
Centenarian-Sourced Lactobacillus casei Combined with Dietary Fiber Complex Ameliorates Brain and Gut Function in Aged Mice


Abstract
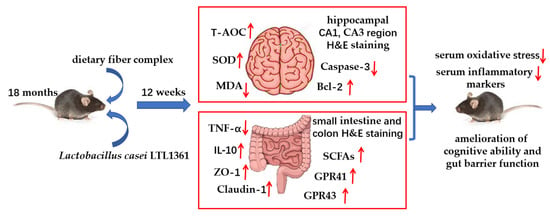
:
1. Introduction
2. Materials and Methods
2.1. Bacteria Strain and Culture
2.2. Preparation of DFC
2.3. Animals and Experimental Design
2.4. Morris Water Maze Test
2.5. Preparation of Tissues and Blood
2.6. Histopathological Analysis
2.7. Measurement of Oxidation-Associated Biomarkers and Inflammatory Cytokines
2.8. SCFA Analysis
2.9. RNA Isolation and Quantitative Real-Time PCR analysis
2.10. Statistical Analysis
3. Results
3.1. Effects of Lactobacillus casei LTL1361 and DFC on Learning and Memory of Aged Mice
3.2. Effects of Lactobacillus casei LTL1361 and DFC on Gene Expression of Apoptosis-Related Protein Markers
3.3. Effects of Lactobacillus casei LTL1361 and DFC on Hippocampus Histology in Aged Mice
3.4. Effects of Lactobacillus casei LTL1361 and DFC on Oxidative Stress
3.5. Effects of Lactobacillus casei LTL1361 and DFC on Inflammatory Markers
3.6. Effects of Lactobacillus casei LTL1361 and DFC on Gut Barrier Function in Aged Mice
3.7. Effects of Lactobacillus casei LTL1361 and DFC on SCFA Concentration and mRNA Expression of SCFA Receptors in Aged Mice
3.8. Correlation among SCFAs, Oxidative Stress, and Inflammatory Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Beard, J.R.; Officer, A.; de Carvalho, I.A.; Sadana, R.; Pot, A.M.; Michel, J.-P.; Lloyd-Sherlock, P.; Epping-Jordan, J.E.; Peeters, G.M.E.E.G.; Mahanani, W.R.; et al. The World report on ageing and health: A policy framework for healthy ageing. Lancet 2016, 387, 2145–2154. [Google Scholar] [CrossRef] [Green Version]
- Barzilai, N.; Huffman, D.M.; Muzumdar, R.H.; Bartke, A. The Critical Role of Metabolic Pathways in Aging. Diabetes 2012, 61, 1315–1322. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Dillin, A.; Gottschling, D.E.; Nyström, T. The good and the bad of being connected: The integrons of aging. Curr. Opin. Cell Biol. 2014, 26, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Harman, D. Aging: A Theory Based on Free Radical and Radiation Chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Rottenberg, H.; Hoek, J.B. The path from mitochondrial ROS to aging runs through the mitochondrial permeability transition pore. Aging Cell 2017, 16, 943–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raz, N.; Daugherty, A.M. Pathways to Brain Aging and Their Modifiers: Free-Radical-Induced Energetic and Neural Decline in Senescence (FRIENDS) Model—A Mini-Review. Gerontology 2018, 64, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; Naidoo, A.; Szamosi, J.; Verschoor, C.P.; Loukov, D.; Schenck, L.P.; Jury, J.; Foley, K.P.; et al. Age-Associated Microbial Dysbiosis Promotes Intestinal Permeability, Systemic Inflammation, and Macrophage Dysfunction. Cell Host Microbe 2017, 21, 455–466.e4. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Colangelo, L.; Reyfman, P.; Estepar, R.S.J.; Ash, S.; Thyagarajan, B.; Wells, J.; Putman, R.; Carnethon, M.; Washko, G.; et al. Systemic inflammation in young adults is associated with interstitial changes in middle age. Am. J. Respir. Crit. Care Med. 2021, 203, A1001. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, D.; Song, L.; Yu, L.; Song, Y.; Wang, D. Experimental study on effect of aging on intestinal mucosal barrier function in mice. Chin. J. Immunology 2018, 34, 1451–1455. [Google Scholar]
- Matt, S.M.; Lawson, M.A.; Johnson, R.W. Aging and peripheral lipopolysaccharide can modulate epigenetic regulators and decrease IL-1 beta promoter DNA methylation in microglia. Neurobiol. Aging. 2016, 47, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.-H.; Lai, C.-S.; Tsai, M.-L.; Wu, J.-C.; Ho, C.-T. Molecular mechanisms for anti-aging by natural dietary compounds. Mol. Nutr. Food Res. 2012, 56, 88–115. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.K.; Yi, Y.; Kim, J.K.; Kim, H.; Hossain, M.A.; Kim, J.; Cho, J.Y. Korean Red Ginseng plays an anti-aging role by mod-ulating expression of aging-related genes and immune cell subsets. Molecules 2020, 25, 1492. [Google Scholar] [CrossRef] [Green Version]
- Fontana, L.; Partridge, L. Promoting Health and Longevity through Diet: From Model Organisms to Humans. Cell 2015, 161, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.; Chase, A.B.; Weihe, C.; Orchanian, S.B.; Riedel, S.F.; Hendrickson, C.L.; Lay, M.; Sewall, J.M.; Martiny, J.B.H.; Whiteson, K. High-Fiber, Whole-Food Dietary Intervention Alters the Human Gut Microbiome but Not Fecal Short-Chain Fatty Acids. mSystems 2021, 6, 6. [Google Scholar] [CrossRef]
- Salamone, D.; Rivellese, A.A.; Vetrani, C. The relationship between gut microbiota, short-chain fatty acids and type 2 diabetes mellitus: The possible role of dietary fibre. Acta Diabetol. 2021, 58, 1131–1138. [Google Scholar] [CrossRef]
- Berding, K.; Carbia, C.; Cryan, J.F. Going with the grain: Fiber, cognition, and the microbiota-gut-brain-axis. Exp. Biol. Med. 2021, 246, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Hamaker, B.R.; Tuncil, Y.E. A Perspective on the Complexity of Dietary Fiber Structures and Their Potential Effect on the Gut Microbiota. J. Mol. Biol. 2014, 426, 3838–3850. [Google Scholar] [CrossRef]
- King, D.E.; Mainous, A.; Lambourne, C.A. Trends in Dietary Fiber Intake in the United States, 1999-J. Acad. Nutr. Diet. 2012, 112, 642–648. [Google Scholar] [CrossRef]
- Biagi, E.; Candela, M.; Franceschi, C.; Brigidi, P. The aging gut microbiota: New perspectives. Ageing Res. Rev. 2011, 10, 428–429. [Google Scholar] [CrossRef]
- Wang, F.; Yu, T.; Huang, G.; Cai, D.; Liang, X.; Su, H.; Zhu, Z.; Li, D.; Yang, Y.; Shen, P.; et al. Gut Microbiota Community and Its Assembly Associated with Age and Diet in Chinese Centenarians. J. Microbiol. Biotechnol. 2015, 25, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wang, F.; Sun, D.-W.; Li, Q. Intestinal Lactobacillus community structure and its correlation with diet of Southern Chinese elderly subjects. J. Microbiol. 2016, 54, 594–601. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO: Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic ACID bacteria. Available online: www.fao.org/3/a-a0512e.pdf (accessed on 1 March 2018).
- Yu, X.; Li, S.; Yang, D.; Qiu, L.; Wu, Y.; Wang, D.; Shah, N.P.; Xu, F.; Wei, H. A novel strain of Lactobacillus mucosae isolated from a Gaotian villager improves in vitro and in vivo antioxidant as well as biological properties in d-galactose-induced aging mice. J. Dairy Sci. 2016, 99, 903–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Tian, F.; Zhao, N.; Zhai, Q.; Zhang, H.; Chen, W. Effects of probiotics on d -galactose-induced oxidative stress in plasma: A meta-analysis of animal models. J. Funct. Foods 2017, 39, 44–49. [Google Scholar] [CrossRef]
- Ho, S.-T.; Hsieh, Y.-T.; Wang, S.-Y.; Chen, M.-J. Improving effect of a probiotic mixture on memory and learning abilities in d-galactose–treated aging mice. J. Dairy Sci. 2019, 102, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-W.; Tsai, Y.-S.; Chen, Y.-L.; Wang, M.-F.; Chen, C.-C.; Lin, W.-H.; Fang, T.J. Lactobacillus plantarum GKM3 Promotes Longevity, Memory Retention, and Reduces Brain Oxidation Stress in SAMP8 Mice. Nutrients 2021, 13, 2860. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yang, Z.; Chen, X.; Huang, Y.; Yin, B.; Guo, F.; Zhao, H.; Huang, J.; Wu, Y.; Gu, R. Effect of Lactobacillus rhamnosus hsryfm 1301 on the Gut Microbiota and Lipid Metabolism in Rats Fed a High-Fat Diet. J. Microbiol. Biotechnol. 2015, 25, 687–695. [Google Scholar] [CrossRef]
- Jiang, J.; Feng, N.; Zhang, C.; Liu, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Lactobacillus reuteri A9 and Lactobacillus mucosae A13 isolated from Chinese superlongevity people modulate lipid metabolism in a hypercholesterolemia rat model. FEMS Microbiol. Lett. 2019, 366, 366. [Google Scholar] [CrossRef]
- Yang, H.-Y.; Liu, S.-L.; Ibrahim, S.A.; Zhao, L.; Jiang, J.-L.; Sun, W.-F.; Ren, F.-Z. Oral administration of live Bifidobacterium substrains isolated from healthy centenarians enhanced immune function in BALB/c mice. Nutr. Res. 2009, 29, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Zhao, S.; Li, D.; Chang, F.; Tian, X.; Huang, G.; Zhu, Z.; Liu, D.; Dou, X.; Li, S.; et al. Nutrient Intake Is Associated with Longevity Characterization by Metabolites and Element Profiles of Healthy Centenarians. Nutrients 2016, 8, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.Y.; Mei, L.H. Lactobacillus casei Strain and Its Application. CN110144304A, 20 August 2019. [Google Scholar]
- Fu, C.-X.; Dai, L.; Yuan, X.-Y.; Xu, Y.-J. Effects of Fish Oil Combined with Selenium and Zinc on Learning and Memory Impairment in Aging Mice and Amyloid Precursor Protein Processing. Biol. Trace Elem. Res. 2021, 199, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wang, L.; Wen, C.; Yang, J.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber and its depolymerized derivative prevent obesity in association with modification of gGut microbiota in high-fat diet-fed mice. Mol. Nutr. Food Res. 2018, 62, 1800446. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wu, S.; Wang, L.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber modulates the gut microbiota and its metabolites in normal mice. Int. J. Biol. Macromol. 2018, 120, 502–512. [Google Scholar] [CrossRef]
- Barnhart, C.D.; Yang, D.; Lein, P.J. Using the Morris Water Maze to Assess Spatial Learning and Memory in Weanling Mice. PLoS ONE 2015, 10, e0124521. [Google Scholar] [CrossRef]
- Ni, Y.; Yang, X.; Zheng, L.; Wang, Z.; Wu, L.; Jiang, J.; Yang, T.; Ma, L.; Fu, Z. Lactobacillus and Bifidobacterium Improves Physiological Function and Cognitive Ability in Aged Mice by the Regulation of Gut Microbiota. Mol. Nutr. Food Res. 2019, 63, e1900603. [Google Scholar] [CrossRef]
- Fuchs, Y.; Steller, H. Programmed cell death in animal development and disease. Cell 2011, 147, 1640. [Google Scholar] [CrossRef] [Green Version]
- Benowitz, L.I.; He, Z.; Goldberg, J.L. Reaching the brain: Advances in optic nerve regeneration. Exp. Neurol. 2017, 287, 365–373. [Google Scholar] [CrossRef]
- Stackman, R.W., Jr.; Cohen, S.J.; Lora, J.C.; Rios, L.M. Temporary inactivation reveals that the CA1 region of the mouse dorsal hippocampus plays an equivalent role in the retrieval of long-term object memory and spatial memory. Neurobiol. Learn. Mem. 2016, 133, 118–128. [Google Scholar] [CrossRef]
- Tash, B.R.; Bewley, M.C.; Russo, M.; Keil, J.; Griffin, K.A.; Sundstrom, J.; Antonetti, D.; Tian, F.; Flanagan, J.M. The occludin and ZO-1 complex, defined by small angle X-ray scattering and NMR, has implications for modulating tight junction permeability. Proc. Natl. Acad. Sci. USA 2012, 109, 10855–10860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skamrahl, M.; Pang, H.; Ferle, M.; Gottwald, J.; Ruebeling, A.; Maraspini, R.; Honigmann, A.; Oswald, T.A.; Janshoff, A. tight junction ZO proteins maintain tissue fluidity, ensuring efficient collective cell migration. Adv. Sci. 2021, 8, 2170122. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-Chain Fatty Acids Activate GPR41 and GPR43 on Intestinal Epithelial Cells to Promote Inflammatory Responses in Mice. Gastroenterology 2013, 145, 396–406.e10. [Google Scholar] [CrossRef]
- Kravitz, E.; Schmeidler, J.; Beeri, M.S. Cognitive Decline and Dementia in the Oldest-Old. Rambam Maimonides Med. J. 2012, 3, e0026. [Google Scholar] [CrossRef] [Green Version]
- Sloan, K.J.; McRorie, J.W., Jr. Structure and function of dietary fiber: The physics of fiber in the gastrointestinal tract. Cereal Food World 2020, 65, 0028. [Google Scholar] [CrossRef]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Murga-Garrido, S.M.; Hong, Q.; Cross, T.-W.L.; Hutchison, E.R.; Han, J.; Thomas, S.P.; Vivas, E.I.; Denu, J.; Ceschin, D.G.; Tang, Z.-Z.; et al. Gut microbiome variation modulates the effects of dietary fiber on host metabolism. Microbiome 2021, 9, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Vizuete, A.A.; Robles, F.; Rodríguez-Rodríguez, E.; López-Sobaler, A.M.; Ortega, R.M. Association between food and nutrient intakes and cognitive capacity in a group of institutionalized elderly people. Eur. J. Nutr. 2010, 49, 293–300. [Google Scholar] [CrossRef]
- Khan, A.N.; Raine, L.B.; Drollette, E.S.; Scudder, M.R.; Kramer, A.; Hillman, C. Dietary Fiber Is Positively Associated with Cognitive Control among Prepubertal Children. J. Nutr. 2015, 145, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Hassevoort, K.M.; Lin, A.S.; Khan, N.A.; Hillman, C.H.; Cohen, N.J. Added sugar and dietary fiber consumption are associated with creativity in preadolescent children. Nutr. Neurosci. 2020, 23, 791–802. [Google Scholar] [CrossRef]
- Gong, R.; Wang, D.; Abbas, G.; Li, S.; Liu, Q.; Cui, M.; Zhang, X.-E. A switch-on molecular biosensor for detection of caspase-3 and imaging of apoptosis of cells. Sci. China Life Sci. 2021, 1–10. [Google Scholar] [CrossRef]
- Sabel, A.B.; Tawfik, M.; Zhang, X.; Grigartzik, L.; Heiduschka, P.; Hintz, W.; Henrich-Noack, P.; van Wachem, B. Gene therapy with caspase-3 small interfering RNA-nanoparticles is neuroprotective after optic nerve damage. Neural Regen. Res. 2021, 16, 2534–2541. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Huh, N.; Lee, J.; Yun, I.; Lee, J.W.; Lee, I.; Jung, M.W. Role of the hippocampal CA1 region in incremental value learning. Sci. Rep. 2018, 8, 9870. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Barja, G. The mitochondrial free radical theory of aging. In Book Progress in Molecular Biology and Translational Science; Osiewacz, H., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 127, pp. 1–27. [Google Scholar] [CrossRef]
- Ubando-Rivera, J.; Navarro-Ocana, A.; Valdivia-Lopez, M.A. Mexican lime peel: Comparative study on contents of dietary fibre and associated antioxidant activity. Food Chem. 2005, 89, 57–61. [Google Scholar] [CrossRef]
- Bhaskar, J.J.; Mahadevamma, S.; Chilkunda, N.D.; Salimath, P.V. Banana (Musa sp. var. elakki bale) Flower and Pseudostem: Dietary Fiber and Associated Antioxidant Capacity. J. Agric. Food Chem. 2012, 60, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary fiber and health outcomes: An umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sichetti, M.; De Marco, S.; Pagiotti, R.; Traina, G.; Pietrella, D. Anti-inflammatory effect of multistrain probiotic formulation (L. rhamnosus, B. lactis, and B. longum). Nutrition 2018, 53, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.P. Redefining Oxidative Stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Cl. Ob. 2011, 25, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.; Nichol, A. Inflammation, immunity and allergy. Anaesth. Intensive Care Med. 2021, 22, 488–493. [Google Scholar] [CrossRef]
- Sargi, S.C.; Dalalio, M.M.D.O.; Visentainer, J.V.; Bezerra, R.C.; Perini, J. Ângelo, D.L.; Stevanato, F.B.; Visentainer, J.E.L. Production of TNF-?, nitric oxide and hydrogen peroxide by macrophages from mice with paracoccidioidomycosis that were fed a linseed oil-enriched diet. Memórias Do Inst. Oswaldo Cruz 2012, 107, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Levast, B.; Li, Z.; Madrenas, J. The role of IL-10 in microbiome-associated immune modulation and disease tolerance. Cytokine 2015, 75, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Dupont, H.L. New Approaches for Bacteriotherapy: Prebiotics, New-Generation Probiotics, and Synbiotics. Clin. Infect. Dis. 2015, 60, S108–S121. [Google Scholar] [CrossRef]
- Milajerdi, A.; Mousavi, S.M.; Sadeghi, A.; Moghaddam, A.S.; Parohan, M.; Larijani, B.; Esmaillzadeh, A. The effect of probiotics on inflammatory biomarkers: A meta-analysis of randomized clinical trials. Eur. J. Nutr. 2020, 59, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, L.; Borrero, M.-J.; Ubeda, M.; Paule, L.; Lario, M.; Díaz, D.; Conde, E.; Rodríguez-Serrano, M.; Sánchez-Díaz, A.-M.; Del Campo, R.; et al. Intestinal inflammation worsens epithelial barrier function in rats with cirrhosis and ascites: Role of commensal gut flora. J. Hepatol. 2017, 66, S391. [Google Scholar] [CrossRef]
- Lambert, J.; Bitzer, Z.; Wopperer, A.; Elias, R. Dietary Soy Protein Concentrate Suppresses Colonic Inflammation and Loss of Gut Barrier Function In Vitro and in Mice. FASEB J. 2015, 29, 922.32. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of short-chain fatty acids and their receptors in inflammation and carcin-ogenesis. Pharmacol. Therapeut. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretta, M.D.; Quiroga, J.; López, R.; Hidalgo, M.A.; Burgos, R.A. Participation of Short-Chain Fatty Acids and Their Receptors in Gut Inflammation and Colon Cancer. Front. Physiol. 2021, 12, 662739. [Google Scholar] [CrossRef] [PubMed]
- Keenan, M.J.; Marco, M.L.; Ingram, D.K.; Martin, R.J. Improving healthspan via changes in gut microbiota and fermentation. AGE 2015, 37, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matt, S.M.; Allen, J.M.; Lawson, M.A.; Mailing, L.J.; Woods, J.A.; Johnson, R.W. Butyrate and Dietary Soluble Fiber Improve Neuroinflammation Associated with Aging in Mice. Front. Immunol. 2018, 9, 1832. [Google Scholar] [CrossRef]
- Chang, G.; Ma, N.; Zhang, H.; Wang, Y.; Huang, J.; Liu, J.; Dai, H.; Shen, X. Sodium butyrate modulates mucosal inflammation injury mediated by GPR41/43 in the cecum of goats fed a high concentration diet. Front. Physiol. 2019, 10, 1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Food | Oral Gavage |
|---|---|---|
| Control | Basal diet | 0.9% saline |
| DFC | Basal diet added 10% DFC | 0.9% saline |
| LTL1361 | Basal diet | 2 × 108 cfu LTL1361/mice per day |
| DFC + LTL1361 | Basal diet added 10% DFC | 2 × 108 cfu LTL1361/mice per day |
| Bacteria | Genbank Access No. | Forward (F) and Reversed (R) Primer Sequence (5′–3′) | Product Length (bp) |
|---|---|---|---|
| Caspase-3 | NM_009810.3 | F: GTCATCTCGCTCTGGTACGG R: CACACACACAAAGCTGCTCC | 169 |
| Bcl-2 | NM_009741.5 | F: TACGAGTGGGATGCTGGAGA R: CGGTAGCGACGAGAGAAGTC | 236 |
| GPR41 | NM_001033316.2 | F: CGGCTCACTGTAGTGTGGTT R: AGTCGTACAGGCAGGAGGAT | 127 |
| GPR43 | NM_001168509.1 | F: TCCTTGATCCTCACGGCCTA R: TTGGATGCTGCTTCCACGAT | 194 |
| ZO-1 | D14340.1 | F: TGTGGATTTACCCGTCAGCC R: AGGACGGCCTCTTCCCTTAT | 267 |
| Claudin-1 | NM_016674.4 | F: CTCCTGTCCCCGGAAAACAA R: CAGAGGGAAGCAGCAGTTCA | 311 |
| β-actin | NM_007393.5 | F: TACTGCTCTGGCTCCTAGCA R: CGGACTCATCGTACTCCTGC | 146 |
| Group | Escape Latency (s) | ||||
|---|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |
| Control | 47.4 ± 7.3 a | 44.7 ± 6.4 a | 40.5 ± 5.6 b | 38.9 ± 8.0 b | 38.1 ± 4.7 b |
| DFC | 46.1 ± 7.6 a | 41.4 ± 7.5 a | 35.2 ± 6.6 ab | 31.0 ± 5.8 ab | 29.4 ± 6.7 a* |
| LTL1361 | 43.2 ± 3.4 a | 38.5 ± 6.4 a | 35.3 ± 5.3 ab | 31.7 ± 5.1 ab | 27.2 ± 7.0 a* |
| DFC + LTL1361 | 42.0 ± 7.6 a | 36.1 ± 10.7 a | 28.0 ± 5.6 a | 25.8 ± 7.0 a | 22.7 ± 6.9 a** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, M.; Li, H.; Fu, Z.; Li, Q. Centenarian-Sourced Lactobacillus casei Combined with Dietary Fiber Complex Ameliorates Brain and Gut Function in Aged Mice. Nutrients 2022, 14, 324. https://doi.org/10.3390/nu14020324
Ren M, Li H, Fu Z, Li Q. Centenarian-Sourced Lactobacillus casei Combined with Dietary Fiber Complex Ameliorates Brain and Gut Function in Aged Mice. Nutrients. 2022; 14(2):324. https://doi.org/10.3390/nu14020324
Chicago/Turabian StyleRen, Minhong, He Li, Zhen Fu, and Quanyang Li. 2022. "Centenarian-Sourced Lactobacillus casei Combined with Dietary Fiber Complex Ameliorates Brain and Gut Function in Aged Mice" Nutrients 14, no. 2: 324. https://doi.org/10.3390/nu14020324