1. Introduction
Congenital heart disease (CHD) is typically defined as a gross structural abnormality of the heart and/or great vessels that is present at birth [
1,
2]. It has been reported that the birth prevalence of CHD has increased significantly since the 1930s and reached a maximum of over 9 per 1000 live births since 1995 [
1,
3]. Ventricular septal defect (VSD) has been recognized as the most common congenital cardiac malformation and accounts for roughly 30–40% of all cardiac anomalies [
1,
4]. Over the past decades, considerable inherited causes and noninherited modifiable factors have been implicated in the development of CHD and its subgroups [
5,
6,
7,
8]. Recently, there has been a consensus that genetic factors and environmental factors interact in the etiology of most nonsyndromal forms of CHD [
9,
10], naturally including VSD.
A recent review showed strong evidence that oral prenatal fortification and supplementation dosing of folic acid (FA) can prevent the incidence of VSD and atrial septal defect (ASD) [
11]. Women with a diverse diet during pregnancy (dietary diversity score, DDS ≥5) had lower risks of having fetuses with total CHD and VSD [
12]. Furthermore, the dietary intake of vitamins and minerals was found to be associated with a reduced risk of CHD in offspring, including B-vitamin, vitamin D, zinc, and selenium [
13,
14,
15]. Since different nutrients interact with one another in many metabolic pathways, it seems that the association would not remain constant when various nutrients coexist in the same food. In addition, the dietary pattern differs a lot owing to the discrepancy in economics, geographical environment, social culture, race, and so on. Therefore, the first concern we would care to discuss is the association between maternal dietary habits and VSD in offspring.
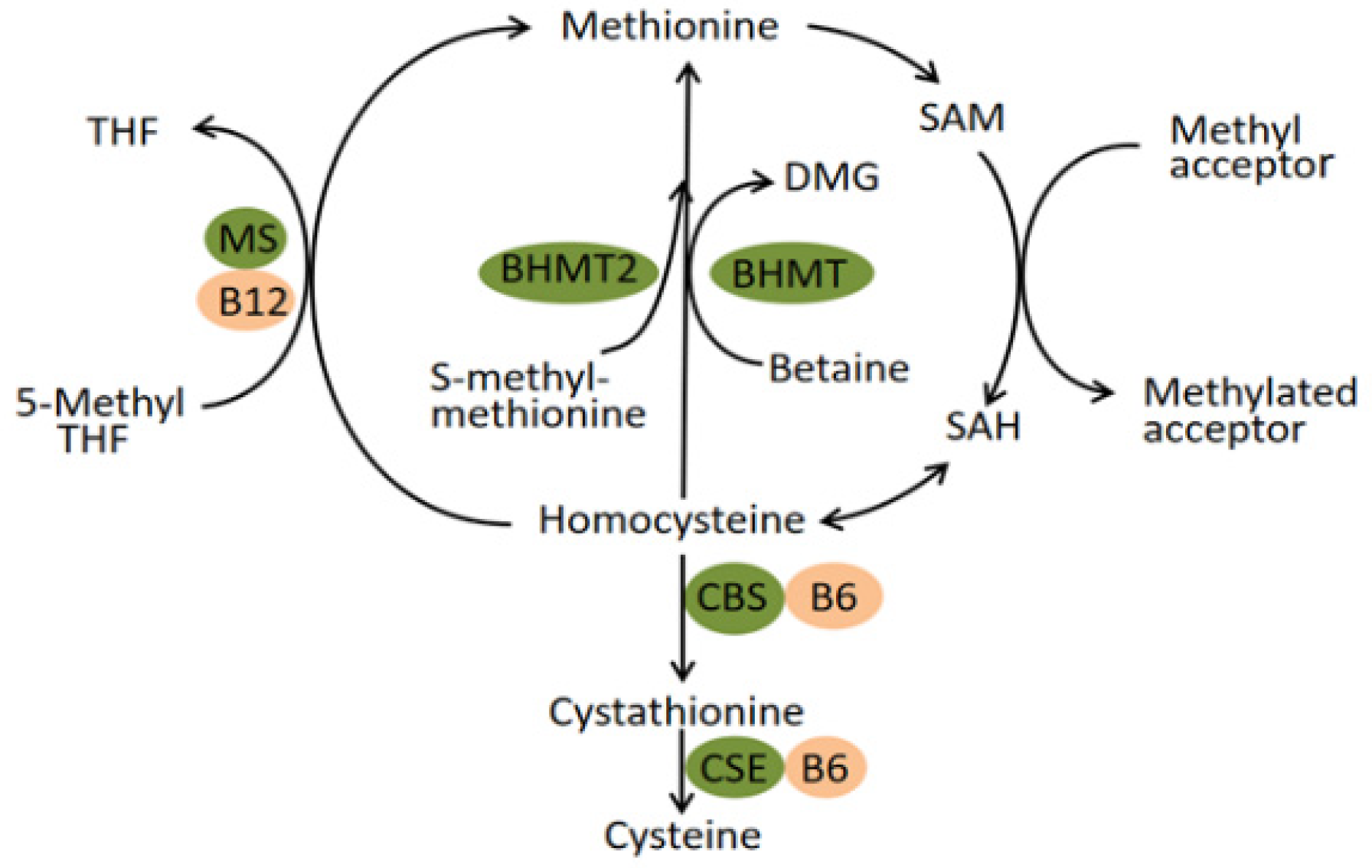
The human betaine-homocysteine methyltransferase (BHMT) gene maps to 5q13.1–q15, spans about 20 kilobases of DNA and contains eight exons and seven introns [
16,
17]. The enzyme it encodes, betaine-homocysteine methyltransferase, catalyzes the transfer of a methyl group from betaine to homocysteine (Hcy), forming dimethylglycine and methionine. Generally, the homeostasis of plasma homocysteine benefits from the transulfuration pathway involving cystathionine β synthase (CBS) and the remethylation pathway involving BHMT, BHMT2, and methionine synthase (MS) (
Figure 1) [
18]. In the latter pathway, the catalytic activity of BHMT2 is absolutely diet-dependent since its substrate, S-methylmethionine, can only be biosynthesized by various plants mainly belonging to the Brassicaceae family rather than mammals [
19,
20]. Experimental research conducted in mice suggested that BHMT is a predominant enzyme for the elimination of Hcy while the MS has little excess capacity to methylate the Hcy [
18]. Therefore, the remethylation reaction catalyzed by BHMT seems to play a vital role in preventing the toxic accumulation of Hcy. In fact, BHMT catalyzes up to 50% of homocysteine metabolism in the human liver, where the enzyme is highly expressed [
21,
22]. The latest literature revealed that elevated Hcy concentrations acted as a risk factor for multiple congenital anomalies in human production, mainly comprising neural tube defects (NTD), orofacial clefts, and CHD [
23,
24,
25]. The discovery has been generally accepted that the 677 C→T mutation in the methylenetetrahydrofolate reductase (MTHFR) gene contributed to elevated tHcy and is a genetic risk factor for diseases associated with hyperhomocysteinaemia [
26]. Moreover, this mutation has been applied to antenatal screening for pregnant women in China. The thought naturally emerged that polymorphisms of the BHMT gene exist that reduce BHMT activity and increase plasma Hcy levels and thus increase malformation risk. In fact, research has been dedicated to exploring the association between BHMT gene polymorphisms and CHD, but with fixed results and little involving subgroups of CHD [
27,
28,
29,
30]. In this study, we focused on the largest subcategory of CHD, namely, VSD, to detect its association with polymorphisms of the maternal BHMT gene.
In addition, betaine, the substrate of BHMT, can be either obtained from food resources or produced from choline endogenously [
31]. Likewise, choline can also be produced endogenously via the hepatic phosphatidylethanolamine N-methyltransferase (PEMT) pathway. However, most people must consume this nutrient exogenously to prevent deficiency [
32]. Therefore, the BHMT activity, to a certain degree, is diet dependent. Animal studies did observe that pane of nutrition or the supply of some nutrients, including choline and methionine, can alter BHMT activity [
33,
34,
35]. In addition, it has been reported that women with a high intake of one-carbon cofactors had a lower risk of congenital anomalies in offspring, such as the neural tube defect (NTD) and perimembranous ventricular septal defect (VSD
pm) [
36,
37]. Overall, these valuable clues were collected to put forward a reasonable hypothesis that BHMT gene polymorphisms may interact with maternal dietary habits on congenital anomalies.
In this study, we determined VSD, the most common subgroup in CHD, as the interested outcome variable, which is relatively more sensitive to maternal nutrient intake. A hospital-based case-control study was carried out in an attempt to learn the following questions: a. the association of maternal dietary habits with risk of VSD in offspring; b. the association of polymorphisms of maternal BHMT gene with risk of VSD in offspring; c. the interaction between BHMT genetic variants and maternal dietary habits on VSD.
2. Materials and Methods
2.1. Design and Participants
This is a hospital-based case-control study that started in February 2018 and was over in March 2020. The cases and controls came from different departments in the same hospital, Hunan children’s hospital, which is famous partly for its sophisticated diagnosis and treatment techniques for CHD within the province. Considering the characteristics of the relatively low incidence of VSD compared with other chronic diseases, a convenient sampling method was used in the recruitment of the cases. VSD children, verified by both doppler echocardiography and surgery, were consecutively recruited from the Department of Cardiothoracic Surgery. Children in the control, free of any congenital malformations, were randomly selected from the Department of Child Healthcare. It is worth noting that cases only included VSD children that may or may not be diagnosed with other congenital heart diseases; those coexisting with any other extra-cardiac malformations were excluded from the study. Additionally, informed consent was obtained from all of the participants, and the possible consequences of the study were explained. The exclusion criterions mainly included: minority mothers, mothers conceiving children through in vitro fertilization or other conception methods, adoptive mothers or stepmothers, and mothers suffering from mental disorders or any other physical diseases so that this did not hinder the provision of accurate exposure information and blood samples. Finally, a total of 426 mothers of VSD children and 740 control mothers were included in the study.
The protocol of this study was in accordance with the guidelines of the 1964 Helsinki Declaration, and the Ethics Committee of Xiangya School of Public Health, Central South University, officially approved this study in January 2018. (no. XYGW-2018-36).
2.2. Information Collection
The outcome we focused on in the study was VSD in offspring, which was diagnosed by professional physicians via both doppler echocardiography and surgery. The interested exposures were maternal dietary habits in early pregnancy, which were collected from a self-designed food frequency questionnaire. We consulted The Dietary Guidelines for Chinese Residents and went deep into the local food culture to develop the questionnaire. Eleven main categories were determined, involving smoked foods, barbecued foods, fried foods, pickled vegetables, fresh vegetables, fresh fruits, fresh meat, fish and shrimp, fresh eggs, beans, and milk products. Each category was provided with three choices: a. hardly (less than or equal to two times per week); b. sometimes (three to five times per week); c. often (more than or equal to six times per week). The questionnaire was pre-investigated using eligible mothers (test–retest reliability: r = 0.826; internal consistency: α = 0.769).
In addition, we also collected various pieces of maternal information that might influence the outcomes of their offspring, mainly including the child-bearing age (<35 years or ≥35 years), pre-pregnancy BMI (calculated with their pre-pregnancy height and weight, <18.5, 18.5–23.9, 24–26.9, or ≥27), education level (less than primary or primary, junior high school, high school or technical secondary school, college or above), consanguineous marriages (yes or no), gestational diabetes mellitus (yes or no), gestational hypertension (yes or no), abnormal pregnancy history before this pregnancy (yes or no), congenital malformations in family members (yes or no), exposure of environmental pollutants (yes or no), antibiotic use in early pregnancy (yes or no), tobacco exposure in early pregnancy (yes or no), alcohol exposure in early pregnancy (yes or no), and periconceptional folate use (yes or no).
An epidemiological survey was conducted by well-trained investigators when participants were waiting for their operation arrangements in the wards or medical check-ups in the Department of Child Health. In China, every expectant mother has a personal Maternal and Child Health Manual, which provides their sociodemographic information, the results of regular medical check-ups, and necessary exposure information. So, in the course of the investigation, we consulted the participants’ manual to further confirm the abovementioned information obtained from face-to-face interviews, which enabled us to reduce recall bias to a certain extent.
2.3. Sample Collection and Genotyping
Five milliliters of peripheral venous blood were collected from every single participant after the face-to-face interview. All of the obtained blood samples would be brought back to the laboratory at low temperatures (≤4 °C) within twelve hours and then divided into two layers using a high-speed centrifuge: the blood cell layer and the plasma layer. Both were stored in an ultra-low-temperature freezer until genotyping. The DNA was extracted from the blood cell samples with the QIAamp DNA Blood Mini Kit (Qiagen, Valencia, CA, USA). Genotyping was performed by matrix-assisted laser desorption and ionization time-of-flight mass spectrometry MassARRAY system (Agena iPLEX assay, San Diego, CA, USA). The laboratory technicians who performed SNP detection and recorded the genotype data were blind to whether each sample was from the cases or controls, thereby reducing selection bias to some extent.
Before genotyping, we consulted the NCBI and HapMap databases to determine the major SNP sites of the BHMT gene and simultaneously excluded the SNPs whose minor allele frequencies (MAF) were less than 10%. Furthermore, we imposed a minimum SNP genotyping call rate at the level of 50%, which was applied to ensure the data integrity of the individual’s genotypes. Moreover, the success rates for the SNPlex assays were >94% for 2 SNPs, from 90 to 94% for 2 SNPs. Finally, these genetic loci (rs3733890, rs1316753, rs567754, and rs1915706) were selected as candidate loci for this study.
2.4. Statistical Analysis
The data for the qualitative variables were expressed as absolute numbers (percentages). The chi-square test was used to assess the differences in qualitative variables across groups. The Hardy–Weinberg equilibrium (HWE) test was used to compare the differences in genotype distribution frequency in the control group (significance level at p < 0.01). We utilized a logistic regression model to detect whether the association between maternal dietary habits in early pregnancy, BHMT gene polymorphisms, and VSD in offspring existed and the level of the association. Both univariate and multivariate logistic regressions were adopted; the crude odds ratio (cOR) and its 95% confidential intervals (CI) were calculated by the former one without any adjustment; the adjusted odds ratio (aOR) and its 95% confidential intervals (CI) were calculated by the latter one, which adjusted for the significant confounders found using the chi-square test. For the significant SNPs and maternal dietary habits, these originally ternary variables were converted into binary variables. We then introduced all of the potential confounders, genetic factors, environmental factors, and their multiplicative interaction term into the same logistic regression model to determine the presence or absence of gene–environment interaction and assess its significance. When it comes to multiple hypothesis testing, the false discovery rate (FDR) based on the Benjamini–Hochberg method was used to correct for bias. A false discovery rate P value (FDR_P) of <0.1 was considered to be statistically significant. The calculation of FDR_P was completed using R software (version 4.1.3, stats package). Basic analyses were performed using SPSS 26.0 software (SPSS Inc., Chicago, IL, USA). The statistically significant results were those with the two-sided p-value < 0.05, except where otherwise specified.
4. Discussion
Research on the causes of CHD has made great strides, and more than 400 genes and important environmental factors have been determined to have substantial evidence in relation to the risk of developing CHD and its subgroups [
5,
6,
7,
8]. However, the conclusion can be easily drawn from the other hand that a single genetic or environmental factor may impose minimal effects on CHD. Moreover, the interaction of the two factors cannot be overlooked in the occurrence and development of CHD and its subgroups. In this study, we attempted to achieve an insight into the etiology of VSD, the most common subtype of CHD. The main purpose was to decide whether the association and interaction effect of BHMT gene polymorphisms and maternal dietary habits with VSD exists, which is conducive to the achievement of molecular genetic diagnostics and provide diet instruction to expectant mothers in early pregnancy.
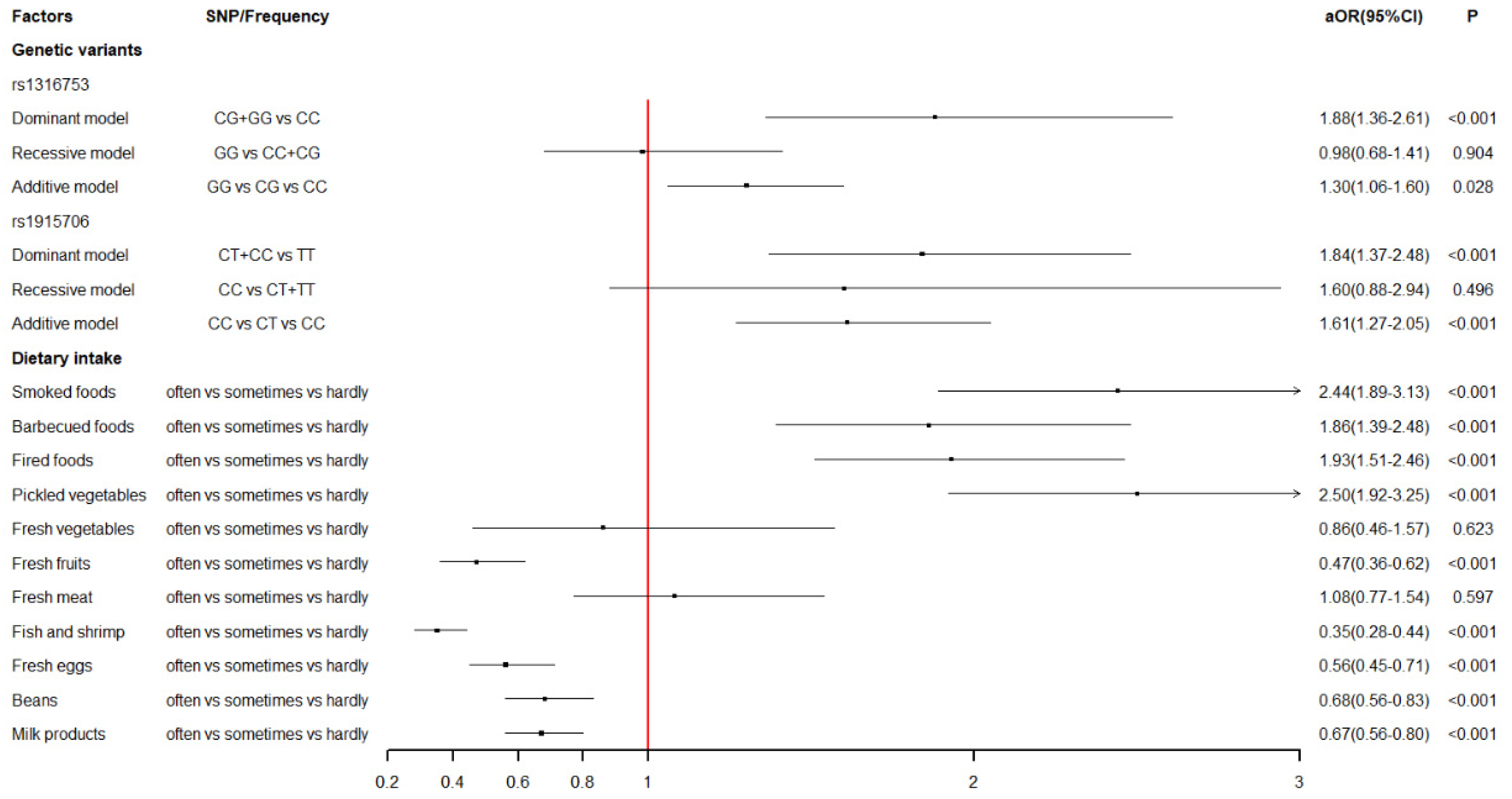
Firstly, we explored the association between maternal dietary intake during early pregnancy and the risk of VSD in offspring. The results sent two messages. On the one hand, mothers who reported excessive intake of smoked foods (aOR = 2.44), barbecued foods (aOR = 1.86), fried foods (aOR = 1.93), and pickled vegetables (aOR = 2.50) were more likely to have a VSD-affected child. On the other hand, mothers with an excessive intake of fresh fruits (aOR = 0.47), fish and shrimp (aOR = 0.35), fresh eggs (aOR = 0.56), beans (aOR = 0.68), and milk products (aOR = 0.67) were less likely to have a VSD-affected child. Generally, various harmful chemicals can be generated from improper food processing, and most of them are teratogens and carcinogens, such as nitrites, acrylamide (ACR), polycyclic aromatic hydrocarbons (PAH), and so on. Pickled vegetables have a wide range of nitrite and nitrate contents. Gravidas, who have an excessive intake of pickles, may suffer hypoxemia because increased nitrite can react with hemoglobin, rendering it incapable of carrying oxygen [
38]. A recent experimental study established a rodent model and reported that hypoxia was able to cause numerous abnormalities in cardiomyocyte gene expression, the electrophysiologic substrate of the heart, and contractile function, thus delaying cardiac maturation [
39]. ACR, identified in food in 2002, is mainly formed via the Maillard reaction, whereby a carbonyl compound reacts with the amino group of asparagine processed at high temperatures (>120 °C, such as barbecuing, frying, and baking) [
40,
41]. Although no direct evidence manifested its relation to heart defects, a number of animal studies have shown strong neurotoxic, genotoxic, carcinogenic, mutagenic, and teratogenic effects [
42]. Food is readily contaminated by PAH during the smoking process involving the combustion of fuel. A recent study reported that greater maternal levels of PAH exposure during pregnancy might be associated with an elevated prevalence of fetal CHD [
43]. Moreover, prior experimental research provided strong evidence that PAH exposure can result in abnormal heart looping, an enlarged ventricle with a thinner ventricular wall, and even developmental cardiac defects [
44,
45]. The protective foods detected in the study, such as fruits, fish and shrimp, eggs, beans, and milk products, are common foods on tables. Furthermore, they are packed with numerous nutrients, such as high-quality protein, vitamins, minerals, and so on, which have been extensively accepted to play a vital role in maintaining the health of gravidas and fetuses.
Moreover, we determined the association between maternal BHMT gene polymorphisms and the risk of VSD in offspring. Four SNPs (rs3733890, rs1316753, rs567754, and rs1915706) were considered in this study, and two SNPs (rs1316753 and rs1915706) were for the first time found to have a statistically significant association with VSD. To date, the BHMT gene remained relatively little studied compared with other folate- and homocysteine-metabolizing genes. The results in our study were only partly in accordance with prior studies. The rs3733890 polymorphism is a well-studied exon of the BHMT gene and undergoes a G-to-A change at nucleotide 716, leading to an arginine-to-glutamine substitution at amino acid 239. Its association with congenital defects has been widely explored but with contradictory results [
27,
46,
47,
48]. In the present study, we did not detect its significance in the occurrence of VSD. Rs567754 is an intronic variant of the BHMT gene, and neither previous data nor our data revealed an association with CHD or VSD in offspring [
27]. The interesting thing is that two other SNPs, rs1316753 and rs1915706, were observed to be statistically associated with a remarkably-increased risk of VSD in offspring. To the best of our knowledge, experimental or epidemiological research involving these two polymorphisms remains a nearly unworked area, meaning that their potential functional effects on BHMT are largely unknown. Qiping Feng et al. performed a genotype–phenotype correlation analysis on betaine-homocysteine methyltransferase and found that three introns (rs41272270, rs6875201, and rs7700790) and an intergenic variant (rs16876512) were significantly correlated with both BHMT activity and protein levels [
22]. Although this convincing research did not cover the two significant SNPs in our study, the analogy seems to be reasonable that the two intergenic variants, rs1316753 and rs1915706, are also capable of producing potential effects on the expression of the BHMT gene and subsequently influencing plasma hcy concentrations. The correlation between maternal hyperhomocysteinemia and CHD has been extensively studied and reviewed [
25,
49,
50]. Meanwhile, hcy-induced CHDs, such as the transposition of the great arteries (TGA), single ventricle defects (SVD), and VSD, have been found in embryos of different species (mice and chicken) [
51]. Therefore, the statistical association between maternal BHMT polymorphisms and VSD in offspring might be explained by the pathway from BHMT activity to elevated hcy levels to multiple congenital anomalies. Nonetheless, more related research is encouraged to provide clearer evidence, thus elucidating the potential mechanism.
Lastly, we also analyzed the gene–environment interaction and observed a significant interaction between genetic variants of the BHMT gene at rs1915706 and maternal bean intake on the risk of VSD in offspring. The expectant mothers who had the variant genotypes (CT + CC) and meanwhile reported a small intake of beans were at a significantly higher risk of VSD in offspring (aOR = 4.00) compared with those with the wild genotype (TT) and reported having a regular intake of beans in early pregnancy. Beans are an excellent source of zinc, choline, and multiple B vitamins (such as folate, thiamin, niacin, riboflavin, and pyridoxine) [
52,
53]. Notably, BHMT is a zinc-dependent cytosolic enzyme, and its substrate, betaine, is partly derived from dietary choline [
31,
54]. In addition, a stronger risk reduction in CHD has been found in the maternal folate + B-vitamin supplementation group compared with the maternal folate supplementation group, both setting the non-users as the reference group [
14]. Concerning whether a single nutrient can exist in various foods, we speculated that the deficiency of diverse nutrients coexisting in beans coincided with a genetic variant that contributes to the occurrence of VSD. This speculation seemed plausible since similar research had been conducted not long ago. Hartmut Cuny et al. demonstrated that when dietary undersupply during pregnancy was combined with a maternal heterozygous variant in Haao, a gene of the nicotinamide adenine dinucleotide (NAD) synthesis pathway, the incidence of embryo loss and malformations was significantly higher [
55]. This is a classical experimental study forcibly indicating a gene-diet interaction. As Gibson G and Berger K commented, the discovery of such interaction suggests that the close monitoring of nutrition in at-risk carrier mothers would be the type of personalized and predictive intervention that advocates of genomic health call for [
56]. Regardless, what we found in our study necessitates more convincing experimental research and crowd investigation to confirm it repeatedly.
Furthermore, several limitations in our study should be acknowledged. Firstly, although we perfected the study design and executed it strictly during the whole process as far as possible, the association found in this study, an observational case-control study, was relatively limited compared to a cohort study or an experimental study. So, in other words, well-designed prospective cohorts or reasonable experimental research are needed to validate our findings further. Secondly, the information on food frequency and related exposure in the questionnaire were obtained through retrospective investigation; we cannot rule out the possible limitation of recall bias. Thirdly, this is a hospital-based case-control study, and all of the cases came from the same department in a hospital; though its representativeness in the province for sophisticated diagnosis and treatment techniques, the selection bias still cannot be ignored. Fourthly, several potential confounders were determined and adjusted in the study, but there undoubtedly are other confounding covariates that might also influence the outcomes, such as common genetic polymorphisms and some nutritional biomarkers. The findings would be more convincing if taking these factors into consideration. Last but not least, maternal hcy concentration was not available in our research, which excludes the possibility of verifying the potential explanation that genetic variants of the BHMT gene may cause VSD by elevating maternal hcy levels.


 ,
, 



{kind=link}
{kind=link}
{kind=link}