
Protection by -Biotics against Hypertension Programmed by Maternal High Fructose Diet: Rectification of Dysregulated Expression of Short-Chain Fatty Acid Receptors in the Hypothalamic Paraventricular Nucleus of Adult Offspring
 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maternal High Fructose Diet Model
2.2. Experimental Design
2.3. Biotic Preparation
2.4. Blood Pressure Measurements and Spectral Analysis of Systolic Blood Pressure Signals
2.5. Measurement of Plasma Norepinephrine
2.6. Tissue Collection from the Hypothalamus and Sample Preparation
2.7. Measurement of Malondialdehyde Level in the PVN
2.8. Measurement of Proinflammatory Cytokines in the PVN
2.9. Measurement of Reactive Oxygen Species in the PVN
2.10. Gas Chromatography-Mass Spectrometric (GC-MS) Analysis
2.11. Immunohistochemical Staining
2.12. Generation of Lentiviral Vector
2.13. Microinjection of Lentiviral Vectors into the PVN
2.14. Quantitative Real-Time Polymerase Chain Reaction
2.15. Statistical Analysis
3. Results
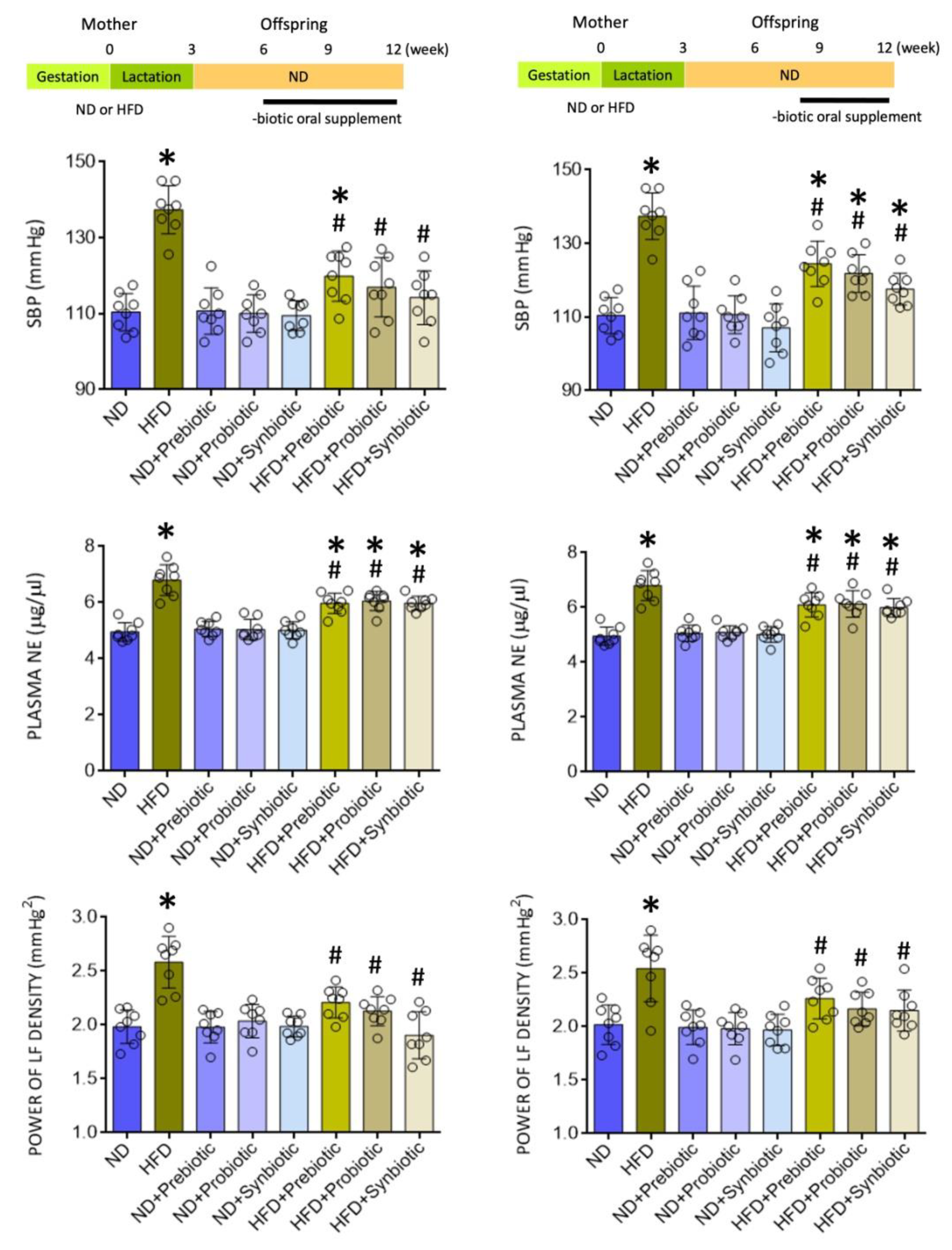
3.1. Oral Supplement with Prebiotic, Probiotic, or Synbiotic Protects against Programmed Hypertension and Sympathetic Activation in Adult HFD Offspring
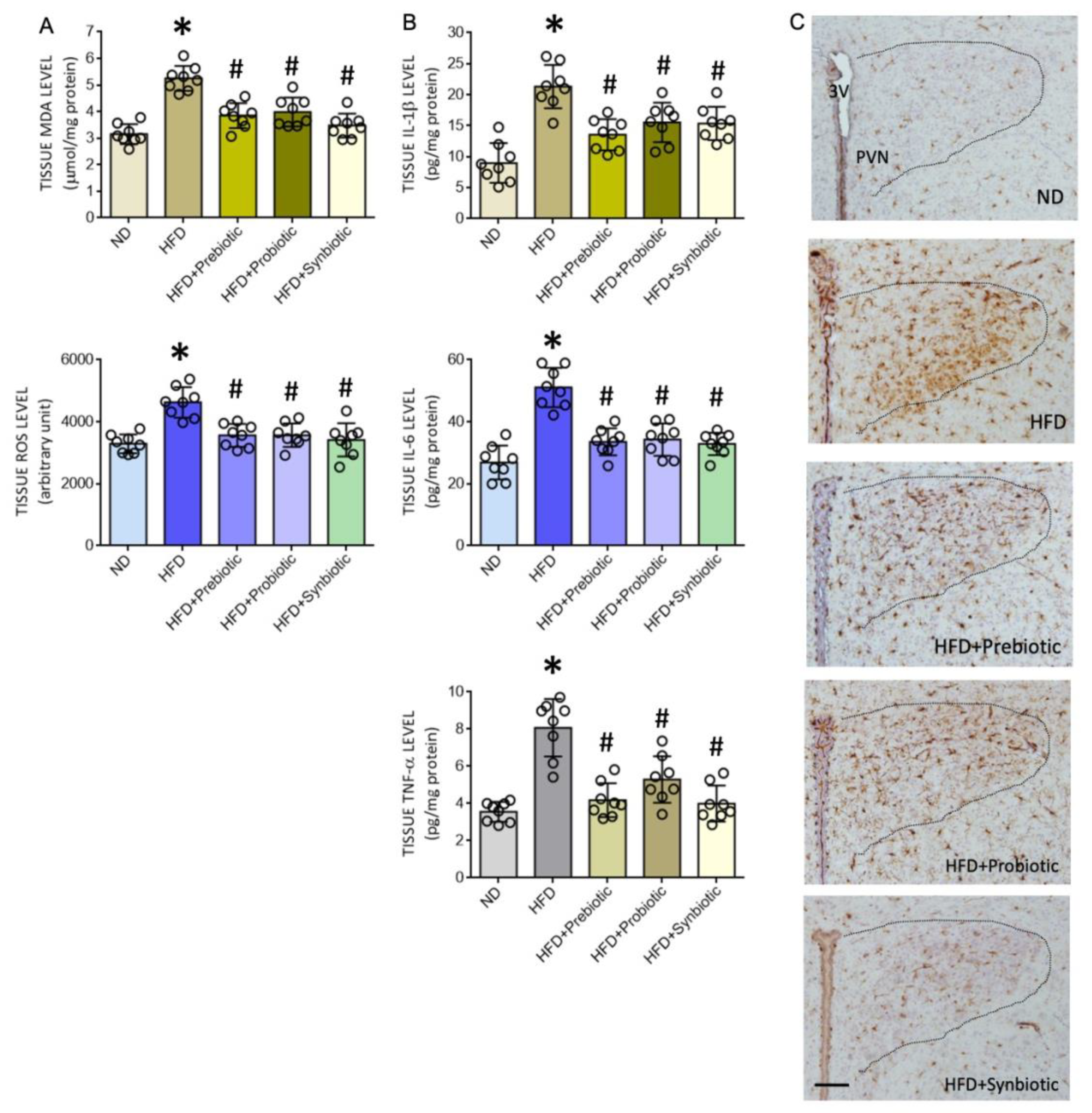
3.2. Oral Supplement with Prebiotic, Probiotic, or Synbiotic Alleviates Tissue Oxidative Stress and Inflammation in the PVN of HFD Offspring
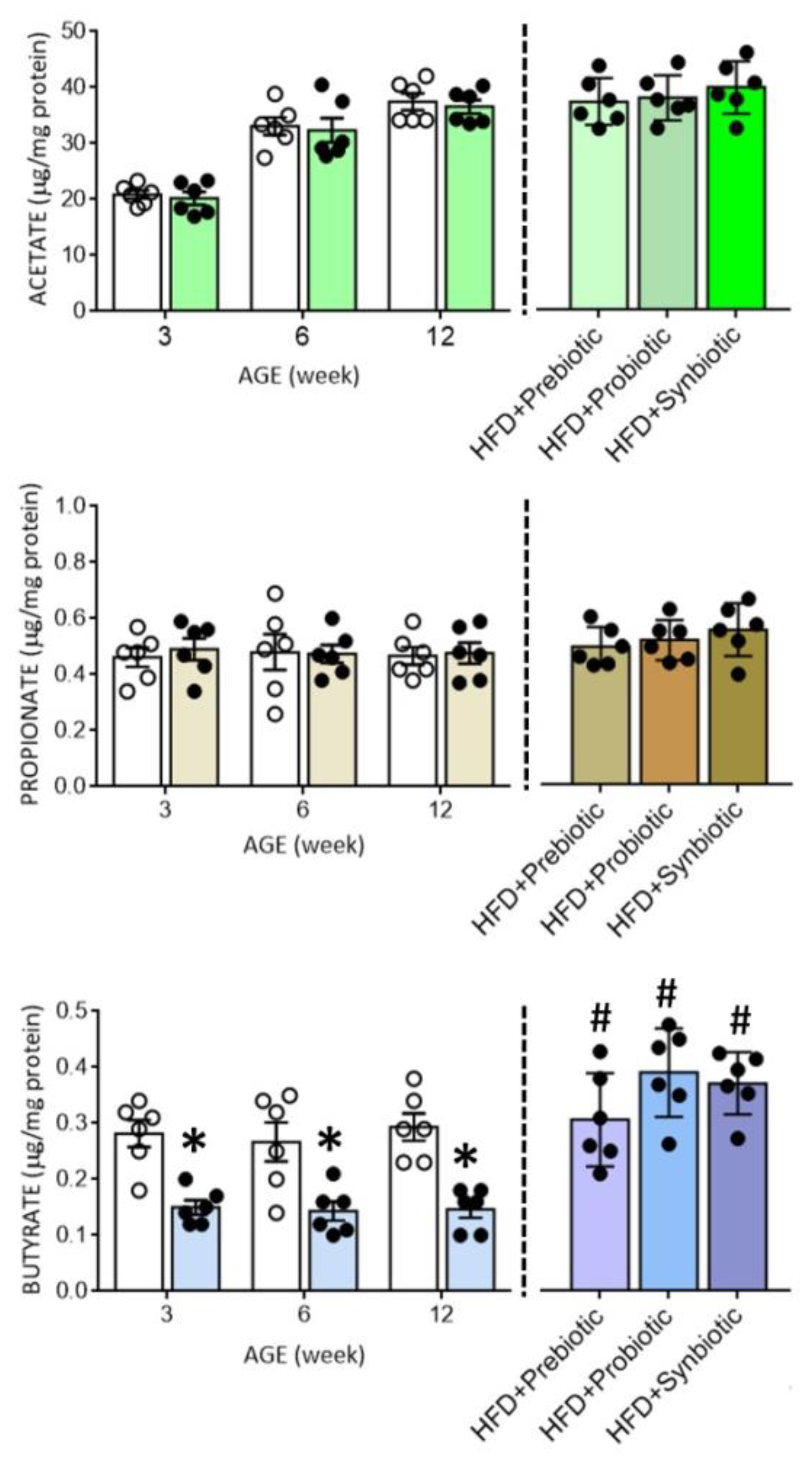
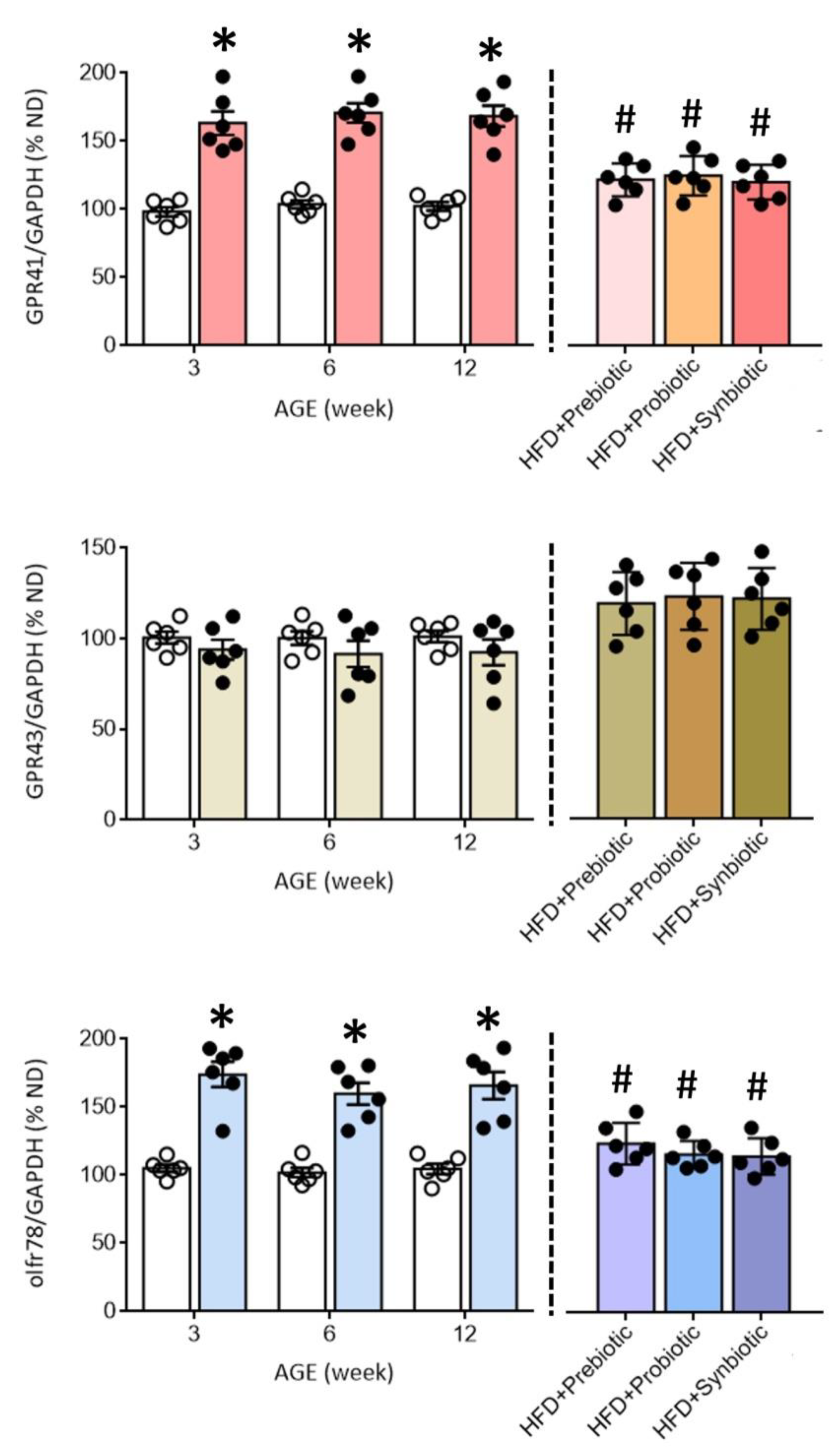
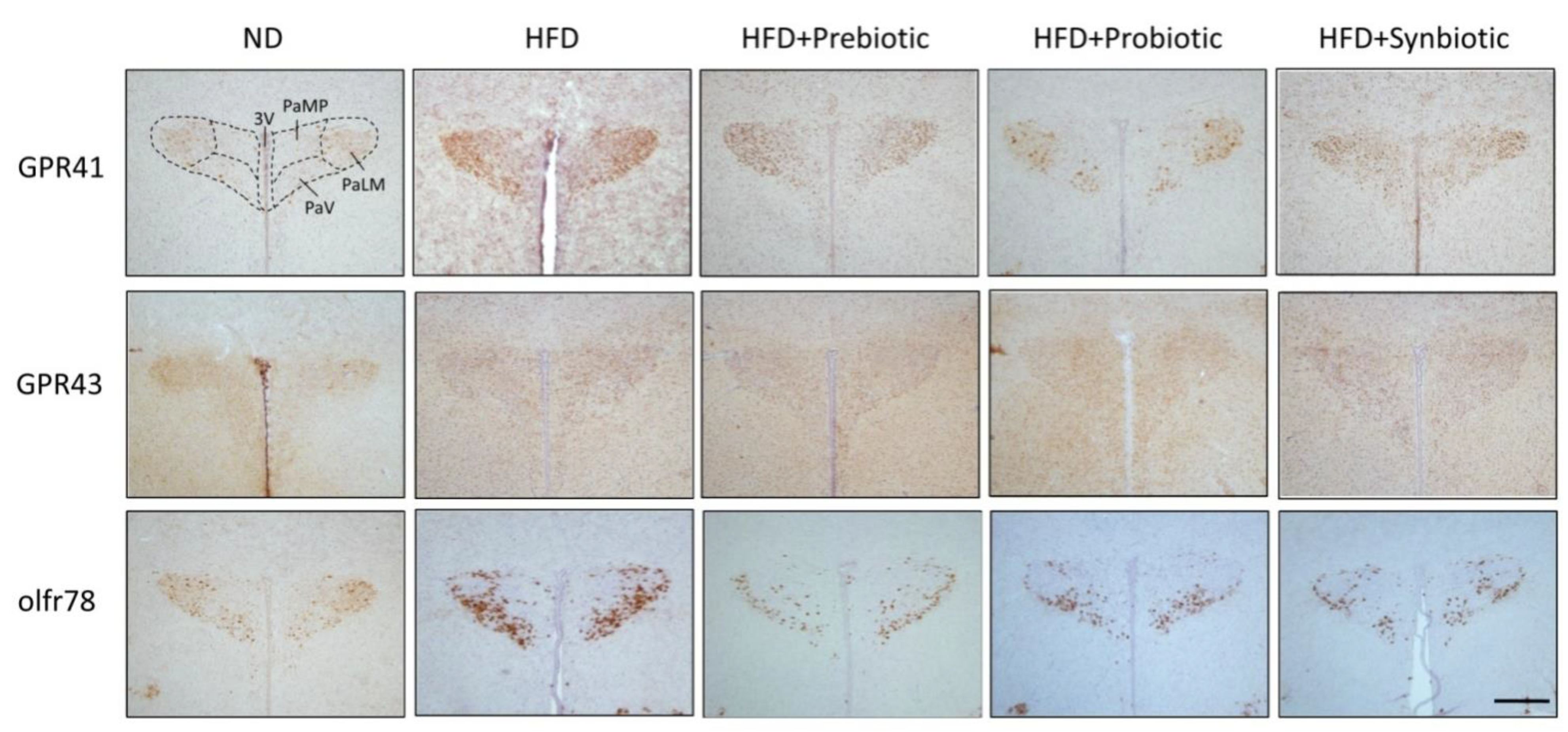
3.3. Maternal HFD Changes Tissue Butyrate Level and Expression of SCFA-Sensing Receptors in the PVN That Are Restored by Oral Supplement with Prebiotic, Probiotic or Synbiotic
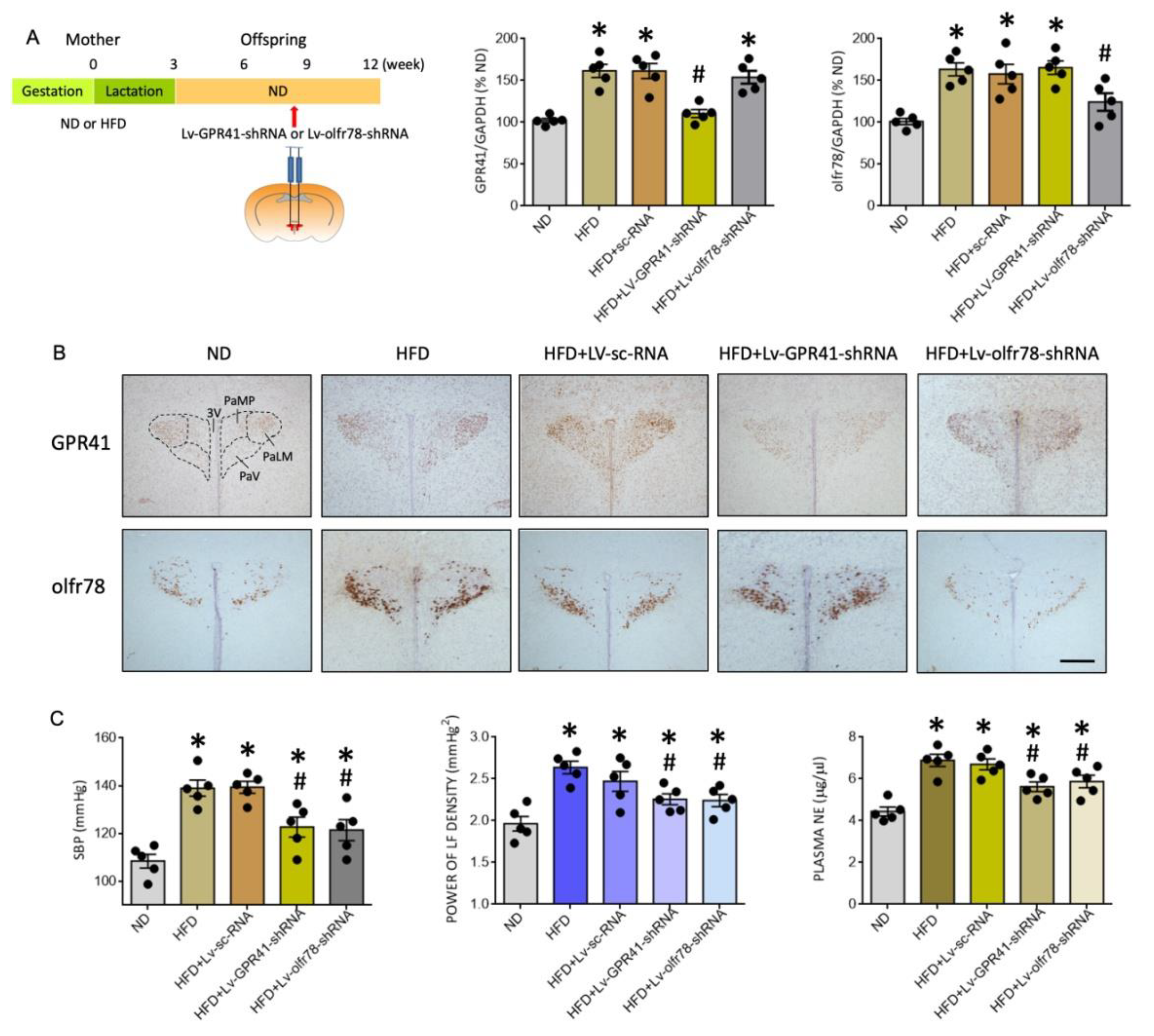
3.4. Gene Silencing of GPR41 and olfr78 in the PVN Alleviates Programmed Hypertension and Sympathetic Activation in HFD Offspring
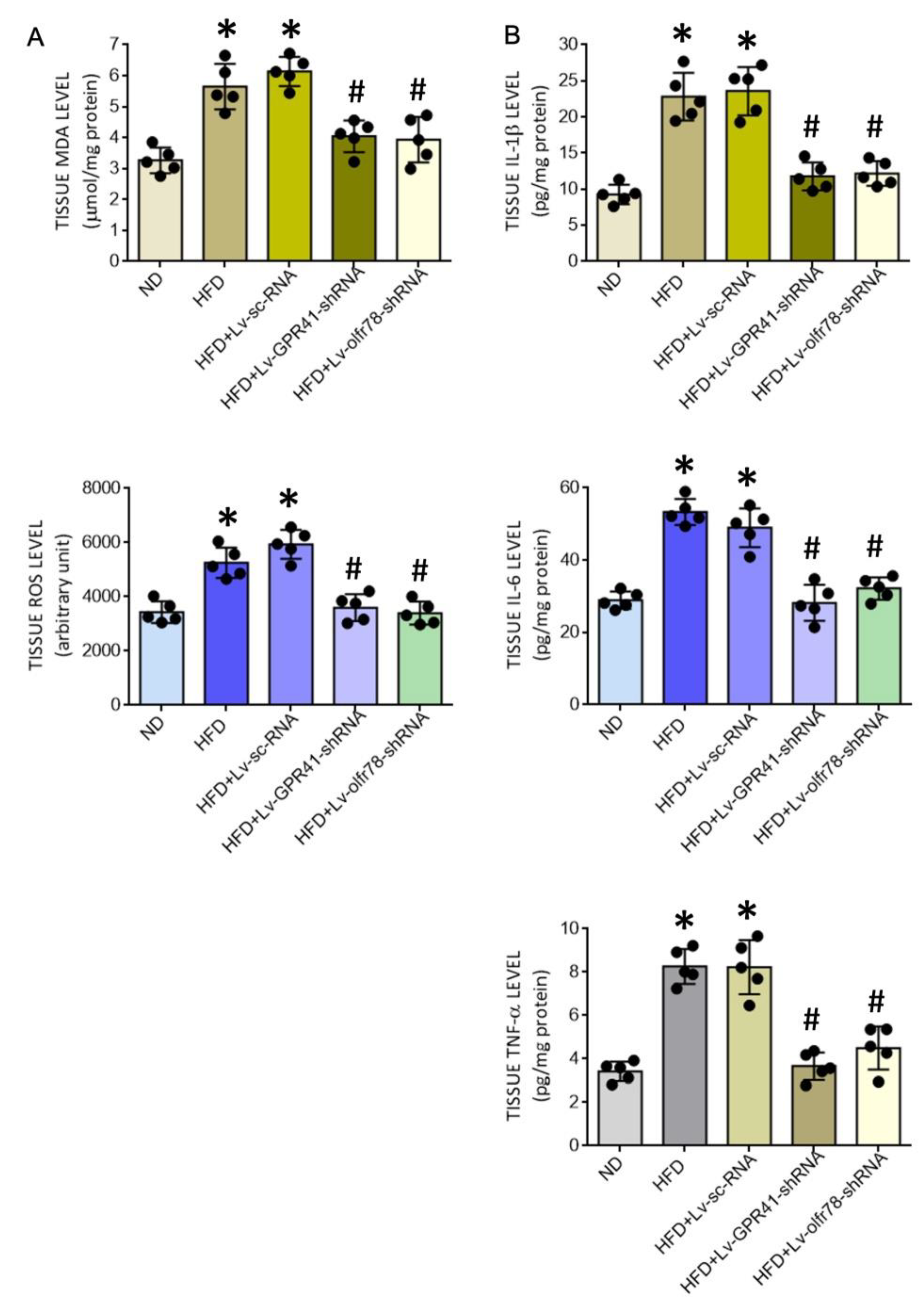
3.5. SCFA-Sensing Receptors Participate in Tissue Oxidative Stress and Inflammation in the PVN of HFD Offspring
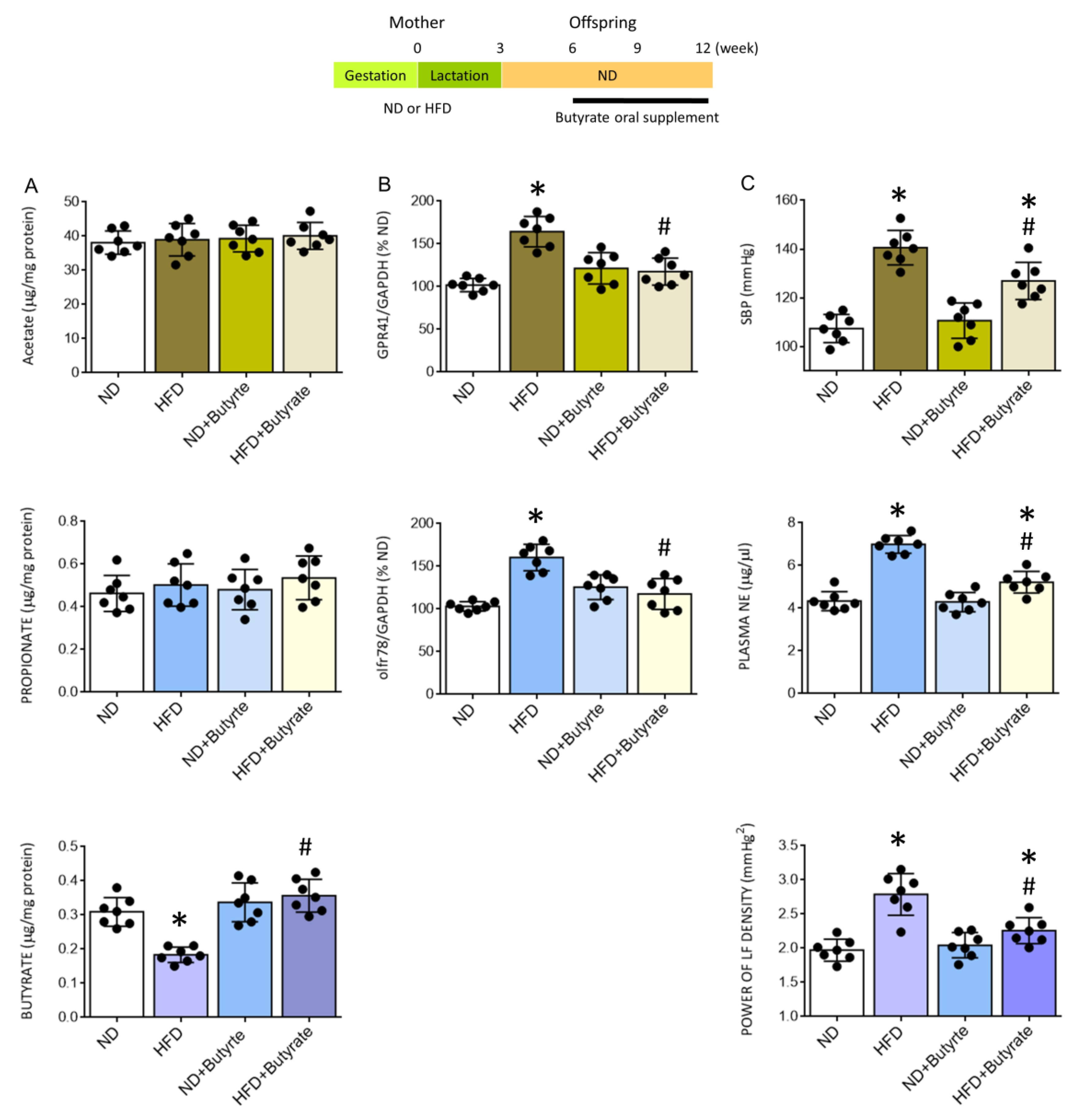
3.6. Oral Supplement with Butyrate Restores the Expression of SCFA-Sensing Receptors in the PVN and Confers Protection against Tissue Oxidative Stress, Neuroinflammation and Programmed Hypertension in HFD Offspring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bays, H.E.; Kulkarni, A.; German, C.; Satish, P.; Iluyomade, A.; Dudum, R.; Thakkar, A.; Rifai, M.I.; Mehta, A.; Thobani, A.; et al. Ten things to know about ten cardiovascular disease risk factors—2022. Am. J. Prev. Cardiol. 2022, 10, 100342. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Cooke, C.L.; Davidge, S.T. In utero origins of hypertension: Mechanisms and targets for therapy. Physiol. Rev. 2016, 96, 549–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasinger, J.H.; Davis, G.K.; Newsome, A.D.; Alexander, B.T. The developmental programming of hypertension: Physiological mechanisms. Hypertension 2016, 68, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Barker, D. Fetal origins of coronary heart disease. Br. Med. J. 1995, 311, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Ducsay, C.A.; Goyal, R.; Pearce, W.J.; Wilson, S.; Hu, X.Q.; Zhang, L. Gestational hypoxia and developmental plasticity. Physiol. Rev. 2018, 98, 1241–1334. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. The good, the bad, and the ugly of pregnancy nutrients and developmental programming of adult disease. Nutrients 2019, 11, 894. [Google Scholar] [CrossRef] [Green Version]
- Schulz, L.C. The Dutch Hunger Winter and the developmental origins of health and disease. Proc. Natl. Acad. Sci. USA 2010, 107, 16757–16758. [Google Scholar] [CrossRef] [Green Version]
- Mariano, V.S.; Boer, P.A.; Gontijo, J.A.R. Fetal undernutrition programming, sympathetic nerve activity, and arterial hypertension development. Front. Physiol. 2021, 12, 704819. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, F.H. How is maternal nutrition related to preterm birth? Annu. Rev. Nutr. 2011, 31, 235–261. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Fraser, A.; Fraser, W.D.; Hyppönen, E.; Davey Smith, G.; Deanfield, J.; Hingorani, A.; Sattar, N.; Lawlor, D.A. Associations of maternal 25-hydroxyvitamin D in pregnancy with offspring cardiovascular risk factors in childhood and adolescence: Findings from the Avon Longitudinal Study of parents and children. Heart 2013, 99, 1849–1856. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. The double-edged sword effects of maternal nutrition in the developmental programming of hypertension. Nutrients 2018, 10, 1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Prince, A.L.; Bader, D.; Hu, M.; Ganu, R.; Baquero, K.; Blundell, P.; Alan Harris, R.; Frias, A.E.; Grove, K.L.; et al. High-fat maternal diet during pregnancy persistently alters the offspring microbiome in a primate model. Nat. Commun. 2014, 5, 3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes. 2016, 7, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maher, S.E.; O’Brien, E.C.; Moore, R.L.; Byrne, D.F.; Geraghty, A.A.; Saldova, R.; Murphy, E.F.; Van Sinderen, D.; Cotter, P.D.; McAuliffe, F.M. The association between the maternal diet and the maternal and infant gut microbiome: A systematic review. Br. J. Nutr. 2020, 4, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Yu, H.R.; Chan, J.Y.H.; Wu, K.L.H.; Lee, W.C.; Tain, Y.L. The impact of gut microbiome on maternal fructose intake-induced developmental programming of adult disease. Nutrients 2022, 14, 1031. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, S.; Sun, J.; Buys, N.; Jayasinghe, R. Effect of probiotics on blood pressure: A systematic review and meta-analysis of randomized, controlled trials. Hypertension 2014, 64, 897–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaar, B.J.H.; Prodan, A.; Nieuwdorp, M.; Muller, M. Gut microbiota in hypertension and atherosclerosis: A review. Nutrients 2020, 12, 2982. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.F.; Li, J.D.; Huo, Y.L.; Zhang, Y.P.; Fang, Z.Q.; Wang, H.P.; Peng, W.; Johnson, A.K.; Xue, B. Blockade of angiotensin-converting enzyme or tumor necrosis factor-α reverses maternal high-fat diet-induced sensitization of angiotensin II hypertension in male rat offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R351–R359. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Maternal high-fat diet and offspring hypertension. Int. J. Mol. Sci. 2022, 23, 8179. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lee, W.C.; Wu, K.L.H.; Leu, S.; Chan, J.Y.H. Resveratrol prevents the development of hypertension programmed by maternal plus post-weaning high-fructose consumption through modulation of oxidative stress, nutrient-sensing signals, and gut microbiota. Mol. Nutr. Food Res. 2018, 62, e1800066. [Google Scholar] [CrossRef] [PubMed]
- Simões-Alves, A.C.; Arcoverde-Mello, A.P.F.C.; Campos, J.D.; Wanderley, A.G.; Leandro, C.V.G.; da Costa-Silva, J.H.; de Oliveira Nogueira Souza, V. Cardiometabolic effects of postnatal high-fat diet consumption in offspring exposed to maternal protein restriction in utero. Front. Physiol. 2022, 13, 829920. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zubcevic, J. Gut–brain axis in regulation of blood pressure. Front. Physiol. 2017, 8, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Raizada, M.K.; Richards, E.M. Gut-brain-bone marrow axis in hypertension. Curr. Opin. Nephrol. Hypertens. 2021, 30, 159–165. [Google Scholar] [CrossRef]
- Santisteban, M.M.; Qi, Y.; Zubcevic, J.; Kim, S.; Yang, T.; Shenoy, V.; Cole-Jeffrey, C.T.; Lobaton, G.O.; Stewart, D.C.; Rubiano, A.; et al. Hypertension-linked pathophysiological alterations in the gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef]
- Li, H.B.; Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. Maternal treatment with captopril persistently alters gut-brain communication and attenuates hypertension of male offspring. Hypertension 2020, 75, 1315–1324. [Google Scholar] [CrossRef]
- Sharma, R.K.; Yang, T.; Oliveira, A.C.; Lobaton, G.O.; Aquino, V.; Kim, S.; Richards, E.M.; Pepine, C.J.; Sumners, C.; Raizada, M.K.; et al. Microglial cells impact gut microbiota and gut pathology in angiotensin II-induced hypertension. Circ. Res. 2019, 124, 727–736. [Google Scholar] [CrossRef]
- Toral, M.; Robles-Vera, I.; de la Visitación, N.; Romero, M.; Yang, T.; Sánchez, M.; Gómez-Guzmán, M.; Jiménez, R.; Raizada, M.K.; Duarte, J. Critical role of the interaction gut microbiota—Sympathetic nervous system in the regulation of blood pressure. Front. Physiol. 2019, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Xia, W.J.; Xu, M.L.; Yu, X.J.; Du, M.M.; Li, X.H.; Yang, T.; Li, L.; Li, Y.; Kang, K.B.; Su, Q.; et al. Antihypertensive effects of exercise involve reshaping of gut microbiota and improvement of gut-brain axis in spontaneously hypertensive rat. Gut Microbes. 2021, 13, 1–24. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Zeimann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of prebiotic fiber and insufficient signaling through gut metabolite-sensing receptors leads to cardiovascular disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 10, 4410–4415. [Google Scholar] [CrossRef] [Green Version]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef] [PubMed]
- Muralitharan, R.; Marques, F.Z. Diet-related gut microbial metabolites and sensing in hypertension. J. Hum. Hypertens. 2021, 35, 162–169. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, Y.J.; Hou, C.Y.; Tain, Y.L. Maternal administration of probiotic or prebiotic prevents male adult rat offspring against developmental programming of hypertension induced by high fructose consumption in pregnancy and lactation. Nutrients 2018, 10, 1229. [Google Scholar] [CrossRef] [Green Version]
- Dampney, R.A.; Michelini, L.C.; Li, D.P.; Pan, H.L. Regulation of sympathetic vasomotor activity by the hypothalamic paraventricular nucleus in normotensive and hypertensive states. Am. J. Physiol. Heart. Circ Physiol. 2018, 315, H1200–H1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, Y.M.; Wu, K.L.H.; Tsai, P.C.; Tain, Y.L.; Leu, S.; Lee, W.C.; Chan, J.Y.H. Anomalous AMPK-regulated angiotensin AT1R expression and SIRT1-mediated mitochondrial biogenesis at RVLM in hypertension programming of offspring to maternal high fructose exposure. J. Biomed. Sci. 2020, 27, 68. [Google Scholar] [CrossRef]
- Reckelhoff, J.F. Gender differences in the regulation of blood pressure. Hypertension 2001, 37, 1199–1208. [Google Scholar] [CrossRef]
- Chunchai, T.; Thunapong, W.; Yasom, S.; Wanchai, K.; Eaimworawuthikul, S.; Metzler, G.; Lungkaphin, A.; Pongchaidecha, A.; Sirilun, S.; Chaiyasut, C.; et al. Decreased microglial activation through gut-brain axis by prebiotics, probiotics, or synbiotics effectively restored cognitive function in obese-insulin resistant rats. J. Neuroinflam. 2018, 15, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, M.K.; Tain, Y.L.; Chen, Y.W.; Hsu, W.H.; Yeh, Y.T.; Chang, S.K.C.; Liao, J.X.; Hou, C.Y. Resveratrol butyrate esters inhibit obesity caused by perinatal exposure to bisphenol A in female offspring rats. Molecules 2021, 26, 4010. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.H.H.; Wang, L.L.; Wang, S.H.; Chan, J.Y.H. Differential cardiovascular responses to blockade of nNOS or iNOS in rostral ventrolateral medulla of the rat. Br. J. Pharmacol. 2001, 133, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.C.; Chao, Y.M.; Chan, J.Y.H. Sympathetic activation of splenic T-lymphocytes in hypertension of adult offspring programmed by maternal high fructose exposure. Chin. J. Physiol. 2020, 63, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; American Press: New York, NY, USA, 2014; pp. 46–50. [Google Scholar]
- Chao, Y.M.; Rauchova, H.; Chan, J.Y.H. Disparate role of oxidative stress in rostral ventrolateral medulla in age-dependent susceptibility to hypertension induced by system L-NAME treatment in rats. Biomedicines 2022, 10, 2232. [Google Scholar] [CrossRef]
- Xiao, W.; Su, J.; Gao, X.; Yang, H.; Weng, R.; Gu, Y. The microbiota-gut-brain axis participates in chronic cerebral hypoperfusion by disrupting the metabolism of short-chain fatty acids. Microbiome 2022, 10, 62. [Google Scholar] [CrossRef]
- Griendling, K.K.; Camargo, L.L.; Rios, F.; Alves-Lopes, R.; Montezano, A.C.; Touyz, R.M. Oxidative stress and hypertension. Circ. Res. 2021, 128, 993–1020. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, Y.; Gua, C.; Wu, B. Elevated oxidative stress and inflammation in hypothalamic paraventricular nucleus are associated with sympathetic excitation and hypertension in rats exposed to chronic itermittent hypoxia. Front. Physiol. 2018, 9, 840. [Google Scholar] [CrossRef]
- Wei, S.G.; Yu, Y.; Felder, R.B. TNF-α-induced sympathetic excitation requires EGFR and ERK1/2 signaling in cardiovascular regulatory regions of the forebrain. Am. J. Physiol. Heart. Circ Physiol. 2021, 320, H772–H786. [Google Scholar] [CrossRef] [PubMed]
- Lymperopoulos, A.; Suster, M.S.; Borges, J.I. Short-chain fatty acid receptors and cardiovascular function. Int. J. Mol Sci. 2022, 23, 3303. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.A.; Ruggiero, D.A.; Joh, T.H.; Park, D.H.; Reis, D.J. Rostral ventrolateral medulla: Selective projections to the thoracic autonomic cell column from the region containing C1 adrenaline neurons. J. Comp. Neurol. 1984, 228, 168–185. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolognini, D.; Tobin, A.B.; Milligan, G.; Moss, C.E. The pharmacology and function of receptors for short-chain fatty acids. Mol. Pharmacol. 2016, 89, 388–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.H.H.; Hsu, K.S.; Huang, C.C.; Wang, L.L.; Ou, C.C.; Chan, J.Y.H. NADPH oxidase-derived superoxide anion mediates angiotensin II-induced pressor effect via activation of p38 mitogen-activated protein kinase in the rostral ventrolateral medulla. Cir. Res. 2005, 97, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.B.; Kang, Y.M.; Zhang, C.; Yu, X.J.; Chen, W.S. Infusion of melatonin into the paraventricular nucleus ameliorates myocardial ischemia–reperfusion injury by regulating oxidative stress and inflammatory cytokines. J. Cardiovasc. Pharmacol. 2019, 74, 336–347. [Google Scholar] [CrossRef]
- Souza, L.A.C.; Worker, C.J.; Li, W.; Trebak, F.; Watkins, T.; Gayban, A.J.B.; Yamasaki, E.; Cooper, S.G.; Drumm, B.T.; Feng, Y. (Pro)renin receptor knockdown in the paraventricular nucleus of the hypothalamus attenuates hypertension development and AT1 receptor-mediated calcium events. Am. J. Physiol. Heart Circ Physiol. 2019, 316, H1389–H1405. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Magee, K.L.; Colon-Perez, L.M.; Larkin, R.; Liao, Y.S.; Balazic, E.; Cowart, J.R.; Arocha, R.; Redler, T.; Febo, M.; et al. Impaired butyrate absorption in the proximal colon, low serum butyrate and diminished central effects of butyrate on blood pressure in spontaneously hypertensive rats. Acta Physiol. 2019, 226, e13256. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M.; SPRING Trial Group. Increased systolic and diastolic blood pressure is associated with altered gut microbiota composition and butyrate production in early pregnancy. Hypertension 2016, 68, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Zubcevic, J.; Richards, E.M.; Yang, T.; Kim, S.; Sumners, C.; Pepine, C.J.; Raizada, M.K. Impaired autonomic nervous system-microbiome circuit in hypertension: A premise for hypertension therapy. Circ. Res. 2019, 125, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L. Microbial short chain fatty acids and blood pressure regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Nakashima, N.; Nakashima, K.; Takano, M. Olfactory receptor 78 is expressed in hypothalamic vasopressin/oxytocin neurons, parenchymal microglia and choroidal macrophages in mice. Mol. Brain 2022, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hodaka, K.; Mohammed, M.; Handberg, E.M.; Richard, E.M.; Pepin, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Ling, Z.; Wang, F.; Chen, W.; Li, H.; Jin, J.; Zhang, H.; Pang, M.; Yu, J.; Liu, J. Clostridium butyricum pretreatment attenuates cerebral ischemia/reperfusion injury in mice via anti-oxidation and anti-apoptosis. Neurosci. Lett. 2016, 613, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Vijay, N.; Morris, M.E. Role of monocarboxylate transporters in drug delivery to the brain. Curr. Pharm. Des. 2014, 20, 1487–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robles-Vera, I.; Toral, M.; Duarte, J. Microbiota and hypertension: Role of the sympathetic nervous system and the immune system. Am. J. Hypertens. 2020, 33, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Yang, M.; Zhang, M.; Xiao, M.; Li, X. Butyrate mitigates TNF-α-induced attachment of monocytes to endothelial cells. J. Bioenerg. Biomembr. 2020, 52, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, C.; Zhu, J.; Lin, Q.; Yu, M.; Wen, J.; Feng, J.; Hu, C. Sodium butyrate ameliorates oxidative stress-induced intestinal epithelium barrier injury and mitochondrial damage through AMPK-mitophagy pathway. Oxid. Med. Cell Longev. 2022, 2022, 3745135. [Google Scholar] [CrossRef] [PubMed]
- Le Bourgot, C.; Ferret-Bernard, S.; Le Normand, L.; Savary, G.; Menendez-Aparicio, E.; Blat, S.; Appert-Bossard, E.; Respondek, F.; Le Huërou-Luron, I. Maternal short-chain fructooligosaccharide supplementation influences intestinal immune system maturation in piglets. PLoS ONE 2014, 9, e107508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vemuri, R.; Shinde, T.; Gundamaraju, R.; Gondalia, S.V.; Karpe, A.V.; Beale, D.J.; Martoni, C.J.; Eri, R. Lactobacillus acidophilus DDS-1, a probiotic strain, has shown promising probiotic character in vitro, as well as in clinical studies. Nutrients 2018, 10, 1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowiak-Kope, P.; Slizewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [Green Version]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Gómez-Guzmán, M.; Romero, M.; Yang, T.; Izquierdo-Garcia, J.; Jiménez, R.; Ruiz-Cabello, J.; et al. Probiotics prevents dysbiosis and rise in blood pressure in genetic hypertension: Role of short-chain fatty acids. Mol. Nutr. Food Res. 2020, 64, e1900616. [Google Scholar] [CrossRef] [PubMed]
- Bartolomaeus, H.; Avery, E.G.; Bartolomaeus, T.; Kozhakhmetov, S.; Zhumadilov, Z.; Müller, D.N.; Wilck, N.; Kushugulova, A.; Forslund, S.K. Blood pressure changes correlate with short-chain fatty acid production potential shifts under a synbiotic intervention. Cardiovasc. Res. 2020, 116, 1252–1253. [Google Scholar] [CrossRef] [PubMed]
- Le Blay, G.; Michel, C.; Blottière, H.M.; Cherbut, C. Prolonged intake of fructo-oligosaccharides induces a short-term elevation of lactic acid-producing bacteria and a persistent increase in cecal butyrate in rats. J. Nutr. 1999, 129, 2231–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Albiñana, P.; Martínez-González, A.; Camacho-Rodríguez, L.; Ferreira-Lazarte, A.; Villamiel, M.; Rodrigues-Díez, R.; Balfagón, G.; García-Redondo, A.B.; Prieto-Nieto, M.I.; Blanco-Rivero, J. Supplementation with the symbiotic formulation Prodefen® increases neuronal nitric oxide synthase and decreases oxidative stress in superior mesenteric artery from spontaneously hypertensive tats. Antioxidants. 2022, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guzmán, M.; Toral, M.; Romero, M.; Jiménez, R.; Galindo, P.; Sánchez, M.; Zarzuelo, M.J.; Olivares, M.; Gálvez, J.; Duarte, J. Antihypertensive effects of probiotics Lactobacillus strains in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2015, 59, 2326–2336. [Google Scholar] [CrossRef]
- Hadi, A.; Pourmasoumi, M.; Kazemi, M.; Najafgholizadeh, A.; Marx, W. Efficacy of synbiotic interventions on blood pressure: A systematic review and meta-analysis of clinical trials. Crit. Rev. Food Sci. Nutr. 2021, 2021, 1888278. [Google Scholar] [CrossRef]
- Arabi, S.M.; Bahrami, L.S.; Rahnama, I.; Sahebkar, A. Impact of synbiotic supplementation on cardiometabolic and anthropometric indices in patients with metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2022, 176, 106061. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Fellows, R.; Denizot, J.; Stellato, C.; Cuomo, A.; Jain, P.; Stoyanova, E.; Balazsi, S.; Hajnady, Z.; Liebert, A.; Kazakevych, J.; et al. Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases. Nat. Commun. 2018, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Aisenberg, W.H.; Huang, J.; Zhu, W.; Rajkumar, P.; Cruz, R.; Santhanam, L.; Natarajan, N.; Yong, H.M.; De Santiago, B.; Oh, J.J.; et al. Defining an olfactory receptor function in airway smooth muscle cells. Sci. Rep. 2016, 6, 38231. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND | HFD | |||
|---|---|---|---|---|
| Water Only | +Butyrate | Water Only | +Butyrate | |
| MDA (μmol/mg protein) | 3.07 ± 0.32 | 3.38 ± 0.55 | 5.20 ± 0.52 * | 3.88 ± 0.53 *,# |
| ROS (arbitrary unit) | 3263 ± 250 | 3052 ± 642 | 4535 ± 515 * | 3502 ± 413 # |
| IL-1β (pg/mg protein) | 7.54 ± 1.83 | 7.95 ± 2.99 | 19.80 ± 2.44 * | 13.16 ± 2.77 *,# |
| IL-6 (pg/mg protein) | 26.38 ± 6.01 | 24.42 ± 4.60 | 51.13 ± 6.89 * | 33.29 ± 6.57 *,# |
| TNF-α (pg/mg protein) | 3.58 ± 0.54 | 3.19 ± 0.95 | 8.46 ± 1.55 * | 4.39 ± 0.74 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chao, Y.-M.; Tain, Y.-L.; Lee, W.-C.; Wu, K.L.H.; Yu, H.-R.; Chan, J.Y.H. Protection by -Biotics against Hypertension Programmed by Maternal High Fructose Diet: Rectification of Dysregulated Expression of Short-Chain Fatty Acid Receptors in the Hypothalamic Paraventricular Nucleus of Adult Offspring. Nutrients 2022, 14, 4306. https://doi.org/10.3390/nu14204306
Chao Y-M, Tain Y-L, Lee W-C, Wu KLH, Yu H-R, Chan JYH. Protection by -Biotics against Hypertension Programmed by Maternal High Fructose Diet: Rectification of Dysregulated Expression of Short-Chain Fatty Acid Receptors in the Hypothalamic Paraventricular Nucleus of Adult Offspring. Nutrients. 2022; 14(20):4306. https://doi.org/10.3390/nu14204306
Chicago/Turabian StyleChao, Yung-Mei, You-Lin Tain, Wei-Chia Lee, Kay L. H. Wu, Hong-Ren Yu, and Julie Y. H. Chan. 2022. "Protection by -Biotics against Hypertension Programmed by Maternal High Fructose Diet: Rectification of Dysregulated Expression of Short-Chain Fatty Acid Receptors in the Hypothalamic Paraventricular Nucleus of Adult Offspring" Nutrients 14, no. 20: 4306. https://doi.org/10.3390/nu14204306