B.; Azumaya, C.M.; Kretsch, R.C.; Powers, A.S.; Gonen, S.; Schneider, S.; Arvedson, T.; Dror, R.O.; Cheng, Y.; Manglik, A. Structure of Hepcidin-Bound Ferroportin Reveals Iron Homeostatic Mechanisms. Nature 2020, 586, 807–811. [Google Scholar] [CrossRef]
- Nemeth, E. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [Green Version]
- Frazer, D.M.; Wilkins, S.J.; Darshan, D.; Mirciov, C.S.G.; Dunn, L.A.; Anderson, G.J. Ferroportin Is Essential for Iron Absorption During Suckling, But Is Hyporesponsive to the Regulatory Hormone Hepcidin. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Darshan, D.; Wilkins, S.J.; Frazer, D.M.; Anderson, G.J. Reduced Expression of Ferroportin-1 Mediates Hyporesponsiveness of Suckling Rats to Stimuli That Reduce Iron Absorption. Gastroenterology 2011, 141, 300–309. [Google Scholar] [CrossRef]
- Chen, X.; Yu, C.; Kang, R.; Tang, D. Iron Metabolism in Ferroptosis. Front. Cell Dev. Biol. 2020, 8, 590226. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.; Huang, Z.; Lin, Z.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, Present and Future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef]
- Niederau, C.; Fischer, R.; Sonnenberg, A.; Stremmel, W.; Trampisch, H.J.; Strohmeyer, G. Survival and Causes of Death in Cirrhotic and in Noncirrhotic Patients with Primary Hemochromatosis. N. Engl. J. Med. 1985, 313, 1256–1262. [Google Scholar] [CrossRef]
- Deugnier, Y.M.; Loréal, O.; Turlin, B.; Guyader, D.; Jouanolle, H.; Moirand, R.; Jacquelinet, C.; Brissot, P. Liver Pathology in Genetic Hemochromatosis: A Review of 135 Homozygous Cases and Their Bioclinical Correlations. Gastroenterology 1992, 102, 2050–2059. [Google Scholar] [CrossRef]
- Houglum, K.; Ramm, G.A.; Crawford, D.H.; Witztum, J.L.; Powell, L.W.; Chojkier, M. Excess Iron Induces Hepatic Oxidative Stress and Transforming Growth Factor? 1 in Genetic Hemochromatosis. Hepatology 1997, 26, 605–610. [Google Scholar] [CrossRef]
- Lönnerdal, B.; Hernell, O. Iron, Zinc, Copper and Selenium Status of Breast-Fed Infants and Infants Fed Trace Element Fortified Milk-Based Infant Formula. Acta Paediatr. 1994, 83, 367–373. [Google Scholar] [CrossRef]
- Budni, P.; de Lima, M.N.M.; Polydoro, M.; Moreira, J.C.F.; Schroder, N.; Dal-Pizzol, F. Antioxidant Effects of Selegiline in Oxidative Stress Induced by Iron Neonatal Treatment in Rats. Neurochem. Res. 2007, 32, 965–972. [Google Scholar] [CrossRef]
- Chen, H.; Wang, X.; Wang, M.; Yang, L.; Yan, Z.; Zhang, Y.; Liu, Z. Behavioral and Neurochemical Deficits in Aging Rats with Increased Neonatal Iron Intake: Silibinin’s Neuroprotection by Maintaining Redox Balance. Front. Aging Neurosci. 2015, 7, 206. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Fox, J.; Thyagarajan, B.; Fox, J.H. Brain Mitochondrial Iron Accumulates in Huntington’s Disease, Mediates Mitochondrial Dysfunction, and Can Be Removed Pharmacologically. Free Radic. Biol. Med. 2018, 120, 317–329. [Google Scholar] [CrossRef]
- Fernandez, L.L.; Carmona, M.; Portero-Otin, M.; Naudi, A.; Pamplona, R.; Schröder, N.; Ferrer, I. Effects of Increased Iron Intake during the Neonatal Period on the Brain of Adult AbetaPP/PS1 Transgenic Mice. J. Alzheimers Dis. 2010, 19, 1069–1080. [Google Scholar] [CrossRef]
- Dal-Pizzol, F.; Klamt, F.; Frota, M.L.J.; Andrades, M.E.; Caregnato, F.F.; Vianna, M.M.; Schröder, N.; Quevedo, J.; Izquierdo, I.; Archer, T.; et al. Neonatal Iron Exposure Induces Oxidative Stress in Adult Wistar Rat. Brain Res. Dev. Brain Res. 2001, 130, 109–114. [Google Scholar] [CrossRef]
- Yu, L.; Wang, X.; Chen, H.; Yan, Z.; Wang, M.; Li, Y. Neurochemical and Behavior Deficits in Rats with Iron and Rotenone Co-Treatment: Role of Redox Imbalance and Neuroprotection by Biochanin A. Front. Neurosci. 2017, 11, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima, M.N.M.; Polydoro, M.; Laranja, D.C.; Bonatto, F.; Bromberg, E.; Moreira, J.C.F.; Dal-Pizzol, F.; Schröder, N. Recognition Memory Impairment and Brain Oxidative Stress Induced by Postnatal Iron Administration. Eur. J. Neurosci. 2005, 21, 2521–2528. [Google Scholar] [CrossRef]
- Lavich, I.C.; de Freitas, B.S.; Kist, L.W.; Falavigna, L.; Dargél, V.A.; Köbe, L.M.; Aguzzoli, C.; Piffero, B.; Florian, P.Z.; Bogo, M.R.; et al. Sulforaphane Rescues Memory Dysfunction and Synaptic and Mitochondrial Alterations Induced by Brain Iron Accumulation. Neuroscience 2015, 301, 542–552. [Google Scholar] [CrossRef]
- Dijkhuizen, M.A.; Winichagoon, P.; Wieringa, F.T.; Wasantwisut, E.; Utomo, B.; Ninh, N.X.; Hidayat, A.; Berger, J. Zinc Supplementation Improved Length Growth Only in Anemic Infants in a Multi-Country Trial of Iron and Zinc Supplementation in South-East Asia. J. Nutr. 2008, 138, 1969–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petry, N.; Olofin, I.; Boy, E.; Donahue Angel, M.; Rohner, F. The Effect of Low Dose Iron and Zinc Intake on Child Micronutrient Status and Development during the First 1000 Days of Life: A Systematic Review and Meta-Analysis. Nutrients 2016, 8, 773. [Google Scholar] [CrossRef] [Green Version]
- Björmsjö, M.; Hernell, O.; Lönnerdal, B.; Berglund, S.K. Reducing Iron Content in Infant Formula from 8 to 2 Mg/L Does Not Increase the Risk of Iron Deficiency at 4 or 6 Months of Age: A Randomized Controlled Trial. Nutrients 2020, 13, 3. [Google Scholar] [CrossRef]
- Lozoff, B. Iron-Fortified vs Low-Iron Infant Formula: Developmental Outcome at 10 Years. Arch. Pediatr. Adolesc. Med. 2012, 166, 208. [Google Scholar] [CrossRef] [Green Version]
- Gahagan, S.; Delker, E.; Blanco, E.; Burrows, R.; Lozoff, B. Randomized Controlled Trial of Iron-Fortified versus Low-Iron Infant Formula: Developmental Outcomes at 16 Years. J. Pediatr. 2019, 212, 124–130. [Google Scholar] [CrossRef]
- Lozoff, B.; De Andraca, I.; Castillo, M.; Smith, J.B.; Walter, T.; Pino, P. Behavioral and Developmental Effects of Preventing Iron-Deficiency Anemia in Healthy Full-Term Infants. Pediatrics 2003, 112, 846–854. [Google Scholar] [CrossRef]
- Walter, T.; Pino, P.; Pizarro, F.; Lozoff, B. Prevention of Iron-Deficiency Anemia: Comparison of High- and Low-Iron Formulas in Term Healthy Infants after Six Months of Life. J. Pediatr. 1998, 132, 635–640. [Google Scholar] [CrossRef]
- Lozoff, B.; Castillo, M.; Clark, K.M.; Smith, J.B.; Sturza, J. Iron Supplementation in Infancy Contributes to More Adaptive Behavior at 10 Years of Age. J. Nutr. 2014, 144, 838–845. [Google Scholar] [CrossRef]
- Friel, J.K.; Aziz, K.; Andrews, W.L.; Harding, S.V.; Courage, M.L.; Adams, R.J. A Double-Masked, Randomized Control Trial of Iron Supplementation in Early Infancy in Healthy Term Breast-Fed Infants. J. Pediatr. 2003, 143, 582–586. [Google Scholar] [CrossRef]
- Iglesias Vázquez, L.; Canals, J.; Voltas, N.; Jardí, C.; Hernández, C.; Bedmar, C.; Escribano, J.; Aranda, N.; Jiménez, R.; Barroso, J.M.; et al. Does the Fortified Milk with High Iron Dose Improve the Neurodevelopment of Healthy Infants? Randomized Controlled Trial. BMC Pediatr. 2019, 19, 315. [Google Scholar] [CrossRef] [Green Version]
- McCann, S.; Perapoch Amadó, M.; Moore, S.E. The Role of Iron in Brain Development: A Systematic Review. Nutrients 2020, 12, 2001. [Google Scholar] [CrossRef]
- Todorich, B.; Pasquini, J.M.; Garcia, C.I.; Paez, P.M.; Connor, J.R. Oligodendrocytes and Myelination: The Role of Iron. Glia 2009, 57, 467–478. [Google Scholar] [CrossRef]
- Ji, P.; B Nonnecke, E.; Doan, N.; Lönnerdal, B.; Tan, B. Excess Iron Enhances Purine Catabolism Through Activation of Xanthine Oxidase and Impairs Myelination in the Hippocampus of Nursing Piglets. J. Nutr. 2019, 149, 1911–1919. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Huang, G.; Su, T.; Gao, H.; Attieh, Z.K.; McKie, A.T.; Anderson, G.J.; Vulpe, C.D. Decreased Hephaestin Activity in the Intestine of Copper-Deficient Mice Causes Systemic Iron Deficiency. J. Nutr. 2006, 136, 1236–1241. [Google Scholar] [CrossRef] [Green Version]
- Sreedhar, B.; Nair, K.M. Modulation of Aconitase, Metallothionein, and Oxidative Stress in Zinc-Deficient Rat Intestine during Zinc and Iron Repletion. Free Radic. Biol. Med. 2005, 39, 999–1008. [Google Scholar] [CrossRef]
- El Hendy, H. Effect of Dietary Zinc Deficiency on Hematological and Biochemical Parameters and Concentrations of Zinc, Copper, and Iron in Growing Rats. Toxicology 2001, 167, 163–170. [Google Scholar] [CrossRef]
- Kondaiah, P.; Yaduvanshi, P.S.; Sharp, P.A.; Pullakhandam, R. Iron and Zinc Homeostasis and Interactions: Does Enteric Zinc Excretion Cross-Talk with Intestinal Iron Absorption? Nutrients 2019, 11, 1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, J.-H.; Doguer, C.; Collins, J.F. Consumption of a High-Iron Diet Disrupts Homeostatic Regulation of Intestinal Copper Absorption in Adolescent Mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 2017, 313, G353–G360. [Google Scholar] [CrossRef] [PubMed]
- Esamai, F.; Liechty, E.; Ikemeri, J.; Westcott, J.; Kemp, J.; Culbertson, D.; Miller, L.V.; Hambidge, K.M.; Krebs, N.F. Zinc Absorption from Micronutrient Powder Is Low but Is Not Affected by Iron in Kenyan Infants. Nutrients 2014, 6, 5636–5651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Barker, S.; Knutson, M.D. Iron and Manganese Transport in Mammalian Systems. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2021, 1868, 118890. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Cousins, R.J. The Multiple Faces of the Metal Transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Y.; Jenkitkasemwong, S.; Duarte, S.; Sparkman, B.K.; Shawki, A.; Mackenzie, B.; Knutson, M.D. ZIP8 Is an Iron and Zinc Transporter Whose Cell-Surface Expression Is Up-Regulated by Cellular Iron Loading. J. Biol. Chem. 2012, 287, 34032–34043. [Google Scholar] [CrossRef] [Green Version]
- Tolkien, Z.; Stecher, L.; Mander, A.P.; Pereira, D.I.A.; Powell, J.J. Ferrous Sulfate Supplementation Causes Significant Gastrointestinal Side-Effects in Adults: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0117383. [Google Scholar] [CrossRef] [Green Version]
- Cancelo-Hidalgo, M.J.; Castelo-Branco, C.; Palacios, S.; Haya-Palazuelos, J.; Ciria-Recasens, M.; Manasanch, J.; Pérez-Edo, L. Tolerability of Different Oral Iron Supplements: A Systematic Review. Curr. Med. Res. Opin. 2013, 29, 291–303. [Google Scholar] [CrossRef]
- Lind, T.; Lönnerdal, B.; Stenlund, H.; Gamayanti, I.L.; Ismail, D.; Seswandhana, R.; Persson, L.-A. A Community-Based Randomized Controlled Trial of Iron and Zinc Supplementation in Indonesian Infants: Effects on Growth and Development. Am. J. Clin. Nutr. 2004, 80, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, J.; Sachdev, H.P.S.; Singh, T.; Mallika, V. A Randomized Placebo-Controlled Trial of Iron Supplementation in Breastfed Young Infants Initiated on Complementary Feeding: Effect on Haematological Status. J. Health Popul. Nutr. 2004, 22, 203–211. [Google Scholar]
- Bora, R.; Ramasamy, S.; Brown, B.; Wolfson, J.; Rao, R. Effect of Iron Supplementation from Neonatal Period on the Iron Status of 6-Month-Old Infants at-Risk for Early Iron Deficiency: A Randomized Interventional Trial. J. Matern. Fetal Neonatal Med. 2019, 34, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Ermis, B.; Demirel, F.; Demircan, N.; Gurel, A. Effects of Three Different Iron Supplementations in Term Healthy Infants after 5 Months of Life. J. Trop. Pediatr. 2002, 48, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smuts, C.M.; Dhansay, M.A.; Faber, M.; van Stuijvenberg, M.E.; Swanevelder, S.; Gross, R.; Benadé, A.J.S. Efficacy of Multiple Micronutrient Supplementation for Improving Anemia, Micronutrient Status, and Growth in South African Infants. J. Nutr. 2005, 135, 653S–659S. [Google Scholar] [CrossRef]
- Silva, D.G.; Franceschini, S.d.C.C.; Sigulem, D.M. Growth in Non-Anemic Infants Supplemented with Different Prophylactic Iron Doses. J. Pediatr. 2008, 84, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonyté Sjödin, K.; Domellöf, M.; Lagerqvist, C.; Hernell, O.; Lönnerdal, B.; Szymlek-Gay, E.A.; Sjödin, A.; West, C.E.; Lind, T. Administration of Ferrous Sulfate Drops Has Significant Effects on the Gut Microbiota of Iron-Sufficient Infants: A Randomised Controlled Study. Gut 2019, 68, 2095–2097. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.; Pfeiffer, C.M.; Georgieff, M.K.; Brittenham, G.; Fairweather-Tait, S.; Hurrell, R.F.; McArdle, H.J.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Iron Review. J. Nutr. 2018, 148, 1001S–1067S. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Frank, D.N.; Hendricks, A.E.; Ir, D.; Esamai, F.; Liechty, E.; Hambidge, K.M.; Krebs, N.F. Iron in Micronutrient Powder Promotes an Unfavorable Gut Microbiota in Kenyan Infants. Nutrients 2017, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Paganini, D.; Uyoga, M.A.; Kortman, G.A.M.; Cercamondi, C.I.; Moretti, D.; Barth-Jaeggi, T.; Schwab, C.; Boekhorst, J.; Timmerman, H.M.; Lacroix, C.; et al. Prebiotic Galacto-Oligosaccharides Mitigate the Adverse Effects of Iron Fortification on the Gut Microbiome: A Randomised Controlled Study in Kenyan Infants. Gut 2017, 66, 1956–1967. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.M.; Hill, D.R.; Aurora, M.; Spence, J.R. Morphogenesis and Maturation of the Embryonic and Postnatal Intestine. Semin. Cell Dev. Biol. 2017, 66, 81–93. [Google Scholar] [CrossRef]
- Black, R.E.; Heidkamp, R. Causes of Stunting and Preventive Dietary Interventions in Pregnancy and Early Childhood. In Nestlé Nutrition Institute Workshop Series; Colombo, J., Koletzko, B., Lampl, M., Eds.; S. Karger AG: Basel, Switzerland, 2018; Volume 89, pp. 105–113. ISBN 978-3-318-06351-6. [Google Scholar]
- Brown, K.H. Diarrhea and Malnutrition. J. Nutr. 2003, 133, 328S–332S. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the Microbiome in Human Development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of Microbial Consortia in the Developing Infant Gut Microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.E.; Townsend, S.D. Temporal Development of the Infant Gut Microbiome. Open Biol. 2019, 9, 190128. [Google Scholar] [CrossRef] [PubMed]
- Mueller, N.T.; Bakacs, E.; Combellick, J.; Grigoryan, Z.; Dominguez-Bello, M.G. The Infant Microbiome Development: Mom Matters. Trends Mol. Med. 2015, 21, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.; Nielsen, D.S.; Lauritzen, L.; Jakobsen, M.; Michaelsen, K.F. Impact of Diet on the Intestinal Microbiota in 10-Month-Old Infants. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 613–618. [Google Scholar] [CrossRef]
- O’Sullivan, A.; He, X.; McNiven, E.M.S.; Haggarty, N.W.; Lönnerdal, B.; Slupsky, C.M. Early Diet Impacts Infant Rhesus Gut Microbiome, Immunity, and Metabolism. J. Proteome Res. 2013, 12, 2833–2845. [Google Scholar] [CrossRef]
- Zivkovic, A.M.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Human Milk Glycobiome and Its Impact on the Infant Gastrointestinal Microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 4653–4658. [Google Scholar] [CrossRef] [Green Version]
- Bode, L. Human Milk Oligosaccharides: Every Baby Needs a Sugar Mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, A.R.; Barile, D.; Underwood, M.A.; Mills, D.A. The Impact of the Milk Glycobiome on the Neonate Gut Microbiota. Annu. Rev. Anim. Biosci. 2015, 3, 419–445. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria Can Protect from Enteropathogenic Infection through Production of Acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Xu, J.; Gordon, J.I. Honor Thy Symbionts. Proc. Natl. Acad. Sci. USA 2003, 100, 10452–10459. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Biomarker | Anemia Cutoff | Iron Deficiency Cutoff | Response to Iron Supplementation | References |
|---|---|---|---|---|
| Hemoglobin | <110 g/L | - | ↑ or no change | [27,28,29,30,31,32] |
| Serum Ferritin 1 | - | <12 µg/L | ↑ | [33,34,35] |
| Transferrin Saturation 1 | - | <10% | ↑ | [32,33] |
| Zinc Protoporphyrin | - | 80 μmol/mol heme | ↓ | [33] |
| Soluble Transferrin Receptor | - | 8.3 mg/L | ↓ | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMillen, S.A.; Dean, R.; Dihardja, E.; Ji, P.; Lönnerdal, B. Benefits and Risks of Early Life Iron Supplementation. Nutrients 2022, 14, 4380. https://doi.org/10.3390/nu14204380
McMillen SA, Dean R, Dihardja E, Ji P, Lönnerdal B. Benefits and Risks of Early Life Iron Supplementation. Nutrients. 2022; 14(20):4380. https://doi.org/10.3390/nu14204380
Chicago/Turabian StyleMcMillen, Shasta A., Richard Dean, Eileen Dihardja, Peng Ji, and Bo Lönnerdal. 2022. "Benefits and Risks of Early Life Iron Supplementation" Nutrients 14, no. 20: 4380. https://doi.org/10.3390/nu14204380