
Identification of a Hydroxygallic Acid Derivative, Zingibroside R1 and a Sterol Lipid as Potential Active Ingredients of Cuscuta chinensis Extract That Has Neuroprotective and Antioxidant Effects in Aged Caenorhabditis elegans

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. TCM Extracts Preparation, Fractionation and Purification of Single Compounds
2.2. UPLC-MS/MS Analysis of C. chinensis and E. ulmoides Extracts and Fractions
2.3. Caenorhabditis elegans Maintenance
2.4. The Treatment of C. elegans
2.5. Chemotaxis Assay
2.6. Learning and Short-Term Associative Memory
2.7. Behavioral Responses of C. elegans to Mechanical Stimuli
2.8. Survival Assays under Stress Conditions
2.9. Reactive Oxygen Species (ROS) Measurements
2.10. Reproduction Assay
2.11. Lifespan Assay
2.12. Swimming Behavior
2.13. Statistical Analysis
3. Results
3.1. C. chinensis and E. ulmoides Did Not Modify Chemotactic Abilities
3.2. C. chinensis Improved the Short-Term Associative Memory
3.3. Cuscuta chinensis Increased the Mechanosensory Response of C. elegans
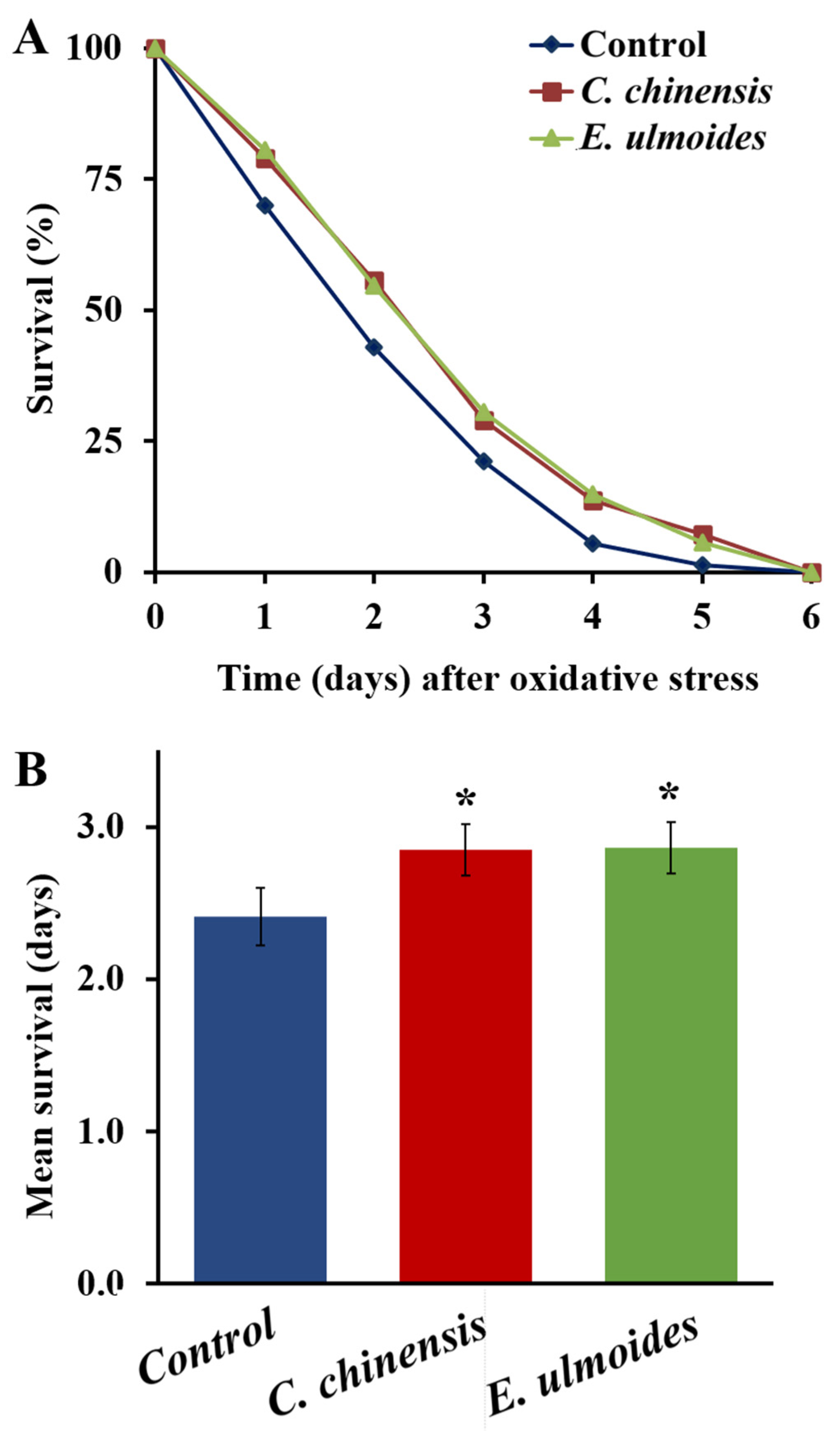
3.4. C. chinensis and E. ulmoides Increased the Oxidative Stress Resistance
3.5. TCM Extracts Decreased the Endogenous ROS Level
3.6. Only One C. chinensis Fraction Improved Locomotion, Mechanosensation as well as Oxidative Stress Resistance
3.7. A Hydroxygallic Acid Derivative Is the Most Abundant Component in C. chinensis Seeds
3.8. Fractions Enriched in Astragalin and Zingibroside R1 Extend the Lifespan of C. elegans
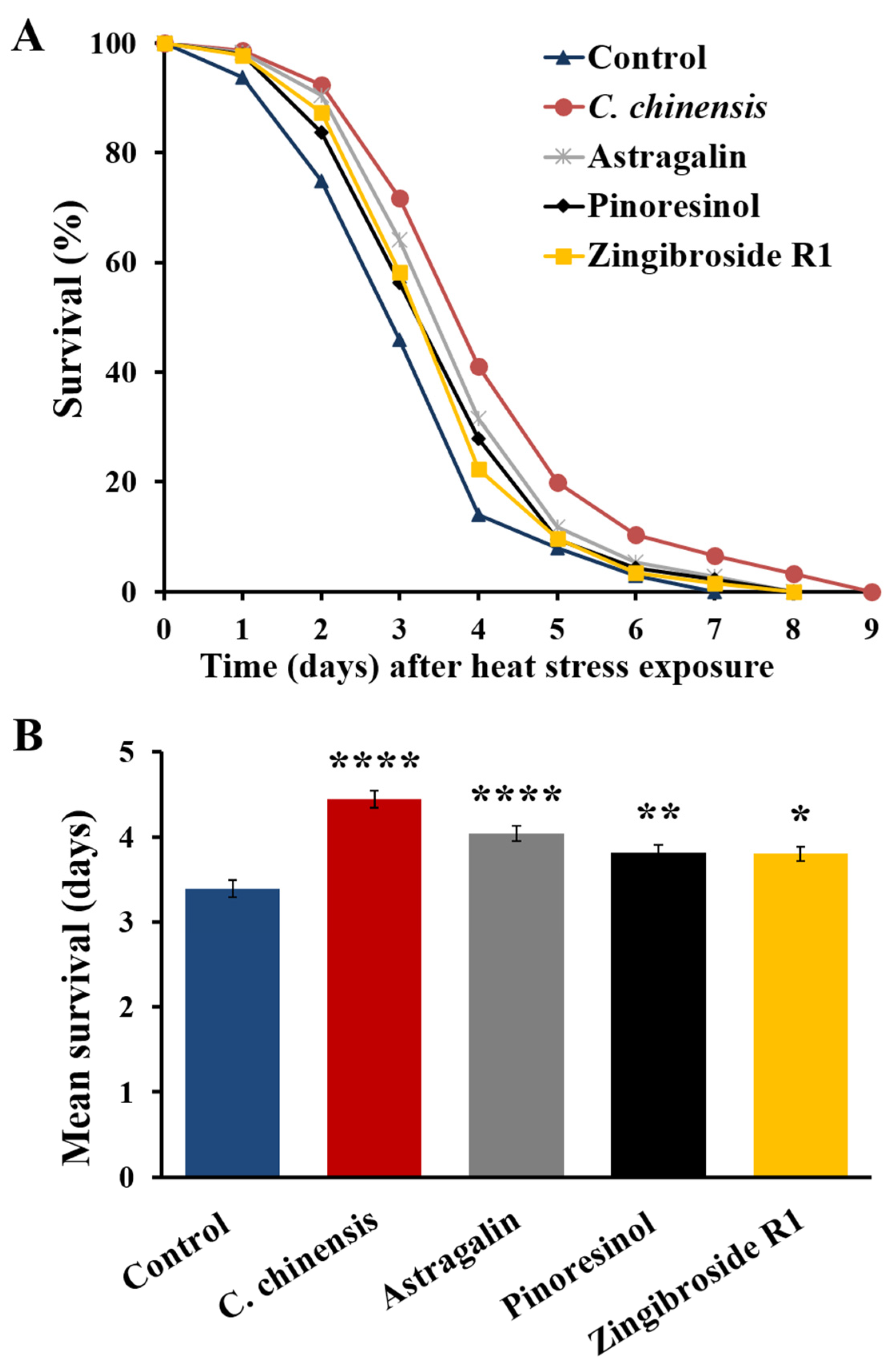
3.9. Fractions Enriched in Astragalin, Pinoresinol and Zingibroside R1 Increased Heat Stress Resistance of C. elegans
3.10. Fraction Enriched in Zingibroside R1 Improved Swimming Behaviour
4. Discussion
4.1. Neuroprotective Potential of C. Chinensis
4.2. Antioxidative Features of C. chinensis and E. ulmoides
4.3. A Hydroxygallic Acid Derivative and a Sterol Lipid Are Potential Triggers for the C. chinensis-Induced Increase of Physical and Cognitive Fitness
4.4. Several Bioactive Compounds Could Be the Underlying Cause of Enhanced Healthspan during C. chinensis and E. ulmoides Treatment
4.5. Zingibroside R1 Mirrors the Beneficial Actions of C. chinensis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arancio, O.; Chao, M.V. Neurotrophins, synaptic plasticity and dementia. Curr. Opin. Neurobiol. 2007, 17, 325–330. [Google Scholar] [CrossRef]
- Gispen, W.H.; Biessels, G.-J. Cognition and synaptic plasticity in diabetes mellitus. Trends Neurosci. 2000, 23, 542–549. [Google Scholar] [CrossRef]
- Shankar, G.M.; Li, S.; Mehta, T.H.; Garcia-Munoz, A.; Shepardson, N.E.; Smith, I.; Brett, F.M.; Farrell, M.A.; Rowan, M.J.; Lemere, C.A.; et al. Amyloid-β protein dimers isolated directly from alzheimer’s brains impair synaptic plasticity and memory. Nat. Med. 2008, 14, 837–842. [Google Scholar] [CrossRef] [Green Version]
- Crawford, Z.; San-Miguel, A. An inexpensive programmable optogenetic platform for controlled neuronal activation regimens in C. elegans. APL Bioeng. 2020, 4, 016101. [Google Scholar] [CrossRef] [Green Version]
- Bellantuono, I. Find Drugs that Delay Many Diseases of Old Age; Nature Publishing Group: Berlin, Germany, 2018. [Google Scholar]
- McDade, E.; Bateman, R.J. Stop alzheimer’s before it starts. Nature 2017, 547, 153–155. [Google Scholar] [CrossRef]
- Kepchia, D.; Currais, A.; Dargusch, R.; Finley, K.; Schubert, D.; Maher, P. Geroprotective effects of alzheimer’s disease drug candidates. Aging 2021, 13, 3269–3289. [Google Scholar] [CrossRef]
- Maresova, P.; Mohelska, H.; Dolejs, J.; Kuca, K. Socio-economic aspects of Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 903–911. [Google Scholar] [CrossRef]
- Fitzpatrick, A.L.; Kuller, L.H.; Ives, D.G.; Lopez, O.L.; Jagust, W.; Breitner, J.C.S.; Jones, B.; Lyketsos, C.; Dulberg, C. Incidence and prevalence of dementia in the cardiovascular health study. J. Am. Geriatr. Soc. 2004, 52, 195–204. [Google Scholar] [CrossRef]
- Evans, D.A.; Bennett, D.A.; Wilson, R.S.; Bienias, J.L.; Morris, M.C.; Scherr, P.A.; Hebert, L.E.; Aggarwal, N.; Beckett, L.A.; Joglekar, R.; et al. Incidence of alzheimer disease in a biracial urban community: Relation to apolipoprotein e allele status. Arch. Neurol. 2003, 60, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Alzheimer’s Association. 2020 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2020, 16, 391–460. [Google Scholar] [CrossRef]
- Miranda-Vizuete, A.; Veal, E.A. Caenorhabditis elegans as a model for understanding ros function in physiology and disease. Redox Biol. 2017, 11, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Labuschagne, C.F.; Brenkman, A.B. Current methods in quantifying ros and oxidative damage in caenorhabditis elegans and other model organism of aging. Ageing Res. Rev. 2013, 12, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Cadenas, E.; Symons, M.C.R.; Scott, G.; Norman, R.O.C.; Hill, H.A.O. Oxidative stress: Damage to intact cells and organs. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1985, 311, 617–631. [Google Scholar] [PubMed]
- Qu, F.; Zhang, Q.; Dai, M.; He, Y.; Wu, J.; Zhang, X.; Zhu, Y.; Gu, Y.; Wang, F.; Xu, X. An evaluation survey of traditional chinese medicine learning among international students majoring in conventional medicine: A study from a university in china. BMC Complement. Med. Ther. 2021, 21, 16. [Google Scholar] [CrossRef]
- Sayed, S.; Siems, K.; Schmitz-Linneweber, C.; Luyten, W.; Saul, N. Enhanced healthspan in Caenorhabditis elegans treated with extracts from the traditional chinese medicine plants Cuscuta chinensis lam. and Eucommia ulmoides oliv. Front. Pharmacol. 2021, 12, 604435. [Google Scholar] [CrossRef]
- Hou, J.-J.; Zhang, J.-Q.; Yao, C.-L.; Bauer, R.; Khan, I.A.; Wu, W.-Y.; Guo, D.-a. Deeper chemical perceptions for better traditional chinese medicine standards. Engineering 2019, 5, 83–97. [Google Scholar] [CrossRef]
- Gao, H.; Wang, Z.; Li, Y.; Qian, Z. Overview of the quality standard research of traditional chinese medicine. Front. Med. 2011, 5, 195–202. [Google Scholar] [CrossRef]
- Lin, H.-b.; Lin, J.-q.; Lu, N.; Lin, J.-q. Study of quality control on Cuscuta chinensis and C. australia. J. Chin. Med. Mater. 2007, 30, 1446–1449. [Google Scholar]
- Donnapee, S.; Li, J.; Yang, X.; Ge, A.-h.; Donkor, P.O.; Gao, X.-m.; Chang, Y.-x. Cuscuta chinensis lam.: A systematic review on ethnopharmacology, phytochemistry and pharmacology of an important traditional herbal medicine. J. Ethnopharmacol. 2014, 157, 292–308. [Google Scholar] [CrossRef]
- Alseekh, S.; Tohge, T.; Wendenberg, R.; Scossa, F.; Omranian, N.; Li, J.; Kleessen, S.; Giavalisco, P.; Pleban, T.; Mueller-Roeber, B.; et al. Identification and mode of inheritance of quantitative trait loci for secondary metabolite abundance in tomato. Plant Cell 2015, 27, 485–512. [Google Scholar] [CrossRef] [Green Version]
- Alseekh, S.; Aharoni, A.; Brotman, Y.; Contrepois, K.; D’Auria, J.; Ewald, J.; Ewald, J.C.; Fraser, P.D.; Giavalisco, P.; Hall, R.D.; et al. Mass spectrometry-based metabolomics: A guide for annotation, quantification and best reporting practices. Nat. Methods 2021, 18, 747–756. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans. In C. elegans: A Practical Approach; Hope, I., Ed.; Oxford University Press: Oxford, UK, 1999; pp. 51–67. [Google Scholar]
- Scerbak, C.; Vayndorf, E.M.; Hernandez, A.; McGill, C.; Taylor, B.E. Mechanosensory neuron aging: Differential trajectories with lifespan-extending alaskan berry and fungal treatments in Caenorhabditis elegans. Front. Aging Neurosci. 2016, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Duangjan, C.; Rangsinth, P.; Gu, X.; Wink, M.; Tencomnao, T. Lifespan extending and oxidative stress resistance properties of a leaf extracts from anacardium occidentale l. In Caenorhabditis elegans. Oxid. Med. Cell. Longev. 2019, 2019, 9012396. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; An, H.S.; Jung, Y.W.; Lee, E.J.; Lee, H.Y.; Choi, E.S.; An, S.W.; Son, H.; Lee, S.J.; Kim, J.B.; et al. Korean mistletoe (Viscum album coloratum) extract extends the lifespan of nematodes and fruit flies. Biogerontology 2014, 15, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Hosono, R. Sterilization and growth inhibition of Caenorhabditis elegans by 5-fluorodeoxyuridine. Exp. Gerontol. 1978, 13, 369–373. [Google Scholar] [CrossRef]
- Margie, O.; Palmer, C.; Chin-Sang, I. C. elegans chemotaxis assay. J. Vis. Exp. 2013, e50069. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, A.L.; Ashraf, J.M.; Corces-Zimmerman, M.R.; Landis, J.N.; Murphy, C.T. Insulin signaling and dietary restriction differentially influence the decline of learning and memory with age. PLoS Biol. 2010, 8, e1000372. [Google Scholar] [CrossRef] [Green Version]
- Chalfie, M.; Hart, A.C.; Rankin, C.H.; Goodman, M.B. Assaying mechanosensation. In Wormbook: The Online Review of C. elegans Biology; WormBook: Pasadena, CA, USA, 2014. [Google Scholar] [CrossRef]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An anthocyanin-rich extract of acai (Euterpe precatoria mart.) increases stress resistance and retards aging-related markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [CrossRef]
- Lim, H.J.; Han, Y.T.; Ahn, J.H.; Jeon, Y.D.; Jeon, H.; Cha, D.S. Longevity effects of hispidol in Caenorhabditis elegans. Biofactors 2020, 46, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Seo, H.W.; Lee, M.H.; Kim, D.K.; Jeon, H.; Cha, D.S. Protocatechuic acid extends lifespan and increases stress resistance in Caenorhabditis elegans. Arch. Pharm. Res. 2014, 37, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Im, J.S.; Lee, H.N.; Oh, J.W.; Yoon, Y.J.; Park, J.S.; Park, J.W.; Kim, J.H.; Kim, Y.S.; Cha, D.S.; Jeon, H. Moringa oleifera prolongs lifespan via daf-16/foxo transcriptional factor in Caenorhabditis elegans. Nat. Prod. Sci. 2016, 22, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Back, P.; De Vos, W.H.; Depuydt, G.G.; Matthijssens, F.; Vanfleteren, J.R.; Braeckman, B.P. Exploring real-time in vivo redox biology of developing and aging Caenorhabditis elegans. Free Radic. Biol. Med. 2012, 52, 850–859. [Google Scholar] [CrossRef]
- McQuin, C.; Goodman, A.; Chernyshev, V.; Kamentsky, L.; Cimini, B.A.; Karhohs, K.W.; Doan, M.; Ding, L.; Rafelski, S.M.; Thirstrup, D. Cellprofiler 3.0: Next-generation image processing for biology. PLoS Biol. 2018, 16, e2005970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restif, C.; Ibáñez-Ventoso, C.; Vora, M.M.; Guo, S.; Metaxas, D.; Driscoll, M. Celest: Computer vision software for quantitative analysis of C. elegans swim behavior reveals novel features of locomotion. PLoS Comp. Biol. 2014, 10, e1003702. [Google Scholar] [CrossRef] [Green Version]
- Han, S.K.; Lee, D.; Lee, H.; Kim, D.; Son, H.G.; Yang, J.-S.; Lee, S.-J.V.; Kim, S. Oasis 2: Online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget 2016, 7, 56147–56152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torayama, I.; Ishihara, T.; Katsura, I. Caenorhabditis elegans integrates the signals of butanone and food to enhance chemotaxis to butanone. J. Neurosci. 2007, 27, 741. [Google Scholar] [CrossRef] [Green Version]
- Hedden, T.; Gabrieli, J.D.E. Insights into the ageing mind: A view from cognitive neuroscience. Nat. Rev. Neurosci. 2004, 5, 87–96. [Google Scholar] [CrossRef]
- Gregory, J.; Vengalasetti, Y.V.; Bredesen, D.E.; Rao, R.V. Neuroprotective herbs for the management of alzheimer’s disease. Biomolecules 2021, 11, 543. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, L.S.C.; Savulich, G.; Sahakian, B.J. Lifestyle use of drugs by healthy people for enhancing cognition, creativity, motivation and pleasure. Br. J. Pharmacol. 2017, 174, 3257–3267. [Google Scholar] [CrossRef]
- Lavretsky, H. Complementary and alternative medicine use for treatment and prevention of late-life mood and cognitive disorders. Aging Health 2009, 5, 61–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Garg, N.; Verma, D.; Rathi, P.; Sharma, V.; Kuca, K.; Prajapati, P.K. Indian medicinal plants as drug leads in neurodegenerative disorders. In Nutraceuticals in Brain Health and Beyond; Ghosh, D., Ed.; Academic Press: Cambridge, MA, USA, 2021; Chapter 4; pp. 31–45. [Google Scholar]
- Mahomoodally, M.F. Traditional medicines in Africa: An appraisal of ten potent african medicinal plants. Evid.-Based Complement. Altern. Med. 2013, 2013, 617459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yao, H.; Wang, J.; Li, Y.; Wang, Q. The role of chinese medicine in health maintenance and disease prevention: Application of constitution theory. Am. J. Chin. Med. 2019, 47, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.B.; Mukerjee, A.; Singh, S. Global approach for drug discovery and development from indian traditional medicine. In Evidence Based Validation of Traditional Medicines: A Comprehensive Approach; Mandal, S.C., Chakraborty, R., Sen, S., Eds.; Springer: Singapore, 2021; pp. 3–27. [Google Scholar]
- Howes, M.-J.R.; Houghton, P.J. Plants used in Chinese and Indian traditional medicine for improvement of memory and cognitive function. Pharmacol. Biochem. Behav. 2003, 75, 513–527. [Google Scholar] [CrossRef]
- Alzobaidi, N.; Quasimi, H.; Emad, N.A.; Alhalmi, A.; Naqvi, M. Bioactive compounds and traditional herbal medicine: Promising approaches for the treatment of dementia. Degener. Neurol. Neuromuscul. Dis. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [Green Version]
- Cerutti, D.T.; Levin, E.D. Cognitive impairment models using complementary species. In Animal Models of Cognitive Impairment; Levin, E.D., Buccafusco, J.J., Eds.; CRC Press/Taylor & Francis: New York, NY, USA, 2006; pp. 315–342. [Google Scholar]
- Arey, R.N.; Murphy, C.T. Conserved regulators of cognitive aging: From worms to humans. Behav. Brain Res. 2017, 322, 299–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardiel, E.L.; Rankin, C.H. An elegant mind: Learning and memory in Caenorhabditis elegans. Learn. Mem. 2010, 17, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Barclay, J.W.; Burgoyne, R.D.; Morgan, A. Using C. elegans to discover therapeutic compounds for ageing-associated neurodegenerative diseases. Chem. Cent. J. 2015, 9, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhang, B.; Lei, H.; Feng, Z.; Liu, J.; Hsu, A.-L.; Xu, X.Z.S. Functional aging in the nervous system contributes to age-dependent motor activity decline in C. elegans. Cell Metab. 2013, 18, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Lian Chew, Y.; Fan, X.; Götz, J.; Nicholas, H.R. Aging in the nervous system of Caenorhabditis elegans. Commun. Integr. Biol. 2013, 6, e25288. [Google Scholar] [CrossRef] [Green Version]
- Toth, M.L.; Melentijevic, I.; Shah, L.; Bhatia, A.; Lu, K.; Talwar, A.; Naji, H.; Ibanez-Ventoso, C.; Ghose, P.; Jevince, A.; et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. J. Neurosci. 2012, 32, 8778–8790. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, T.R.; Liu, Q.; Itoh, T.; Luo, S.; Hadwiger, G.; Vincent, R.; Wang, Z.W.; Fukuda, M.; Nonet, M.L. Regulation of synaptic transmission by rab-3 and rab-27 in Caenorhabditis elegans. Mol. Biol. Cell 2006, 17, 2617–2625. [Google Scholar] [CrossRef] [Green Version]
- Deems, D.A.; Doty, R.L.; Settle, R.G.; Moore-Gillon, V.; Shaman, P.; Mester, A.F.; Kimmelman, C.P.; Brightman, V.J.; Snow, J.B. Smell and taste disorders, a study of 750 patients from the university of pennsylvania smell and taste center. Arch. Otolaryngol.—Head Neck Surg. 1991, 117, 519–528. [Google Scholar] [CrossRef]
- Goelet, P.; Castellucci, V.F.; Schacher, S.; Kandel, E.R. The long and the short of long–term memory—A molecular framework. Nature 1986, 322, 419–422. [Google Scholar] [CrossRef]
- Lin, M.K.; Lee, M.S.; Huang, H.C.; Cheng, T.J.; Cheng, Y.D.; Wu, C.R. Cuscuta chinensis and C. campestris attenuate scopolamine-induced memory deficit and oxidative damage in mice. Molecules 2018, 23, 3060. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.; Du, S. Study on the anti-aging action of the extracts of Cuscuta chinensis in natural aging mice. China Pharm. 2010, 39, 3667–3669. [Google Scholar]
- Ji, Z.H.; Zhang, X.L.; Dong, L.F. Effect of the water extract of Cuscuta chinensis lam. On memory impairment in cerebral ischemic rats. Chin. J. Behav. Med. Brain Sci. 2006, 15, 681–682. [Google Scholar]
- Pandey, R.; Gupta, S.; Shukla, V.; Tandon, S.; Shukla, V. Antiaging, antistress and ros scavenging activity of crude extract of Ocimum sanctum (L.) in Caenorhabditis elegans (maupas, 1900). Indian J. Exp. Biol. 2013, 51, 515–521. [Google Scholar]
- Brown, M.K.; Evans, J.L.; Luo, Y. Beneficial effects of natural antioxidants egcg and α-lipoic acid on life span and age-dependent behavioral declines in Caenorhabditis elegans. Pharmacol. Biochem. Behav. 2006, 85, 620–628. [Google Scholar] [CrossRef]
- Akhoon, B.A.; Pandey, S.; Tiwari, S.; Pandey, R. Withanolide a offers neuroprotection, ameliorates stress resistance and prolongs the life expectancy of Caenorhabditis elegans. Exp. Gerontol. 2016, 78, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Phulara, S.C.; Shukla, V.; Tiwari, S.; Pandey, R. Bacopa monnieri promotes longevity in Caenorhabditis elegans under stress conditions. Pharmacogn. Mag. 2015, 11, 410–416. [Google Scholar]
- Rathor, L.; Pant, A.; Awasthi, H.; Mani, D.; Pandey, R. An antidiabetic polyherbal phytomedicine confers stress resistance and extends lifespan in Caenorhabditis elegans. Biogerontology 2017, 18, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Dilberger, B.; Baumanns, S.; Schmitt, F.; Schmiedl, T.; Hardt, M.; Wenzel, U.; Eckert, G.P. Mitochondrial oxidative stress impairs energy metabolism and reduces stress resistance and longevity of C. elegans. Oxid. Med. Cell. Longev. 2019, 2019, 6840540. [Google Scholar] [CrossRef] [PubMed]
- Sampayo, J.N.; Olsen, A.; Lithgow, G.J. Oxidative stress in Caenorhabditis elegans: Protective effects of superoxide dismutase/catalase mimetics. Aging Cell 2003, 2, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Voth, W.; Jakob, U. Stress-activated chaperones: A first line of defense. Trends Biochem. Sci. 2017, 42, 899–913. [Google Scholar] [CrossRef]
- Gao, J.-M.; Li, R.; Zhang, L.; Jia, L.-L.; Ying, X.-X.; Dou, D.-Q.; Li, J.-C.; Li, H.-B. Cuscuta chinensis seeds water extraction protecting murine osteoblastic MC3T3-E1 cells against tertiary butyl hydroperoxide induced injury. J. Ethnopharmacol. 2013, 148, 587–595. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Wang, Y.-L.; Li, Q.-L.; Yang, L. Improved antimelanogenesis and antioxidant effects of polysaccharide from Cuscuta chinensis lam seeds after enzymatic hydrolysis. Braz. J. Med. Biol. Res. 2018, 51. [Google Scholar] [CrossRef]
- Mo, H.; Zhang, N.; Li, H.; Li, F.; Pu, R. Beneficial effects of Cuscuta chinensis extract on glucocorticoid-induced osteoporosis through modulation of rankl/opg signals. Braz. J. Med. Biol. Res. 2019, 52. [Google Scholar] [CrossRef]
- Wang, J.H.; Liu, D.M.; Liu, Y.L.; Li, C.L.; Cheng, Y.Y.; Li, Y. Antioxidant capacity and polyphenolic content of Eucommia ulmoides oliv leaf extract. Adv. Mater. Res. 2012, 396–398, 1349–1352. [Google Scholar] [CrossRef]
- Hou, P.; Wang, Q.; Qi, W.; Zhang, Y.; Xie, J. Comprehensive determination of seven polyphenols in Eucommia ulmoides and its anti-oxidative stress activity in C. elegans. J. Food Meas. Charact. 2019, 13, 2903–2909. [Google Scholar] [CrossRef]
- Ding, H.; Cao, A.; Li, H.; Zhao, Y.; Feng, J. Effects of Eucommia ulmoides leaf extracts on growth performance, antioxidant capacity and intestinal function in weaned piglets. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1169–1177. [Google Scholar] [CrossRef]
- Yen, F.-L.; Wu, T.-H.; Lin, L.-T.; Lin, C.-C. Hepatoprotective and antioxidant effects of Cuscuta chinensis against acetaminophen-induced hepatotoxicity in rats. J. Ethnopharmacol. 2007, 111, 123–128. [Google Scholar] [CrossRef]
- Ristow, M. Unraveling the truth about antioxidants: Mitohormesis explains ros-induced health benefits. Nat. Med. 2014, 20, 709–711. [Google Scholar] [CrossRef]
- Ristow, M.; Schmeisser, K. Mitohormesis: Promoting health and lifespan by increased levels of reactive oxygen species (ros). Dose Response 2014, 12, 288–341. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Phenolic compounds of pomegranate byproducts (outer skin, mesocarp, divider membrane) and their antioxidant activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef]
- Fathoni, A.; Saepudin, E.; Cahyana, A.H.; Rahayu, D.U.C.; Haib, J. Identification of nonvolatile compounds in clove (Syzygium aromaticum) from manado. AIP Conf. Proc. 2017, 1862, 030079. [Google Scholar]
- El-Hawary, S.S.; Mubarek, M.M.; Lotfy, R.A.; Hassan, A.R.; Sobeh, M.; Okba, M.M. Validation of antidiabetic potential of Gymnocarpos decandrus forssk. Nat. Prod. Res. 2021, 35, 5954–5959. [Google Scholar] [CrossRef]
- Saul, N.; Pietsch, K.; Sturzenbaum, S.R.; Menzel, R.; Steinberg, C.E. Diversity of polyphenol action in Caenorhabditis elegans: Between toxicity and longevity. J. Nat. Prod. 2011, 74, 1713–1720. [Google Scholar] [CrossRef]
- Choubey, S.; Goyal, S.; Varughese, L.R.; Kumar, V.; Sharma, A.K.; Beniwal, V. Probing gallic acid for its broad spectrum applications. Mini Rev. Med. Chem. 2018, 18, 1283–1293. [Google Scholar] [CrossRef]
- Li, X.; Lu, H.-Y.; Jiang, X.-W.; Yang, Y.; Xing, B.; Yao, D.; Wu, Q.; Xu, Z.-H.; Zhao, Q.-C. Cinnamomum cassia extract promotes thermogenesis during exposure to cold via activation of brown adipose tissue. J. Ethnopharmacol. 2021, 266, 113413. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghari, B.; Yazdiniapour, Z.; Sadeghi, M.; Akbari, M.; Troiano, R.; Lanzotti, V. Cinnamic acid derivatives from welsh onion (Allium fistulosum) and their antibacterial and cytotoxic activities. Phytochem. Anal. 2021, 32, 84–90. [Google Scholar] [CrossRef]
- Buxton, T.; Takahashi, S.; Eddy Doh, A.-M.; Baffoe-Ansah, J.; Owusu, E.O.; Kim, C.-S. Insecticidal activities of cinnamic acid esters isolated from Ocimum gratissimum L. and Vitellaria paradoxa gaertn leaves against Tribolium castaneum hebst (coleoptera: Tenebrionidae). Pest Manag. Sci. 2020, 76, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Mancilla-Montelongo, G.; Castañeda-Ramírez, G.S.; Torres-Acosta, J.F.d.J.; Sandoval-Castro, C.A.; Borges-Argáez, R. Evaluation of cinnamic acid and six analogues against eggs and larvae of Haemonchus contortus. Vet. Parasitol. 2019, 270, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.-S.; Hou, J.-W.; Liu, Y.; Ding, Y.; Zhang, Y.; Li, L.; Zhang, T. Design, synthesis and evaluation of novel cinnamic acid derivatives bearing n-benzyl pyridinium moiety as multifunctional cholinesterase inhibitors for Alzheimer’s disease. J. Enzym. Inhib. Med. Chem. 2017, 32, 776–788. [Google Scholar] [CrossRef]
- Hu, S.; Yang, X.; Xue, J.; Chen, X.; Bai, X.-h.; Yu, Z.-h. Graphene/dodecanol floating solidification microextraction for the preconcentration of trace levels of cinnamic acid derivatives in traditional chinese medicines. J. Sep. Sci. 2017, 40, 2959–2966. [Google Scholar] [CrossRef]
- Savych, A.; Marchyshyn, S.; Kyryliv, M.; Bekus, I. Cinnamic acid and its derivatives in the herbal mixtures and their antidiabetic activity. Farmacia 2021, 69, 595–601. [Google Scholar] [CrossRef]
- Aderibigbe, S.; Oyeniran, O.; Idowu, S. Anthelmintic activity of Nauclea diderrichii leaf extracts and fractions against adult Haemonchus placei. Niger. J. Pharm. Res. 2020, 16, 81–86. [Google Scholar] [CrossRef]
- Mazzone, G.; Russo, N.; Toscano, M. Antioxidant properties comparative study of natural hydroxycinnamic acids and structurally modified derivatives: Computational insights. Comput. Theor. Chem. 2016, 1077, 39–47. [Google Scholar] [CrossRef]
- Li, P.-X.; Li, Y.-M.; Mu, W.-W.; Sun, Y.-L.; Li, Y.; Yang, J.; Liu, R.-M.; Liu, G.-Y. Cinnamic acid/β-ionone hybrids: Synthesis and in vitro anticancer activity evaluation. Mon. Chem./Chem. Mon. 2021, 152, 863–870. [Google Scholar] [CrossRef]
- Fukuda, H.; Nakamura, S.; Chisaki, Y.; Takada, T.; Toda, Y.; Murata, H.; Itoh, K.; Yano, Y.; Takata, K.; Ashihara, E. Daphnetin inhibits invasion and migration of lm8 murine osteosarcoma cells by decreasing rhoa and cdc42 expression. Biochem. Biophys. Res. Commun. 2016, 471, 63–67. [Google Scholar] [CrossRef]
- Shen, L.; Zhou, T.; Wang, J.; Sang, X.; Lan, L.; Luo, L.; Yin, Z. Daphnetin reduces endotoxin lethality in mice and decreases lps-induced inflammation in raw264.7 cells via suppressing jak/stats activation and ros production. Inflamm. Res. 2017, 66, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Amari, N.O.; Razafimandimby, B.; Auberon, F.; Azoulay, S.; Fernandez, X.; Berkani, A.; Bouchara, J.-P.; Landreau, A. Antifungal and antiaging evaluation of aerial part extracts of Thymelaea hirsuta (L.) endl. Nat. Prod. Commun. 2021, 16, 1934578X20987932. [Google Scholar] [CrossRef]
- Li, Y.; Yang, D.; Jia, Y.; He, L.; Li, J.; Yu, C.; Liao, C.; Yu, Z.; Zhang, C. Research note: Anti-inflammatory effects and antiviral activities of baicalein and chlorogenic acid against infectious bursal disease virus in embryonic eggs. Poult. Sci. 2021, 100, 100987. [Google Scholar] [CrossRef]
- Park, J.B. 5-caffeoylquinic acid and caffeic acid orally administered suppress p-selectin expression on mouse platelets. J. Nutr. Biochem. 2009, 20, 800–805. [Google Scholar] [CrossRef]
- Wan, F.; Cai, X.; Wang, M.; Chen, L.; Zhong, R.; Liu, L.; Yi, B.; Hou, F.; Zhang, H. Chlorogenic acid supplementation alleviates dextran sulfate sodium (dss)-induced colitis via inhibiting inflammatory responses and oxidative stress, improving gut barrier integrity and nrf-2/ho-1 pathway. J. Funct. Foods 2021, 87, 104808. [Google Scholar] [CrossRef]
- Li, Y.; Shi, W.; Li, Y.; Zhou, Y.; Hu, X.; Song, C.; Ma, H.; Wang, C.; Li, Y. Neuroprotective effects of chlorogenic acid against apoptosis of pc12 cells induced by methylmercury. Environ. Toxicol. Pharmacol. 2008, 26, 13–21. [Google Scholar] [CrossRef]
- Fernandes, M.Y.D.; Dobrachinski, F.; Silva, H.B.; Lopes, J.P.; Gonçalves, F.Q.; Soares, F.A.A.; Porciúncula, L.O.; Andrade, G.M.; Cunha, R.A.; Tomé, A.R. Neuromodulation and neuroprotective effects of chlorogenic acids in excitatory synapses of mouse hippocampal slices. Sci. Rep. 2021, 11, 10488. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.I.; Kwon, J.H.; Park, K.H.; Oh, M.H.; Kim, M.H.; Kim, H.H.; Cho, S.H.; Chung, E.K.; Ha, S.Y.; Lee, M.W. The antioxidant and anti-inflammatory effects of phenolic compounds isolated from the root of Rhodiola sachalinensis a. Bor. Molecules 2012, 17, 11484–11494. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Hwang, K.-A.; Choi, K.-C. Treatment with kaempferol suppresses breast cancer cell growth caused by estrogen and triclosan in cellular and xenograft breast cancer models. J. Nutr. Biochem. 2016, 28, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.-T.; Tsai, Y.-C.; Wu, H.-C.; Ho, Y.-J.; Chen, Y.-S.; Yao, C.-H.; Yao, C.-H. Radiosensitization of non-small cell lung cancer by kaempferol. Oncol. Rep. 2015, 34, 2351–2356. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Park, J.H.Y. Kaempferol induces cell cycle arrest in ht-29 human colon cancer cells. J. Cancer Prev. 2013, 18, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Dang, Q.; Song, W.; Xu, D.; Ma, Y.; Li, F.; Zeng, J.; Zhu, G.; Wang, X.; Chang, L.S.; He, D.; et al. Kaempferol suppresses bladder cancer tumor growth by inhibiting cell proliferation and inducing apoptosis. Mol. Carcinog. 2015, 54, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Mijangos-Ramos, I.F.; Zapata-Estrella, H.E.; Ruiz-Vargas, J.A.; Escalante-Erosa, F.; Gómez-Ojeda, N.; García-Sosa, K.; Cechinel-Filho, V.; Meira-Quintão, N.L.; Peña-Rodríguez, L.M. Bioactive dicaffeoylquinic acid derivatives from the root extract of Calea urticifolia. Rev. Bras. Farmacogn. 2018, 28, 339–343. [Google Scholar] [CrossRef]
- Yang, P.-F.; Feng, Z.-M.; Yang, Y.-N.; Jiang, J.-S.; Zhang, P.-C. Neuroprotective caffeoylquinic acid derivatives from the flowers of Chrysanthemum morifolium. J. Nat. Prod. 2017, 80, 1028–1033. [Google Scholar] [CrossRef]
- Wang, W.W.; Han, R.; He, H.J.; Li, J.; Chen, S.Y.; Gu, Y.; Xie, C. Administration of quercetin improves mitochondria quality control and protects the neurons in 6-ohda-lesioned Parkinson’s disease models. Aging 2021, 13, 11738–11751. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Quispe, C.; Hossain, R.; Islam, M.T.; Al-Harrasi, A.; Al-Rawahi, A.; Martorell, M.; Mamurova, A.; Seilkhan, A.; Altybaeva, N.; et al. Neuropharmacological effects of quercetin: A literature-based review. Front. Pharmacol. 2021, 12, 665031. [Google Scholar] [CrossRef]
- Li, H.; Chen, F.-J.; Yang, W.-L.; Qiao, H.-Z.; Zhang, S.-J. Quercetin improves cognitive disorder in aging mice by inhibiting nlrp3 inflammasome activation. Food Funct. 2021, 12, 717–725. [Google Scholar] [CrossRef]
- Masek, A.; Latos, M.; Piotrowska, M.; Zaborski, M. The potential of quercetin as an effective natural antioxidant and indicator for packaging materials. Food Packag. Shelf Life 2018, 16, 51–58. [Google Scholar] [CrossRef]
- Jaisinghani, R.N. Antibacterial properties of quercetin. Microbiol. Res. 2017, 8, 6877. [Google Scholar] [CrossRef] [Green Version]
- Osonga, F.J.; Akgul, A.; Miller, R.M.; Eshun, G.B.; Yazgan, I.; Akgul, A.; Sadik, O.A. Antimicrobial activity of a new class of phosphorylated and modified flavonoids. ACS Omega 2019, 4, 12865–12871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauf, A.; Imran, M.; Khan, I.A.; ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Kampkötter, A.; Timpel, C.; Zurawski, R.F.; Ruhl, S.; Chovolou, Y.; Proksch, P.; Wätjen, W. Increase of stress resistance and lifespan of Caenorhabditis elegans by quercetin. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 149, 314–323. [Google Scholar] [CrossRef]
- Saul, N.; Pietsch, K.; Menzel, R.; Steinberg, C.E. Quercetin-mediated longevity in Caenorhabditis elegans: Is daf-16 involved? Mech. Ageing Dev. 2008, 129, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Liu, D.; Liu, Y.L.; Liu, B.; Chen, X.H. Quercetin and its mixture increase the stress resistance of Caenorhabditis elegans to uv-b. Int. J. Environ. Res. Public Health 2020, 17, 1572. [Google Scholar] [CrossRef] [PubMed]
- Issinger, O.-G.; Guerra, B. Phytochemicals in cancer and their effect on the pi3k/akt-mediated cellular signalling. Biomed. Pharmacother. 2021, 139, 111650. [Google Scholar] [CrossRef] [PubMed]
- Matboli, M.; Saad, M.; Hasanin, A.H.; Saleh, L.A.; Baher, W.; Bekhet, M.M.; Eissa, S. New insight into the role of isorhamnetin as a regulator of insulin signaling pathway in type 2 diabetes mellitus rat model: Molecular and computational approach. Biomed. Pharmacother. 2021, 135, 111176. [Google Scholar] [CrossRef] [PubMed]
- Kandakumar, S.; Manju, D. Pharmacological applications of isorhamnetin: A short review. Int. J. Trend Sci. Res. Dev. 2017, 1, 672–678. [Google Scholar] [CrossRef]
- Zaki Rashed, K.N. Biological activities of isorhamnetin: A review. Plantae Sci. 2020, 3, 78–81. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, Z.; Li, S.; Ye, X.; Li, X.; He, K. Synergy effects of herb extracts: Pharmacokinetics and pharmacodynamic basis. Fitoterapia 2014, 92, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Riaz, A.; Rasul, A.; Hussain, G.; Zahoor, M.K.; Jabeen, F.; Subhani, Z.; Younis, T.; Ali, M.; Sarfraz, I.; Selamoglu, Z. Astragalin: A bioactive phytochemical with potential therapeutic activities. Adv. Pharmacol. Sci. 2018, 2018, 9794625. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Qi, Y.; Kim, R.; Song, J.; Kim, H.; Kim, H.Y.; Jang, D.S.; Kang, K.S. Methyl caffeate isolated from the flowers of Prunus persica (L.) batsch enhances glucose-stimulated insulin secretion. Biomolecules 2021, 11, 279. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Lee, J.S.; Quilantang, N.G.; Jacinto, S.D.; Lee, S. Determination of afzelin and astragalin from Lespedeza cuneata on aldose reductase inhibition. J. Chromatogr. Sci. 2021, 59, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Oldoni, T.L.C.; Merlin, N.; Bicas, T.C.; Prasniewski, A.; Carpes, S.T.; Ascari, J.; de Alencar, S.M.; Massarioli, A.P.; Bagatini, M.D.; Morales, R. Antihyperglycemic activity of crude extract and isolation of phenolic compounds with antioxidant activity from Moringa oleifera lam. Leaves grown in southern brazil. Food Res. Int. 2021, 141, 110082. [Google Scholar] [CrossRef]
- Kwon, H.-J.; Park, Y.-D. Determination of astragalin and astragaloside content in radix astragali using high-performance liquid chromatography coupled with pulsed amperometric detection. J. Chromatogr. 2012, 1232, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.; Mikhova, B.; Najdenski, H.; Tsvetkova, I.; Kostova, I. Chemical composition and antimicrobial activity of wild garlic Allium ursinum of bulgarian origin. Nat. Prod. Commun. 2009, 4, 1059–1062. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Yang, W.; Ye, M.; Wang, Q.; Guo, D. Differentiation of Cuscuta chinensis and Cuscuta australis by hplc-dad-ms analysis and hplc-uv quantitation. Planta Med. 2011, 77, 1950–1957. [Google Scholar] [CrossRef] [Green Version]
- Jia, Q.; Wang, T.; Wang, X.; Xu, H.; Liu, Y.; Wang, Y.; Shi, Q.; Liang, Q. Astragalin suppresses inflammatory responses and bone destruction in mice with collagen-induced arthritis and in human fibroblast-like synoviocytes. Front. Pharmacol. 2019, 10, 94. [Google Scholar] [CrossRef] [Green Version]
- Harikrishnan, H.; Jantan, I.; Alagan, A.; Haque, M.A. Modulation of cell signaling pathways by Phyllanthus amarus and its major constituents: Potential role in the prevention and treatment of inflammation and cancer. Inflammopharmacology 2020, 28, 1–18. [Google Scholar] [CrossRef]
- Luo, H.; Daddysman, M.K.; Rankin, G.O.; Jiang, B.-H.; Chen, Y.C. Kaempferol enhances cisplatin’s effect on ovarian cancer cells through promoting apoptosis caused by down regulation of cmyc. Cancer Cell Int. 2010, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Nguyen, H.T.; Choi, E.M.; Bae, K.H.; Kim, Y.H. Rhusonoside a, a new megastigmane glycoside from Rhus sylvestris, increases the function of osteoblastic mc3t3-e1 cells. Planta Med. 2009, 75, 158–162. [Google Scholar] [CrossRef]
- Liu, L.; Wang, D.; Qin, Y.; Xu, M.; Zhou, L.; Xu, W.; Liu, X.; Ye, L.; Yue, S.; Zheng, Q.; et al. Astragalin promotes osteoblastic differentiation in mc3t3-e1 cells and bone formation in vivo. Front. Endocrinol. 2019, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, W.-Y.; Xie, J.; Wang, Z.-L.; Wen, Y.-L.; Zhao, C.-C.; Tao, L.; Li, L.-F.; Tian, Y.; Sheng, J. Astragalin inhibits the proliferation and migration of human colon cancer hct116 cells by regulating the nf-κb signaling pathway. Front. Pharmacol. 2021, 12, 639256. [Google Scholar] [CrossRef] [PubMed]
- Rey, D.; Miranda Sulis, P.; Alves Fernandes, T.; Gonçalves, R.; Silva Frederico, M.J.; Costa, G.M.; Aragon, M.; Ospina, L.F.; Mena Barreto Silva, F.R. Astragalin augments basal calcium influx and insulin secretion in rat pancreatic islets. Cell Calcium 2019, 80, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Li, Y.; Chu, Q.; Liu, Y.; Ye, X.; Jiang, Y.; Zheng, X. Radix tetrastigma flavonoid ameliorates inflammation and prolongs the lifespan of Caenorhabditis elegans through jnk, p38 and nrf2 pathways. Free Radic. Res. 2019, 53, 562–573. [Google Scholar] [CrossRef]
- Lei, S.; Wu, S.; Wang, G.; Li, B.; Liu, B.; Lei, X. Pinoresinol diglucoside attenuates neuroinflammation, apoptosis and oxidative stress in a mice model with Alzheimer’s disease. Neuroreport 2021, 32, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zou, Z.; Wang, X.; Ji, X.; Yang, J. Pinoresinol diglucoside alleviates oxldl-induced dysfunction in human umbilical vein endothelial cells. Evid.-Based Complement. Altern. Med. 2016, 2016, 3124519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, B.; Lee, J.; Liu, Q.-H.; Woo, E.-R.; Lee, D.G. Antifungal effect of (+)-pinoresinol isolated from Sambucus williamsii. Molecules 2010, 15, 3507–3516. [Google Scholar] [CrossRef] [PubMed]
- Milder, I.E.; Arts, I.C.; van de Putte, B.; Venema, D.P.; Hollman, P.C. Lignan contents of dutch plant foods: A database including lariciresinol, pinoresinol, secoisolariciresinol and matairesinol. Br. J. Nutr. 2005, 93, 393–402. [Google Scholar] [CrossRef]
- Sok, D.-E.; Cui, H.S.; Kim, M.R. Isolation and boactivities of furfuran type lignan compounds from edible plants. Recent Pat. Food Nutr. Agric. 2009, 1, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Kwon, H.; Cho, E.; Jeon, J.; Kang, R.H.; Youn, K.; Jun, M.; Lee, Y.C.; Ryu, J.H.; Kim, D.H. The effects of pinoresinol on cholinergic dysfunction-induced memory impairments and synaptic plasticity in mice. Food Chem. Toxicol. 2019, 125, 376–382. [Google Scholar] [CrossRef]
- During, A.; Debouche, C.; Raas, T.; Larondelle, Y. Among plant lignans, pinoresinol has the strongest antiinflammatory properties in human intestinal caco-2 cells. J. Nutr. 2012, 142, 1798–1805. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Sun, L.-N.; Xin, H.-L.; Nian, H.; Song, H.-T.; Jiang, Y.-P.; Wei, Z.-Q.; Qin, L.-P.; Han, T. Anti-inflammatory constituents from the root of Litsea cubeba in lps-induced raw 264.7 macrophages. Pharm. Biol. 2016, 54, 1741–1747. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Cho, E.; Kwon, H.; Jeon, J.; Seong Sin, J.; Kwon Park, J.; Kim, J.-S.; Woong Choi, J.; Jin Park, S.; Jun, M.; et al. Akt and calcium-permeable ampa receptor are involved in the effect of pinoresinol on amyloid β-induced synaptic plasticity and memory deficits. Biochem. Pharmacol. 2021, 184, 114366. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Büchter, C.; Havermann, S.; Wätjen, W. The lignan pinoresinol induces nuclear translocation of daf-16 in Caenorhabditis elegans but has no effect on lifespan. Phytother. Res. 2015, 29, 894–901. [Google Scholar] [CrossRef]
- Saul, N.; Möller, S.; Cirulli, F.; Berry, A.; Luyten, W.; Fuellen, G. Health and longevity studies in C. elegans: The “healthy worm database” reveals strengths, weaknesses and gaps of test compound-based studies. Biogerontology 2021, 22, 215–236. [Google Scholar] [CrossRef]
- Piao, X.M.; Huo, Y.; Kang, J.P.; Mathiyalagan, R.; Zhang, H.; Yang, D.U.; Kim, M.; Yang, D.C.; Kang, S.C.; Wang, Y.P. Diversity of ginsenoside profiles produced by various processing technologies. Molecules 2020, 25, 4390. [Google Scholar] [CrossRef]
- Hou, M.; Wang, R.; Zhao, S.; Wang, Z. Ginsenosides in panax genus and their biosynthesis. Acta Pharm. Sin. B 2021, 11, 1813–1834. [Google Scholar] [CrossRef]
- Hasegawa, H.; Matsumiya, S.; Uchiyama, M.; Kurokawa, T.; Inouye, Y.; Kasai, R.; Ishibashi, S.; Yamasaki, K. Inhibitory effect of some triterpenoid saponins on glucose transport in tumor cells and its application to in vitro cytotoxic and antiviral activities. Planta Med. 1994, 60, 240–243. [Google Scholar] [CrossRef]
- Yoshizaki, K.; Devkota, H.P.; Fujino, H.; Yahara, S. Saponins composition of rhizomes, taproots, and lateral roots of satsuma-ninjin (Panax japonicus). Chem. Pharm. Bull. 2013, 61, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.-L.; Li, Y.-J.; Chen, J.; Li, P. Triterpenoid saponins in roots of Achyranthese bidentata. Chin. J. Nat. Med. 2012, 10, 98–101. [Google Scholar] [CrossRef]
- Chen, W.; Balan, P.; Popovich, D.G. Ginsenosides analysis of New Zealand–Grown forest Panax ginseng by lc-qtof-ms/ms. J. Ginseng Res. 2020, 44, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Sagaya Jansi, R.; Khusro, A.; Agastian, P.; Alfarhan, A.; Al-Dhabi, N.A.; Arasu, M.V.; Rajagopal, R.; Barcelo, D.; Al-Tamimi, A. Emerging paradigms of viral diseases and paramount role of natural resources as antiviral agents. Sci. Total Environ. 2021, 759, 143539. [Google Scholar] [CrossRef]
- Ren, T.; Zhang, H.; Wang, J.; Zhu, J.; Jin, M.; Wu, Y.; Guo, X.; Ji, L.; Huang, Q.; Yang, H.; et al. Mcu-dependent mitochondrial Ca2+ inhibits NAD+/SIRT3/SOD2 pathway to promote ros production and metastasis of hcc cells. Oncogene 2017, 36, 5897–5909. [Google Scholar] [CrossRef]
- Schulz, T.J.; Zarse, K.; Voigt, A.; Urban, N.; Birringer, M.; Ristow, M. Glucose restriction extends Caenorhabditis elegans life span by inducing mitochondrial respiration and increasing oxidative stress. Cell Metab. 2007, 6, 280–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pun, P.B.; Gruber, J.; Tang, S.Y.; Schaffer, S.; Ong, R.L.; Fong, S.; Ng, L.F.; Cheah, I.; Halliwell, B. Ageing in nematodes: Do antioxidants extend lifespan in Caenorhabditis elegans? Biogerontology 2010, 11, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.I.; Harrison, D.G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Jiang, L.; Zhang, H.; Shimoda, L.A.; DeBerardinis, R.J.; Semenza, G.L. Analysis of hypoxia-induced metabolic reprogramming. Methods Enzymol. 2014, 542, 425–455. [Google Scholar]
- Braeckman, B.P.; Smolders, A.; Back, P.; De Henau, S. In vivo detection of reactive oxygen species and redox status in Caenorhabditis elegans. Antioxid. Redox Signal. 2016, 25, 577–592. [Google Scholar] [CrossRef] [Green Version]
- Schlotterer, A.; Hamann, A.; Kukudov, G.; Ibrahim, Y.; Heckmann, B.; Bozorgmehr, F.; Pfeiffer, M.; Hutter, H.; Stern, D.; Du, X.; et al. Apurinic/apyrimidinic endonuclease 1, p53, and thioredoxin are linked in control of aging in C. elegans. Aging Cell 2010, 9, 420–432. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Apanasenko, I.E.; Kim, A.V.; Shelepova, E.A.; Khalikov, S.S.; Polyakov, N.E. Spectroscopic and molecular dynamics characterization of glycyrrhizin membrane-modifying activity. Colloids Surf. B Biointerfaces 2016, 147, 459–466. [Google Scholar] [CrossRef]
- Selyutina, O.Y.; Polyakov, N.E.; Korneev, D.V.; Zaitsev, B.N. Influence of glycyrrhizin on permeability and elasticity of cell membrane: Perspectives for drugs delivery. Drug Deliv. 2016, 23, 858–865. [Google Scholar] [CrossRef]
- Li, N.; Zhou, T.; Wu, F.; Wang, R.; Zhao, Q.; Zhang, J.Q.; Yang, B.C.; Ma, B.L. Pharmacokinetic mechanisms underlying the detoxification effect of glycyrrhizae radix et rhizoma (gancao): Drug metabolizing enzymes, transporters, and beyond. Expert Opin. Drug Metab. Toxicol. 2019, 15, 167–177. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mean Survival | Min. Survival [Days] | Med. Survival [Days] | Max. Survival [Days] | n | p-Value (Compared to Control) | |
|---|---|---|---|---|---|---|---|
| Days ± SE | % | ||||||
| Control | 2.41 ± 0.19 | 100 | 0.84 | 2.19 | 6 | 147 | |
| C. chinensis | 2.85 ± 0.17 | 118.3 | 1.17 | 3.71 | 6 | 124 | 0.0132 |
| E. ulmoides | 2.86 ± 0.17 | 118.7 | 1.21 | 3.43 | 6 | 108 | 0.0144 |
| Compound | m/z (Da) | RT (min) | C. chinensis | E. ulmoides | Fraction O |
|---|---|---|---|---|---|
| Peak Area (%) | Peak Area (%) | Peak Area (%) | |||
| Hydroxygallic acid derivative | 187.096 | 8.002 | 5.02 | 0.67 | 4.70 |
| Unidentified | 195.308 | 0.630 | 4.50 | 0.60 | 0.01 |
| 2-C-Methyl-D-erythritol 4-phosphate | 215.032 | 0.643 | 2.15 | 1.02 | 3.5 × 10−3 |
| Dienoic acid derivative | 311.222 | 14.082 | 2.10 | 2.1 × 10−3 | 7.0 × 10−3 |
| d-Pinoresinol-4-O-glucoside | 519.186 | 7.558 | 1.23 | 0.13 | 0.01 |
| Stearic acid or hexadecenoic acid | 311.201 | 14.806 | 1.15 | 0.12 | 0.03 |
| Cinnamic acid derivative | 265.147 | 14.665 | 1.14 | 0.17 | 0.07 |
| Coumarin derivative | 147.044 | 7.373 | 1.12 | 1.8 × 10−3 | 2.3 × 10−4 |
| Hexadecanoic acid | 315.253 | 13.870 | 1.07 | 0.09 | 0.35 |
| Disaccharide | 341.108 | 0.707 | 1.05 | 0.29 | 0.03 |
| Disaccharide | 377.085 | 0.708 | 1.05 | 0.46 | 4.2 × 10−3 |
| Disaccharide | 387.114 | 0.707 | 0.85 | 0.29 | 0.03 |
| Palmitic acid or hexadecanoate | 315.253 | 13.720 | 0.94 | 0.05 | 0.20 |
| Unidentified | 187.006 | 4.116 | 0.91 | 0.01 | 2.2 × 10−5 |
| 4-alpha-formyl-stigmasta-7,24(241)-dien-3-beta-ol | 455.352 | 15.514 | 0.83 | 0 | 4.7 × 10−3 |
| Oleic acid derivative | 339.232 | 15.544 | 1.00 | 2.62 | 4.0 × 10−4 |
| Zingibroside R1 | 793.437 | 12.132 | 0.07 | 0.02 | 0.02 |
| Genistein derivative | 431.228 | 13.926 | 0.04 | 2.0 × 10−4 | 3.2 × 10−4 |
| Chlorogenic acid | 353.087 | 4.978 | 0.03 | 0.12 | 3.0 × 10−3 |
| Compound | m/z (Da) | RT (min) | C. chinensis | E. ulmoides | Fraction O |
|---|---|---|---|---|---|
| Peak Area (%) | Peak Area (%) | Peak Area (%) | |||
| Apigenin | 269.045 | 10.484 | 5.5 × 10−4 | 0.03 | 3.3 × 10−3 |
| Astragalin | 447.092 | 8.269 | 5.1 × 10−5 | 3.8 × 10−4 | 2.4 × 10−4 |
| Caffeoylquinic acid derivative | 515.139 | 5.473 | 5.3 × 10−4 | 0.30 | 1.0 × 10−3 |
| Caffeoylquinic acid derivative | 515.140 | 4.958 | |||
| Caffeoylquinic acid derivative | 515.140 | 4.898 | |||
| Caffeoylquinic acid derivative | 515.136 | 5.402 | |||
| Caffeoylquinic acid derivative | 515.139 | 5.183 | |||
| Caffeoylquinic acid derivative | 515.139 | 4.602 | |||
| Caffeoylquinic acid derivative | 515.139 | 4.832 | |||
| Caffeoylquinic acid derivative | 515.140 | 4.339 | |||
| Caffeoylquinic acid derivative | 515.140 | 3.576 | |||
| Caffeoylquinic acid derivative | 515.140 | 4.029 | |||
| Caffeoylquinic acid derivative | 515.140 | 3.678 | |||
| Chlorogenic acid derivative | 353.087 | 5.584 | 0.02 | 0.08 | 9.1 × 10−4 |
| Chlorogenic acid derivative | 353.087 | 5.120 | |||
| Isorhamnetin-o-glucoside | 477.103 | 7.783 | 0 | 0.20 | 2.8 × 10−3 |
| Isorhamnetin-o-glucoside | 477.103 | 7.675 | |||
| Kaempferol 3-O-rhamnoside-7-O-glucoside | 593.148 | 5.787 | 3.9 × 10−3 | 0 | 5.2 × 10−5 |
| Kaempferol 3-O-rhamnoside-7-O-glucoside | 593.149 | 0.738 | |||
| Kaempferol-3-rhamnosyhexose | 593.129 | 9.449 | 0 | 0.12 | 2.3 × 10−3 |
| Kaempferol-glycosides derivative | 593.129 | 9.335 | 0 | 0.15 | 0.01 |
| Kaempferol-glycosides derivative | 755.182 | 7.402 | |||
| Kaempferol-glycosides derivative | 755.181 | 7.663 | |||
| Kaempferol-o-coumaroyglucoside-o-hexoside | 755.182 | 7.941 | 0 | 0.01 | 3.7 × 10−3 |
| Kaempferol-O-dihexoside | 609.145 | 5.505 | 0 | 0.07 | 3.7 × 10−3 |
| Lignan-coumaroylglucoside | 697.364 | 11.582 | 5.1 × 10−4 | 0.26 | 0.02 |
| Lignan-coumaroylglucoside derivative | 697.364 | 11.440 | |||
| Lignan-coumaroylglucoside derivative | 697.364 | 11.141 | |||
| Lignan-o-coumaroylglucoside derivative | 683.349 | 10.991 | |||
| Quercetin | 301.035 | 7.002 | 1.6 × 10−4 | 0.03 | 2.0 × 10−4 |
| Quercetin-apiosyl-galactose | 595.129 | 6.579 | 0 | 1.07 | 3.5 × 10−3 |
| Quercetin-glucosides derivative | 625.141 | 6.702 | 2.6 × 10−5 | 1.08 | 3.5 × 10−3 |
| Quercetin-glucosides derivative | 625.140 | 5.970 | |||
| Quercetin-glucosides derivative | 625.141 | 6.702 | |||
| Quercetin-glucosides derivative | 625.141 | 6.910 | |||
| Rutin | 609.145 | 6.814 | 3.1 × 10−4 | 0.01 | 1.5 × 10−4 |
| Secoisolariciresinol | 361.165 | 12.401 | 3.8 × 10−4 | 1.0 × 10−4 | 1.8 × 10−4 |
| Treatment | Mean Lifespan | Min. [Days] | Med. [Days] | Max. [Days] | n | p-Value (Compared to Control) | |
|---|---|---|---|---|---|---|---|
| Days ± SE | % | ||||||
| Control | 19.69 ± 0.42 | 100.0 | 16.0 | 18.8 | 28.0 | 91 | |
| C. chinensis | 21.74 ± 0.47 | 110.4 | 17.4 | 20.8 | 31.0 | 98 | 0.001 |
| Astragalin | 21.28 ± 0.43 | 108.1 | 17.4 | 20.2 | 30.0 | 98 | 0.003 |
| Pinoresinol | 19.23 ± 0.40 | 97.7 | 15.9 | 18.3 | 28.0 | 93 | 1.000 |
| Zingibroside R1 | 21.66 ± 0.46 | 110.0 | 17.5 | 21.0 | 30.0 | 94 | 0.041 |
| Treatment | Mean Lifespan | Min. [Days] | Med. [Days] | Max. [Days] | n | p-Value (Compared to Control) | |
|---|---|---|---|---|---|---|---|
| Days ± SE | % | ||||||
| Control | 3.39 ± 0.10 | 100.0 | 1.99 | 2.86 | 7.00 | 179 | |
| C. chinensis | 4.44 ± 0.11 | 131.0 | 2.84 | 3.71 | 9.00 | 212 | <0.0001 |
| Astragalin | 4.04 ± 0.09 | 119.2 | 2.59 | 3.43 | 8.00 | 229 | <0.0001 |
| Pinoresinol | 3.82 ± 0.09 | 112.7 | 2.32 | 3.22 | 8.00 | 233 | 0.0098 |
| Zingibroside R1 | 3.80 ± 0.08 | 112.1 | 2.42 | 3.23 | 8.00 | 261 | 0.0137 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, S.M.A.; Alseekh, S.; Siems, K.; Fernie, A.R.; Luyten, W.; Schmitz-Linneweber, C.; Saul, N. Identification of a Hydroxygallic Acid Derivative, Zingibroside R1 and a Sterol Lipid as Potential Active Ingredients of Cuscuta chinensis Extract That Has Neuroprotective and Antioxidant Effects in Aged Caenorhabditis elegans. Nutrients 2022, 14, 4199. https://doi.org/10.3390/nu14194199
Sayed SMA, Alseekh S, Siems K, Fernie AR, Luyten W, Schmitz-Linneweber C, Saul N. Identification of a Hydroxygallic Acid Derivative, Zingibroside R1 and a Sterol Lipid as Potential Active Ingredients of Cuscuta chinensis Extract That Has Neuroprotective and Antioxidant Effects in Aged Caenorhabditis elegans. Nutrients. 2022; 14(19):4199. https://doi.org/10.3390/nu14194199
Chicago/Turabian StyleSayed, Shimaa M. A., Saleh Alseekh, Karsten Siems, Alisdair R. Fernie, Walter Luyten, Christian Schmitz-Linneweber, and Nadine Saul. 2022. "Identification of a Hydroxygallic Acid Derivative, Zingibroside R1 and a Sterol Lipid as Potential Active Ingredients of Cuscuta chinensis Extract That Has Neuroprotective and Antioxidant Effects in Aged Caenorhabditis elegans" Nutrients 14, no. 19: 4199. https://doi.org/10.3390/nu14194199