
The Neuroprotective Effects of Spray-Dried Porcine Plasma Supplementation Involve the Microbiota−Gut−Brain Axis

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Diets
2.2. Sample Collection
2.3. Open Field Test
2.4. Novel Object Recognition Test
2.5. Western Blot
2.6. Real-Time PCR
2.7. Quantification of Cytokines
2.8. Lipopolysaccharide Determination
2.9. Extraction and Purification of Total Genomic DNA
2.10. 16S rDNA Gene Analysis
2.11. Statistical Analysis
3. Results
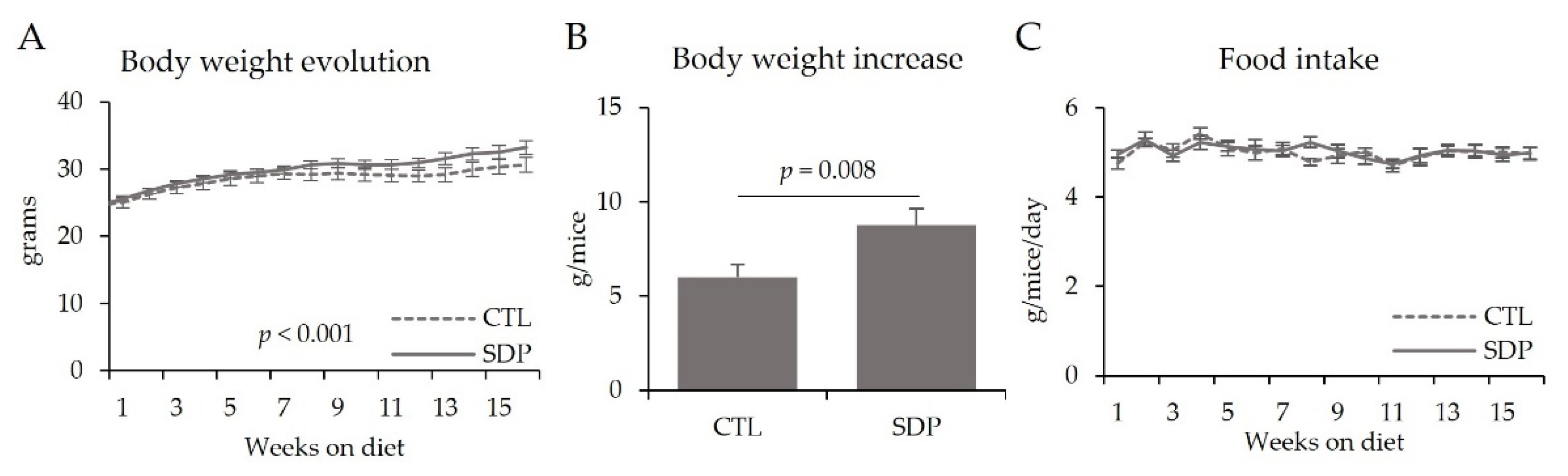
3.1. Body Weight and Food Intake
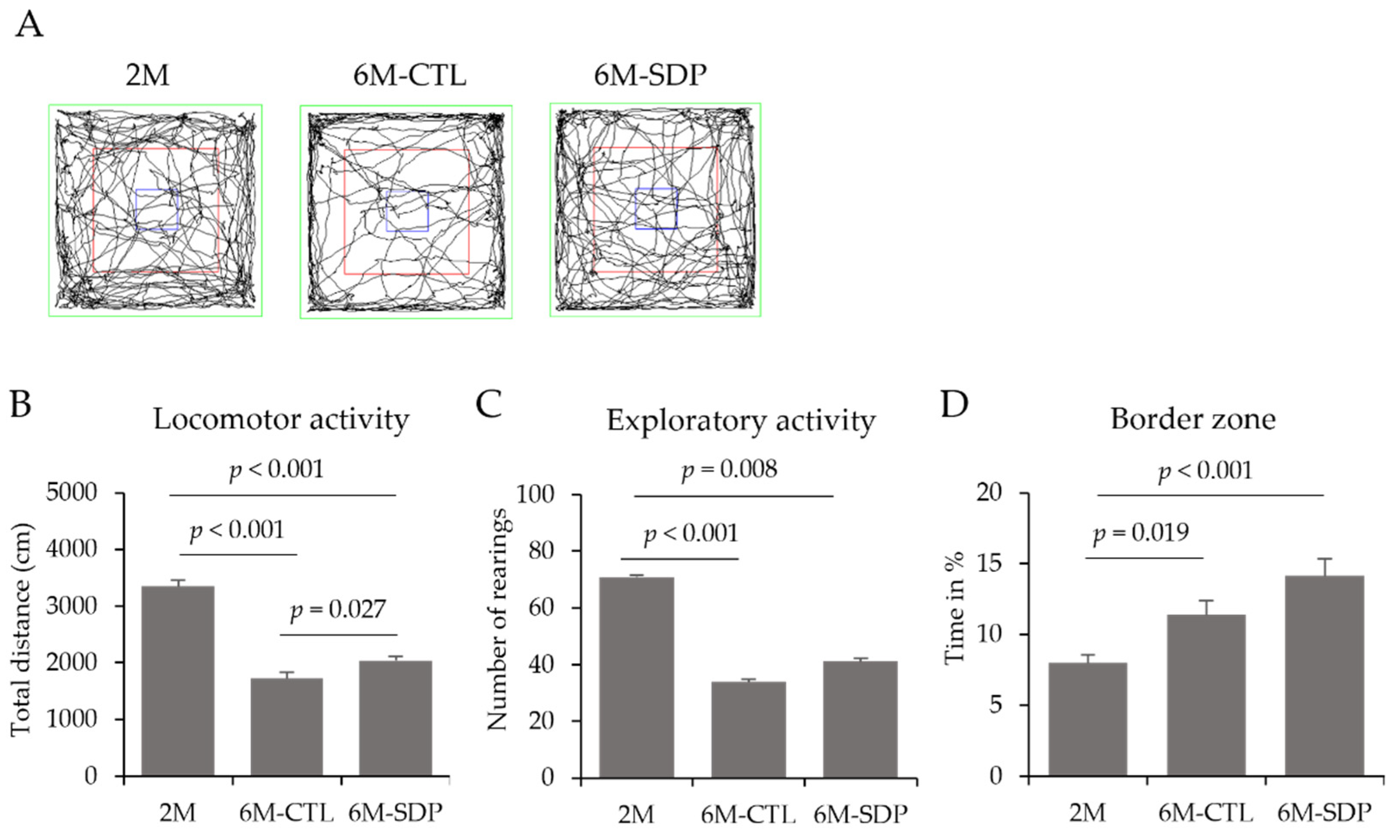
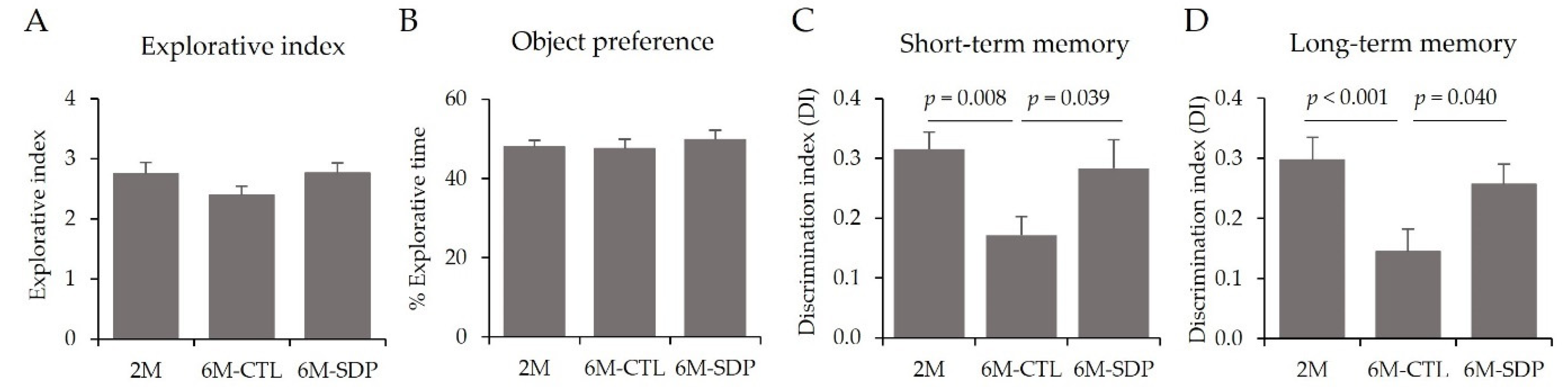
3.2. Behavioral and Cognitive Tests
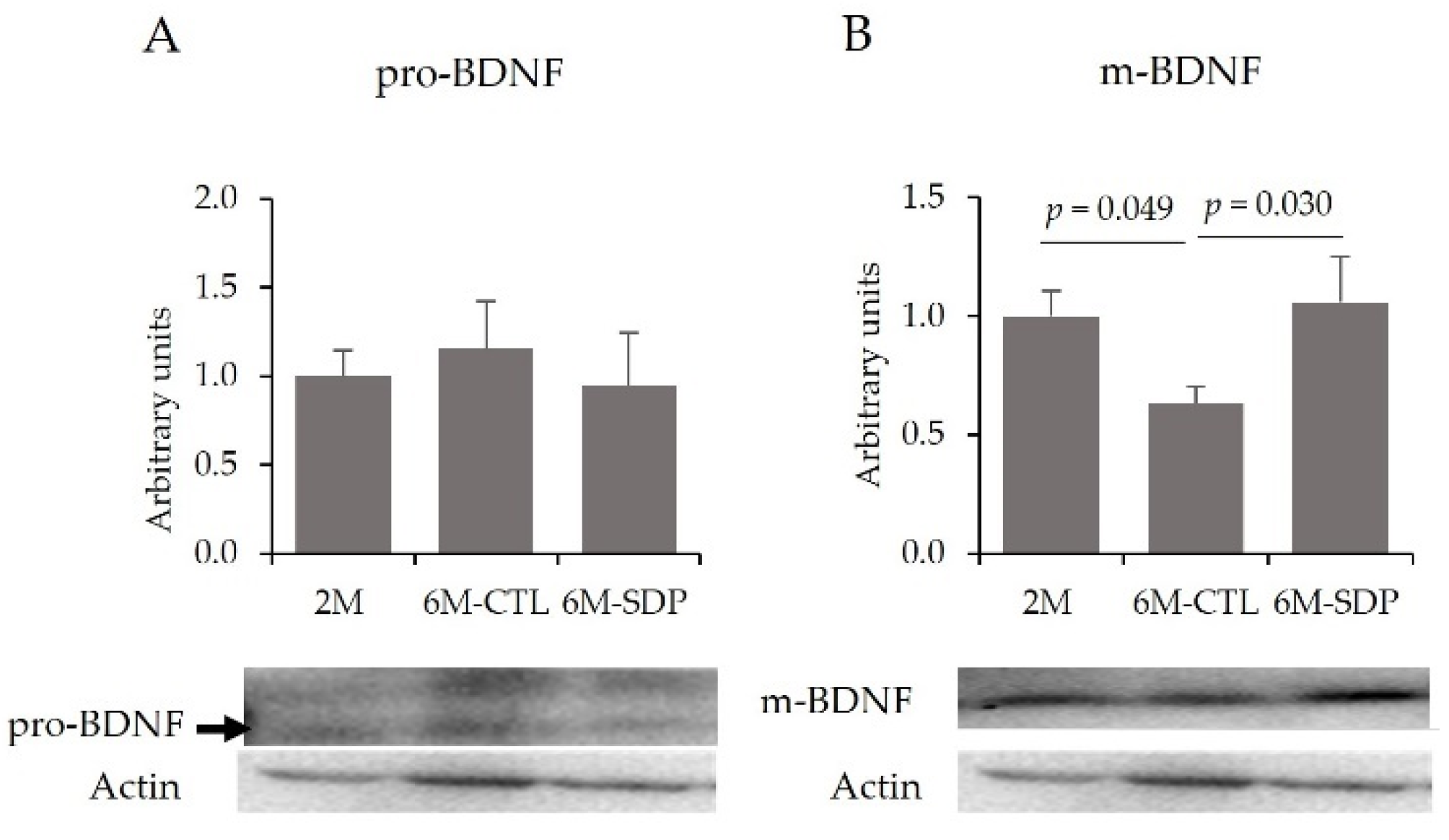
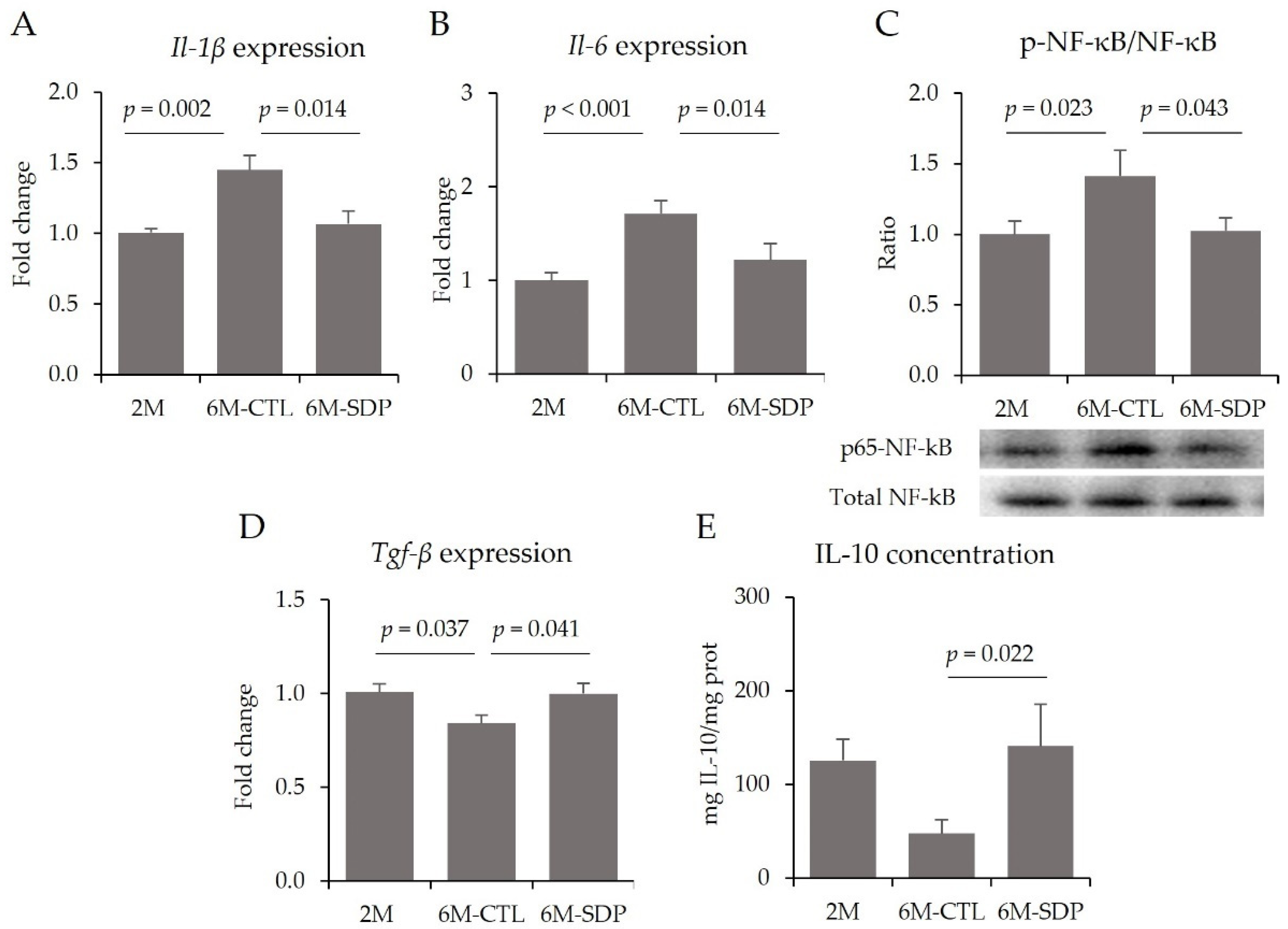
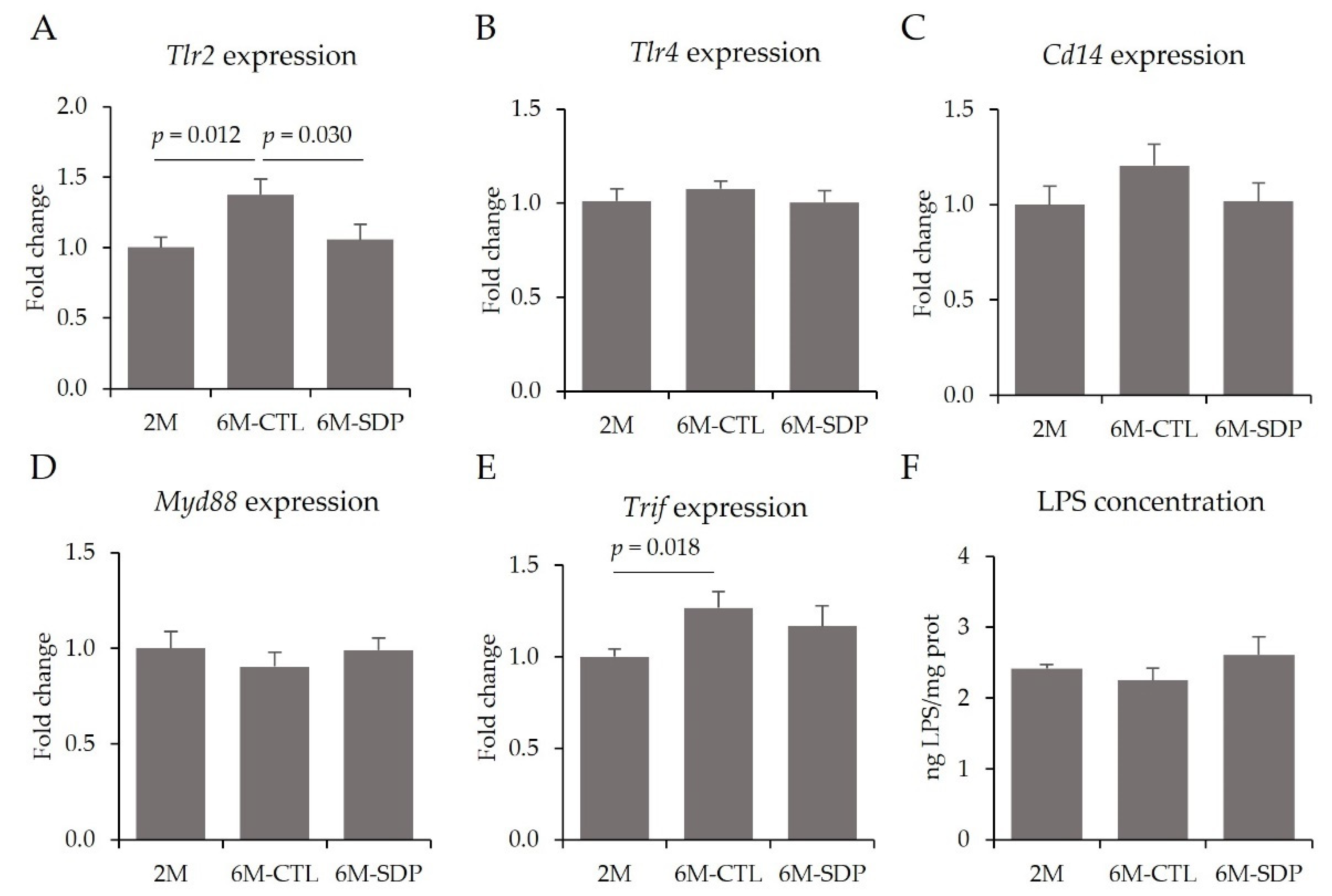
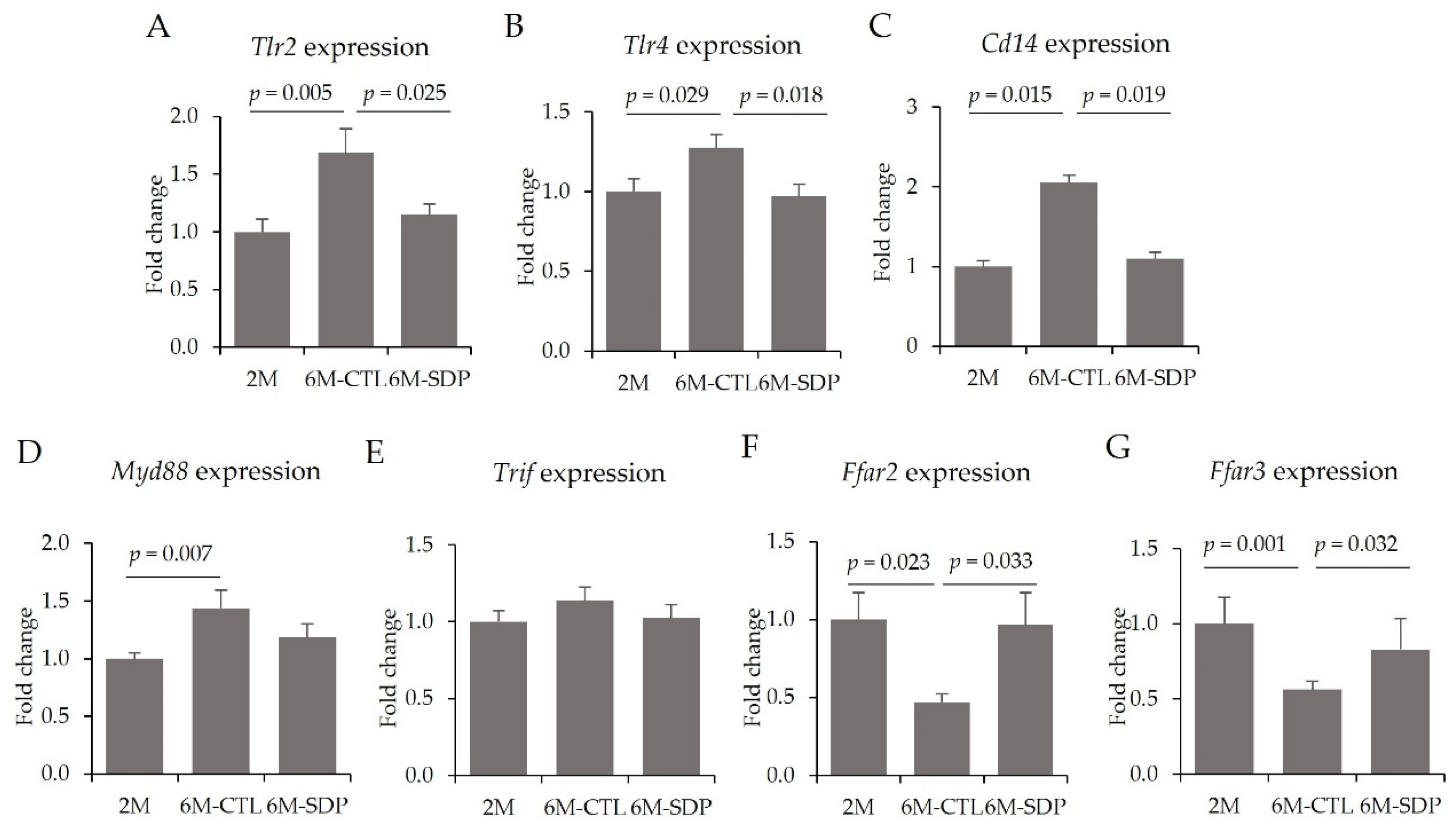
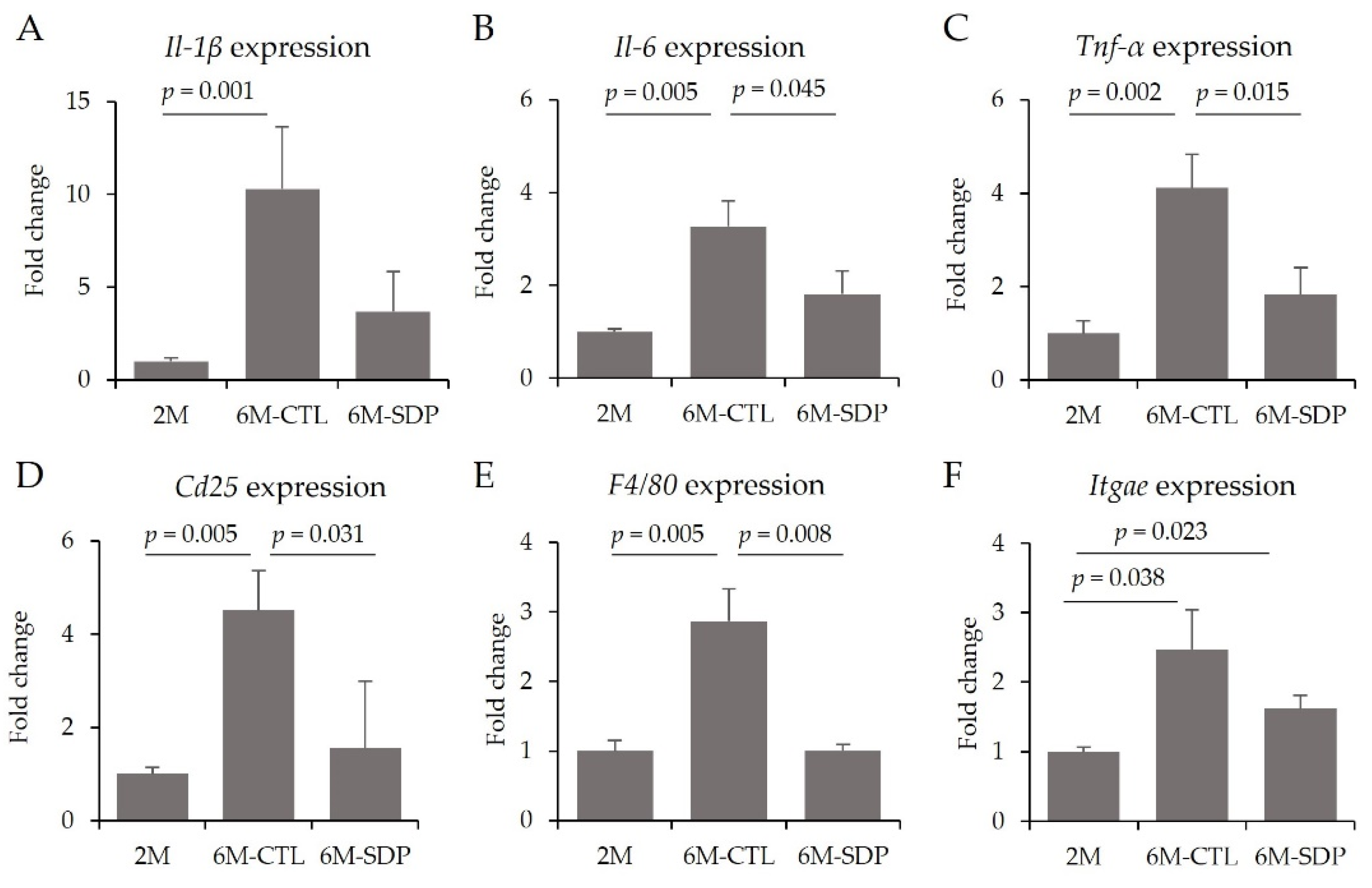
3.3. Effects on the CNS
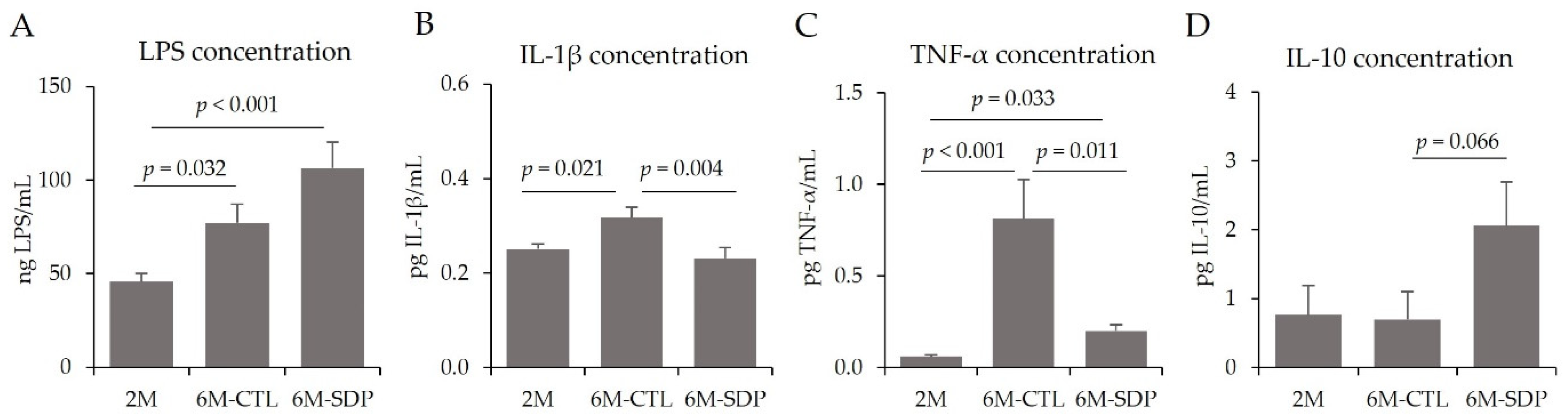
3.4. Systemic Effects
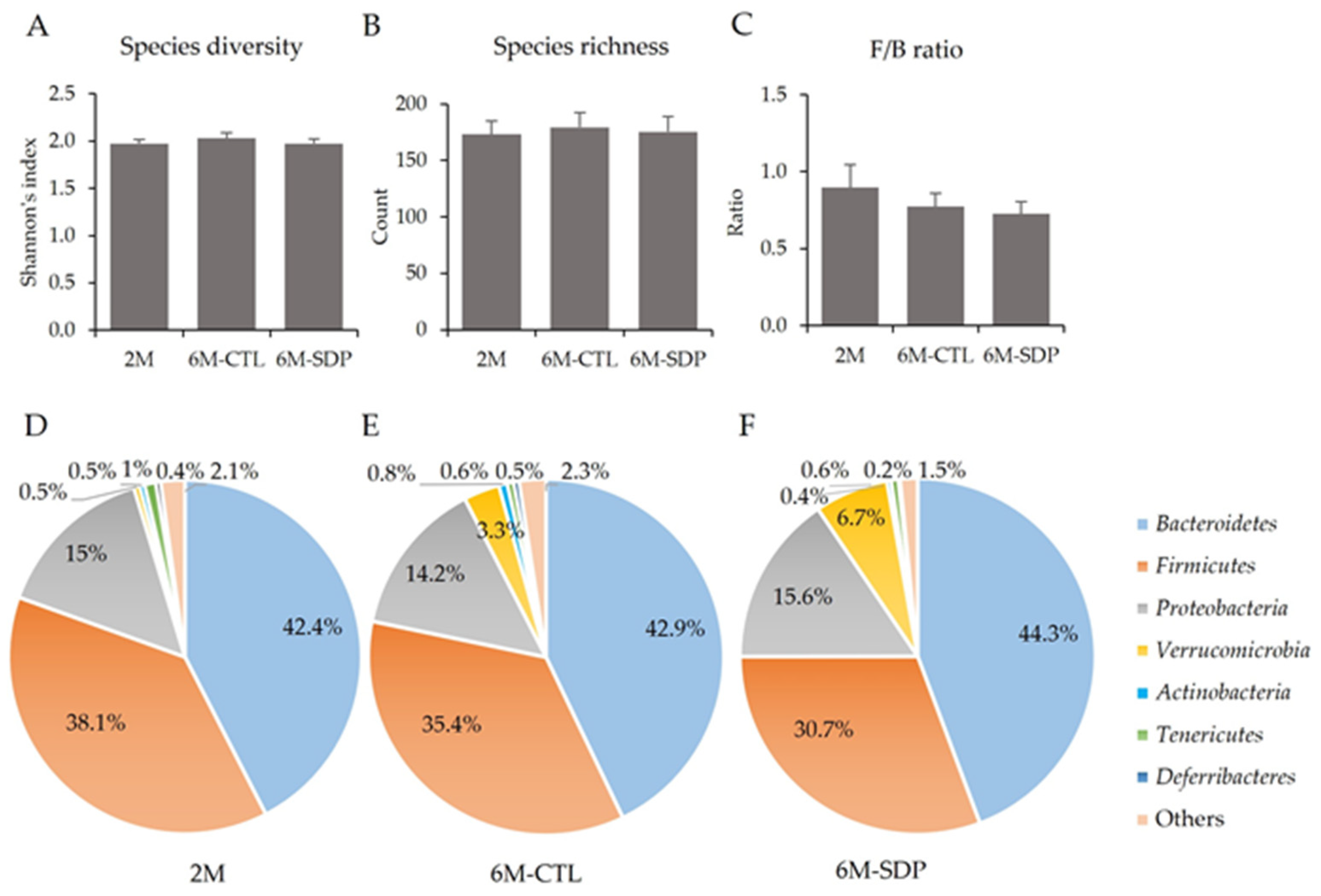
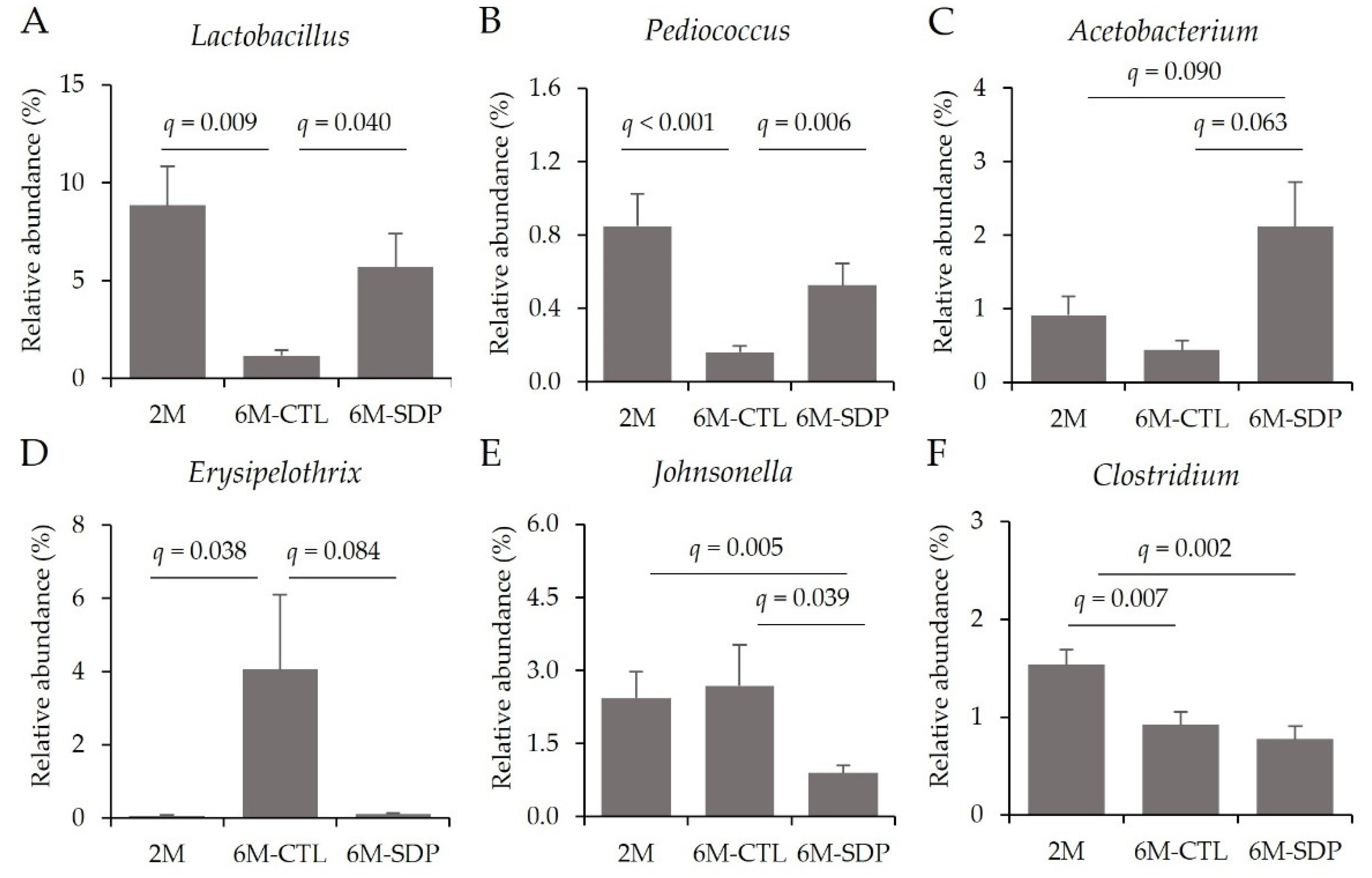
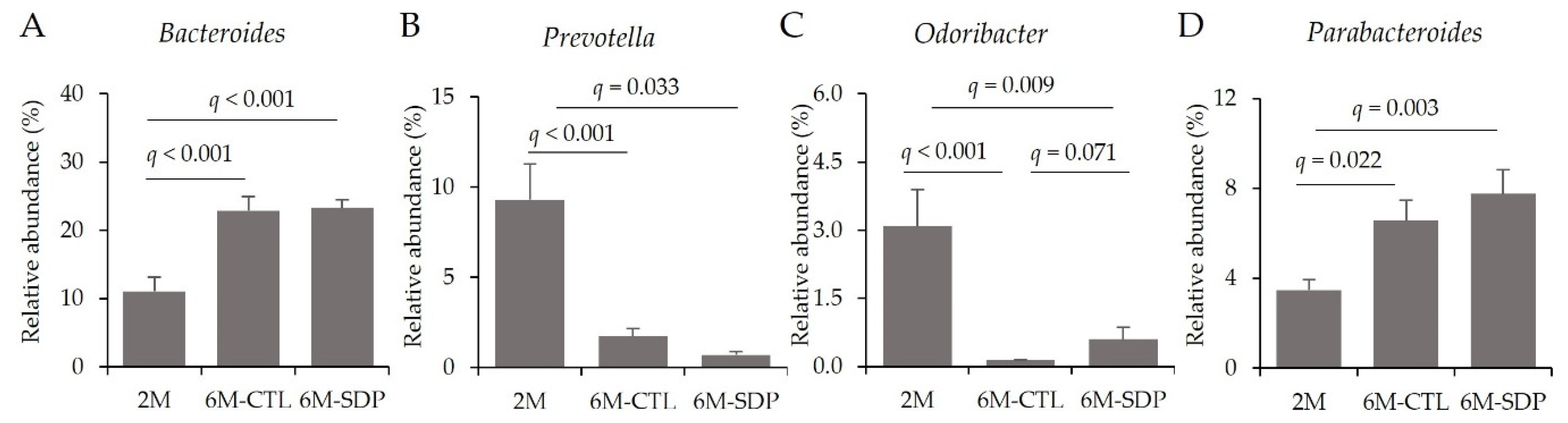
3.5. Gut Microbiota
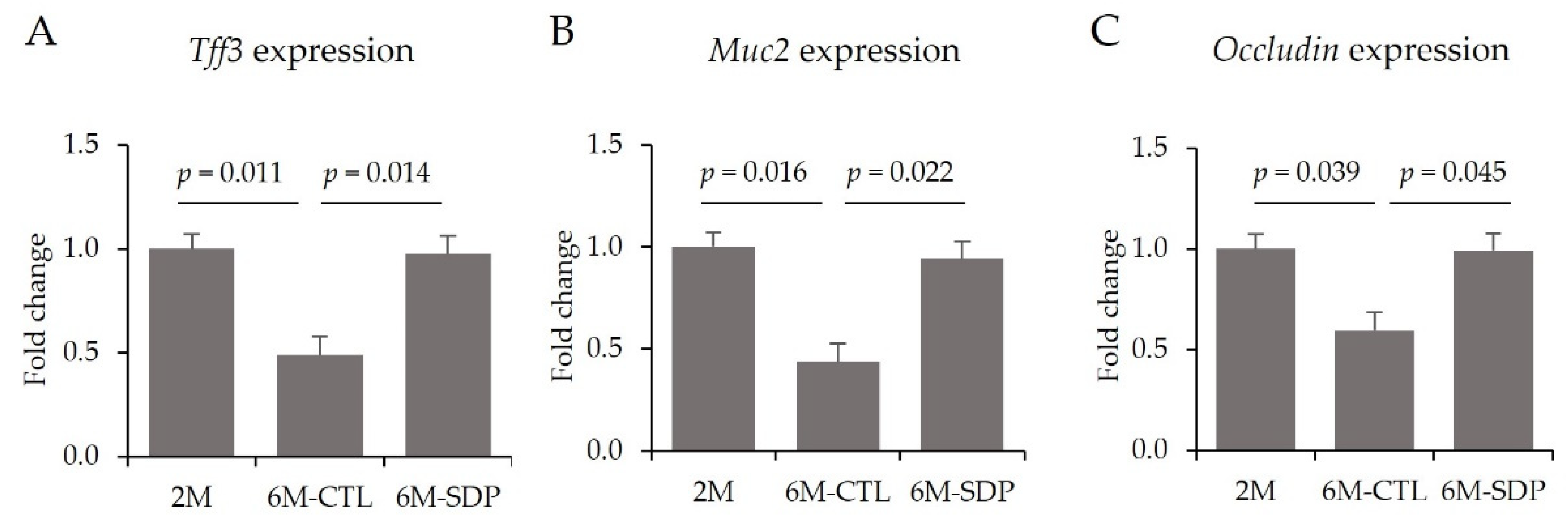
3.6. Effects on Colon Tissue
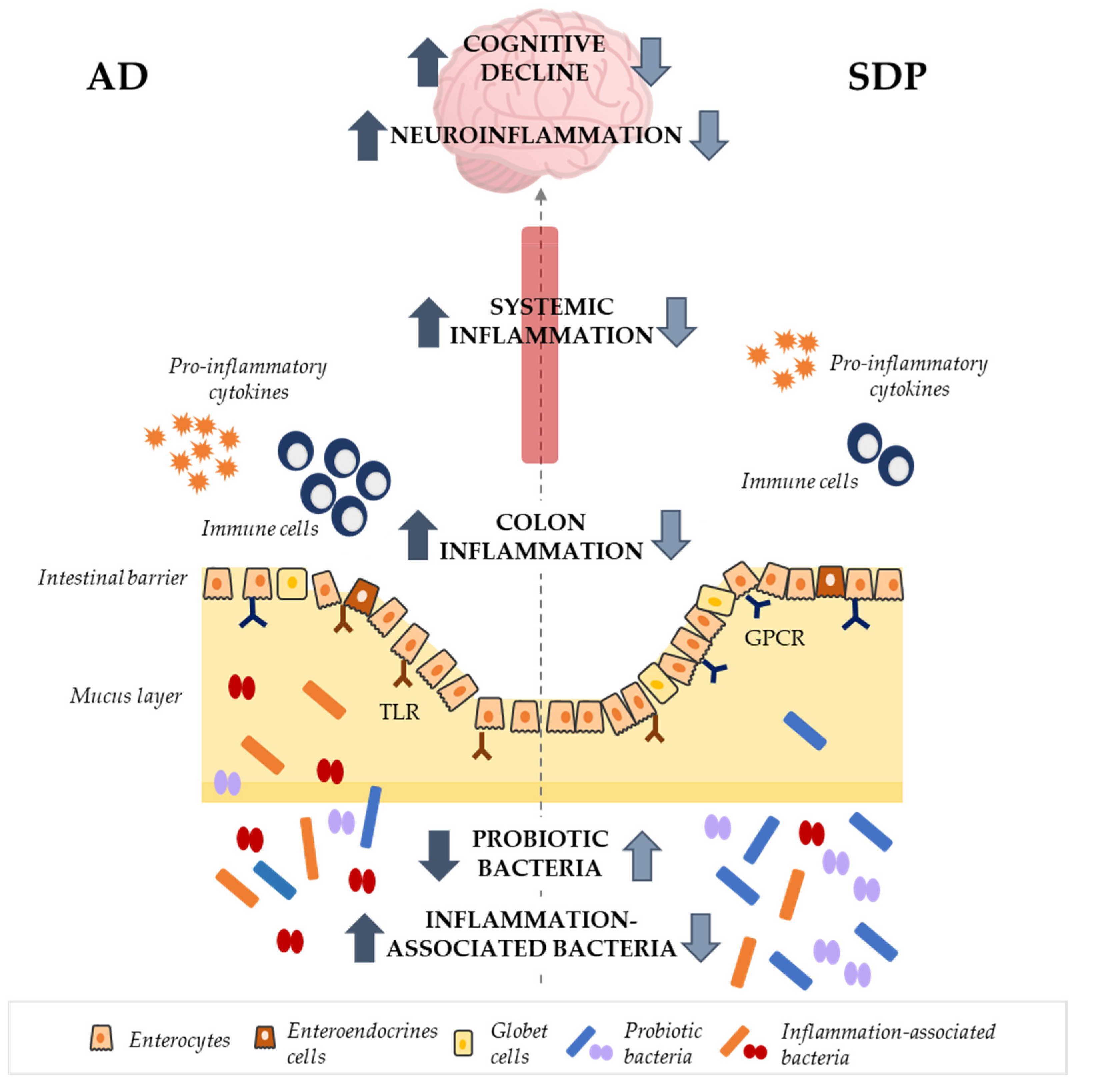
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimer’s Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Soria-Lopez, J.A.; González, H.M.; Léger, G.C. Alzheimer’s disease. In Handbook of Clinical Neurology, 3rd ed.; DeKosky, S.T., Asthana, S., Eds.; Elsevier: Boston, MA, USA, 2019; Volume 167, pp. 231–255. [Google Scholar] [CrossRef]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef] [PubMed]
- Biedermann, L.; Rogler, G. The intestinal microbiota: Its role in health and disease. Eur. J. Pediatr. 2015, 174, 151–167. [Google Scholar] [CrossRef]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bi, W.; Xiao, S.; Lan, X.; Cheng, X.; Zhang, J.; Lu, D.; Wei, W.; Wang, Y.; Li, H.; et al. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci. Rep. 2019, 9, 5790. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Zhao, S.; Zhu, Y.; Fan, Z.; Wang, J.; Zhang, B.; Chen, Y. Microbiota-gut-brain axis and toll-like receptors in Alzheimer’s disease. Comput. Struct. Biotechnol. J. 2019, 17, 1309–1317. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 2017, 7, 13537. [Google Scholar] [CrossRef]
- Zhan, G.; Yang, N.; Li, S.; Huang, N.; Fang, X.; Zhang, J.; Zhu, B.; Yang, L.; Yang, C.; Luo, A. Abnormal gut microbiota composition contributes to cognitive dysfunction in SAMP8 mice. Aging 2018, 10, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Kincaid, H.J.; Nagpal, R.; Yadav, H. Diet-Microbiota-Brain Axis in Alzheimer’s Disease. Ann. Nutr. Metab. 2021, 77 (Suppl. S2), 21–27. [Google Scholar] [CrossRef] [PubMed]
- Rosell-Cardona, C.; Griñan-Ferré, C.; Pérez-Bosque, A.; Polo, J.; Pallàs, M.; Amat, C.; Moretó, M.; Miró, L. Reply to Nifli, A.-P. Comment on “Rosell-Cardona et al. Dietary Spray-Dried Porcine Plasma Reduces Neuropathological Alzheimer’s Disease Hallmarks in SAMP8 Mice. Nutrients 2021, 13, 4065. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; González-Esquerra, R.; Polo, J. Impact of Spray-Dried Plasma on Intestinal Health and Broiler Performance. Microorganisms 2019, 7, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maijó, M.; Miró, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moretó, M.; Pérez-Bosque, A. Dietary Plasma Proteins Modulate the Adaptive Immune Response in Mice with Acute Lung Inflammation. J. Nutr. 2012, 142, 264–270. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Lee, J.J.; Che, T.M.; Soares-Almeida, J.A.; Chun, J.L.; Campbell, J.M.; Polo, J.; Crenshaw, J.D.; Seo, S.W.; et al. Spray-dried plasma attenuates inflammation and improves pregnancy rate of mated female mice1. J. Anim. Sci. 2015, 93, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Just, A.; Miró, L.; Pérez-Bosque, A.; Amat, C.; Polo, J.; Pallàs, M.; Griñán-Ferré, C.; Moretó, M. Dietary Spray-Dried Porcine Plasma Prevents Cognitive Decline in Senescent Mice and Reduces Neuroinflammation and Oxidative Stress. J. Nutr. 2020, 150, 303–311. [Google Scholar] [CrossRef]
- Rosell-Cardona, C.; Griñan-Ferré, C.; Pérez-Bosque, A.; Polo, J.; Pallàs, M.; Amat, C.; Moretó, M.; Miró, L. Dietary Spray-Dried Porcine Plasma Reduces Neuropathological Alzheimer’s Disease Hallmarks in SAMP8 Mice. Nutrients 2021, 13, 2369. [Google Scholar] [CrossRef]
- Miró, L.; Amat, C.; Rosell-Cardona, C.; Campbell, J.M.; Polo, J.; Pérez-Bosque, A.; Moretó, M. Dietary Supplementation with Spray-Dried Porcine Plasma Attenuates Colon Inflammation in A Genetic Mouse Model of Inflammatory Bowel Disease. Int. J. Mol. Sci. 2020, 21, 6760. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Miró, L.; Amat, C.; Polo, J.; Moretó, M. The Anti-Inflammatory Effect of Spray-Dried Plasma Is Mediated by a Reduction in Mucosal Lymphocyte Activation and Infiltration in a Mouse Model of Intestinal Inflammation. Nutrients 2016, 8, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretó, M.; Miró, L.; Amat, C.; Polo, J.; Manichanh, C.; Pérez-Bosque, A.; Moretó, M.; Miró, L.; Amat, C.; Polo, J.; et al. Dietary supplementation with spray-dried porcine plasma has prebiotic effects on gut microbiota in mice. Sci. Rep. 2020, 10, 2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, J.; Bodea, L.-G.; Goedert, M. Rodent models for Alzheimer disease. Nat. Rev. Neurosci. 2018, 19, 583–598. [Google Scholar] [CrossRef]
- Blázquez, E.; Rodríguez, C.; Ródenas, J.; Segalés, J.; Pujols, J.; Polo, J. Biosafety steps in the manufacturing process of spray-dried plasma: A review with emphasis on the use of ultraviolet irradiation as a redundant biosafety procedure. Porc. Health Manag. 2020, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Laboratory Animals, 4th ed.; National Academy Press: Washington, DC, USA, 1995. [Google Scholar]
- Puigoriol-Illamola, D.; Griñán-Ferré, C.; Vasilopoulou, F.; Leiva, R.; Vázquez, S.; Pallàs, M. 11β-HSD1 Inhibition by RL-118 Promotes Autophagy and Correlates with Reduced Oxidative Stress and Inflammation, Enhancing Cognitive Performance in SAMP8 Mouse Model. Mol. Neurobiol. 2018, 55, 8904–8915. [Google Scholar] [CrossRef]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Bosi, P.; Casini, L.; Finamore, A.; Cremokolini, C.; Merialdi, G.; Trevisi, P.; Nobili, F.; Mengheri, E. Spray-dried plasma improves growth performance and reduces inflammatory status of weaned pigs challenged with enterotoxigenic Escherichia coli K88. J. Anim. Sci. 2004, 82, 1764–1772. [Google Scholar] [CrossRef]
- Che, L.; Hu, L.; Zhou, Q.; Peng, X.; Liu, Y.; Luo, Y.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B.; et al. Microbial insight into dietary protein source affects intestinal function of pigs with intrauterine growth retardation. Eur. J. Nutr. 2020, 59, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Griñán-Ferré, C.; Marsal-García, L.; Bellver-Sanchis, A.; Kondengaden, S.M.; Turga, R.C.; Vázquez, S.; Pallàs, M. Pharmacological inhibition of G9a/GLP restores cognition and reduces oxidative stress, neuroinflammation and β-Amyloid plaques in an early-onset Alzheimer’s disease mouse model. Aging 2019, 11, 11591–11608. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, J.; Zuo, Y. Changes of Synaptic Structures Associated with Learning, Memory and Diseases. Brain Sci. Adv. 2018, 4, 99–117. [Google Scholar] [CrossRef]
- Singhal, G.; Morgan, J.; Jawahar, M.C.; Corrigan, F.; Jaehne, E.J.; Toben, C.; Breen, J.; Pederson, S.M.; Manavis, J.; Hannan, A.J.; et al. Effects of aging on the motor, cognitive and affective behaviors, neuroimmune responses and hippocampal gene expression. Behav. Brain Res. 2020, 383, 112501. [Google Scholar] [CrossRef]
- Tsumagari, R.; Maruo, K.; Nakao, T.; Ueda, S.; Yamanoue, M.; Shirai, Y. Motor Dyscoordination and Alteration of Functional Correlation Between DGKγ and PKCγ in Senescence-Accelerated Mouse Prone 8 (SAMP8). Front. Aging Neurosci. 2021, 13, 573966. [Google Scholar] [CrossRef]
- Liu, Y.; Choe, J.; Lee, J.J.; Kim, J.; Campbell, J.M.; Polo, J.; Crenshaw, J.D.; Pettigrew, J.E.; Song, M. Spray-dried plasma attenuates inflammation and lethargic behaviors of pregnant mice caused by lipopolysaccharide. PLoS ONE 2018, 13, e0203427. [Google Scholar] [CrossRef] [Green Version]
- Arancio, O.; Chao, M.V. Neurotrophins, synaptic plasticity and dementia. Curr. Opin. Neurobiol. 2007, 17, 325–330. [Google Scholar] [CrossRef]
- Tanila, H. The role of BDNF in Alzheimer’s disease. Neurobiol. Dis. 2017, 97 Pt B, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.-K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef]
- Savignac, H.M.; Corona, G.; Mills, H.; Chen, L.; Spencer, J.P.; Tzortzis, G.; Burnet, P.W. Prebiotic feeding elevates central brain derived neurotrophic factor, N-methyl-d-aspartate receptor subunits and d-serine. Neurochem. Int. 2013, 63, 756–764. [Google Scholar] [CrossRef] [Green Version]
- Letiembre, M.; Liu, Y.; Walter, S.; Hao, W.; Pfander, T.; Wrede, A.; Schulz-Schaeffer, W.; Fassbender, K. Screening of innate immune receptors in neurodegenerative diseases: A similar pattern. Neurobiol. Aging 2009, 30, 759–768. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Dansokho, C.; Heneka, M.T. Neuroinflammatory responses in Alzheimer’s disease. J. Neural Transm. 2018, 125, 771–779. [Google Scholar] [CrossRef]
- Kumar, V. Toll-like receptors in the pathogenesis of neuroinflammation. J. Neuroimmunol. 2019, 332, 16–30. [Google Scholar] [CrossRef]
- Srinivasan, M.; Lahiri, D.K. Significance of NF-κB as a pivotal therapeutic target in the neurodegenerative pathologies of Alzheimer’s disease and multiple sclerosis. Expert Opin. Ther. Targets 2015, 19, 471–487. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, C.; Santangelo, R. Alzheimer’s disease and gut microbiota modifications: The long way between preclinical studies and clinical evidence. Pharmacol. Res. 2018, 129, 329–336. [Google Scholar] [CrossRef]
- Walrath, T.; Dyamenahalli, K.U.; Hulsebus, H.J.; McCullough, R.L.; Idrovo, J.; Boe, D.M.; McMahan, R.H.; Kovacs, E.J. Age-related changes in intestinal immunity and the microbiome. J. Leukoc. Biol. 2020, 109, 1045–1061. [Google Scholar] [CrossRef]
- Schaubeck, M.; Clavel, T.; Calasan, J.; Lagkouvardos, I.; Haange, S.B.; Jehmlich, N.; Basic, M.; Dupont, A.; Hornef, M.; von Bergen, M.; et al. Dysbiotic gut microbiota causes transmissible Crohn’s disease-like ileitis independent of failure in antimicrobial defence. Gut 2016, 65, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Bäuerl, C.; Collado, M.C.; Diaz Cuevas, A.; Viña, J.; Pérez Martínez, G. Shifts in gut microbiota composition in an APP/PSS1 transgenic mouse model of Alzheimer’s disease during lifespan. Lett. Appl. Microbiol. 2018, 66, 464–471. [Google Scholar] [CrossRef]
- Shen, L.; Liu, L.; Ji, H.-F. Alzheimer’s Disease Histological and Behavioral Manifestations in Transgenic Mice Correlate with Specific Gut Microbiome State. J. Alzheimer’s Dis. 2017, 56, 385–390. [Google Scholar] [CrossRef]
- Rodriguez, C.; Blanch, F.; Romano, V.; Saborido, N.; Rodenas, J.; Polo, J. Porcine immunoglobulins survival in the intestinal tract of adult dogs and cats fed dry food kibbles containing spray-dried porcine plasma (SDPP) or porcine immunoglobulin concentrate (PIC). Anim. Feed Sci. Technol. 2007, 139, 201–211. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Quan, M.; Zhao, H.; Jia, J. Gut Microbiota Changes and Their Correlation with Cognitive and Neuropsychiatric Symptoms in Alzheimer’s Disease. J. Alzheimer’s Dis. 2021, 81, 583–595. [Google Scholar] [CrossRef]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yu, D.; Xue, L.; Li, H.; Du, J. Probiotics modulate the microbiota–gut–brain axis and improve memory deficits in aged SAMP8 mice. Acta Pharm. Sin. B 2020, 10, 475–487. [Google Scholar] [CrossRef]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Jin, F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef]
- Ross, D.E.; Marshall, C.W.; Gulliver, D.; May, H.D.; Norman, R.S. Defining Genomic and Predicted Metabolic Features of the Acetobacterium Genus. mSystems 2020, 5, e00277-20. [Google Scholar] [CrossRef]
- Currais, A.; Huang, L.; Goldberg, J.; Petrascheck, M.; Ates, G.; Pinto-Duarte, A.; Shokhirev, M.N.; Schubert, D.; Maher, P. Elevating acetyl-CoA levels reduces aspects of brain aging. eLife 2019, 8, e47866. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, A.; Nakatani, A.; Hasegawa, S.; Irie, J.; Ozawa, K.; Tsujimoto, G.; Suganami, T.; Itoh, H.; Kimura, I. The short chain fatty acid receptor GPR43 regulates inflammatory signals in adipose tissue M2-type macrophages. PLoS ONE 2017, 12, e0179696. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, K.; Mulak, A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Krstic, D.; Madhusudan, A.; Doehner, J.; Vogel, P.; Notter, T.; Imhof, C.; Manalastas, A.; Hilfiker, M.; Pfister, S.; Schwerdel, C.; et al. Systemic immune challenges trigger and drive Alzheimer-like neuropathology in mice. J. Neuroinflamm. 2012, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Marizzoni, M.; Cattaneo, A.; Mirabelli, P.; Festari, C.; Lopizzo, N.; Nicolosi, V.; Mombelli, E.; Mazzelli, M.; Luongo, D.; Naviglio, D.; et al. Short-Chain Fatty Acids and Lipopolysaccharide as Mediators Between Gut Dysbiosis and Amyloid Pathology in Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 78, 683–697. [Google Scholar] [CrossRef]
- Khailova, L.; Baird, C.H.; Rush, A.A.; Barnes, C.; Wischmeyer, P.E. Lactobacillus rhamnosus GG treatment improves intestinal permeability and modulates inflammatory response and homeostasis of spleen and colon in experimental model of Pseudomonas aeruginosa pneumonia. Clin. Nutr. 2017, 36, 1549–1557. [Google Scholar] [CrossRef]
- Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigón, G.D.V.; Cazorla, S.I.; Maldonado-Galdeano, C.; Weill, R.; De Paula, J.; Perdigón, G.D.V. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Front. Microbiol. 2018, 9, 736. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Amat, C.; Polo, J.; Campbell, J.M.; Crenshaw, J.; Russell, L.; Moretó, M. Spray-Dried Animal Plasma Prevents the Effects of Staphylococcus aureus Enterotoxin B on Intestinal Barrier Function in Weaned Rats. J. Nutr. 2006, 136, 2838–2843. [Google Scholar] [CrossRef]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune–metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- Swardfager, W.; Lanctôt, K.; Rothenburg, L.; Wong, A.; Cappell, J.; Herrmann, N. A Meta-Analysis of Cytokines in Alzheimer’s Disease. Biol. Psychiatry 2010, 68, 930–941. [Google Scholar] [CrossRef]
- Park, J.-C.; Han, S.-H.; Mook-Jung, I. Peripheral inflammatory biomarkers in Alzheimer’s disease: A brief review. BMB Rep. 2020, 53, 10–19. [Google Scholar] [CrossRef]
- Wang, F.; Graham, W.V.; Wang, Y.; Witkowski, E.D.; Schwarz, B.T.; Turner, J.R. Interferon-γ and Tumor Necrosis Factor-α Synergize to Induce Intestinal Epithelial Barrier Dysfunction by Up-Regulating Myosin Light Chain Kinase Expression. Am. J. Pathol. 2005, 166, 409–419. [Google Scholar] [CrossRef]
- Laukoetter, M.G.; Nava, P.; Nusrat, A. Role of the intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 401–407. [Google Scholar] [CrossRef]
- Miró, L.; Garcia-Just, A.; Amat, C.; Polo, J.; Moretó, M.; Pérez-Bosque, A. Dietary Animal Plasma Proteins Improve the Intestinal Immune Response in Senescent Mice. Nutrients 2017, 9, 1346. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| q | |||||||
|---|---|---|---|---|---|---|---|
| Phylum | Family | 2M | 6M-CTL | 6M-SDP | 2M 6M-CTL | 2M 6M-SDP | 6M-CTL 6M-SDP |
| Firmicutes | Lactobacillaceae | 10.1 ± 2.2 | 1.3 ± 0.3 | 6.6 ± 2.0 | 0.003 | NS | 0.019 |
| Eubacteriaceae | 0.8 ± 0.2 | 0.4 ± 0.1 | 2.0 ± 0.6 | NS | NS | 0.033 | |
| Erysipelotrichaceae | 0.1 ± 0.01 | 3.9 ± 1.9 | 0.5 ± 0.2 | 0.029 | NS | NS | |
| Lachnospiraceae | 20.1 ± 2.2 | 23.2 ± 2.2 | 17.6 ± 1.7 | NS | NS | NS | |
| Clostridiaceae | 6.7 ± 0.6 | 4.0 ± 0.4 | 4.7 ± 0.7 | 0.008 | 0.030 | NS | |
| Ruminococcaceae | 4.6 ± 0.5 | 3.6 ± 0.5 | 3.8 ± 0.6 | NS | NS | NS | |
| Bacteroidetes | Bacteroidaceae | 10.2 ± 1.9 | 21.2 ± 1.9 | 21.8 ± 2.0 | <0.001 | <0.001 | NS |
| Porphyromonadaceae | 8.0 ± 1.3 | 11.3 ± 1.0 | 12.1 ± 1.2 | 0.082 | 0.060 | NS | |
| Sphingobacteriaceae | 5.5 ± 0.6 | 3.8 ± 0.3 | 4.3 ± 0.3 | 0.036 | NS | NS | |
| Flexibacteraceae | 0.3 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.1 | NS | NS | NS | |
| Flavobacteriaceae | 4.2 ± 0.6 | 2.2 ± 0.3 | 2.1 ± 0.4 | 0.005 | 0.005 | NS | |
| Odoribacteriaceae | 3.2 ± 0.7 | 1.9 ± 0.3 | 2.4 ± 0.5 | NS | NS | NS | |
| Prevotellaceae | 9.2 ± 2.2 | 1.6 ± 0.4 | 0.6 ± 0.2 | 0.018 | 0.017 | NS | |
| Proteobacteria | Desulfovibrionaceae | 1.9 ± 0.4 | 2.6 ± 0.4 | 3.1 ± 0.4 | NS | NS | NS |
| Enterobacteriaceae | 0.6 ± 0.1 | 0.5 ± 0.1 | 0.6 ± 0.1 | NS | NS | NS | |
| Helicobacteriaceae | 3.6 ± 0.7 | 1.4 ± 0.3 | 0.8 ± 0.3 | 0.004 | <0.001 | NS | |
| Verrucomicrobia | Verrucomicrobiaceae | 0.03 ± 0.01 | 3.2 ± 1.6 | 8.1 ± 1.9 | NS | 0.016 | NS |
| Actinobacteria | Bifidobacteriaceae | 0.05 ± 0.01 | 0.5 ± 0.3 | 0.05 ± 0.01 | NS | NS | 0.069 |
| q | |||||||
|---|---|---|---|---|---|---|---|
| Family | Species | 2M | 6M-CTL | 6M-SDP | 2M | 2M | 6M-CTL |
| 6M-CTL | 6M-SDP | 6M-SDP | |||||
| Lactobacillaceae | Lactobacillus hayakitensis | 5.1 ± 1.7 | 0.9 ± 0.2 | 2.0 ± 0.6 | NS | NS | NS |
| Lactobacillus taiwanensis | 1.9 ± 0.7 | 0.2 ± 0.1 | 0.5 ± 0.3 | 0.003 | 0.070 | NS | |
| Lactobacillus siliginis | 1.8 ± 0.5 | 0.3 ± 0.06 | 0.8 ± 0.2 | 0.035 | 0.075 | 0.047 | |
| Lactobacillus antri | 1.1 ± 0.3 | 0.1 ± 0.02 | 0.3 ± 0.2 | 0.018 | 0.016 | NS | |
| Lactobacillus intermedius | 0.5 ± 0.2 | 0.1 ± 0.03 | 0.3 ± 0.1 | NS | NS | NS | |
| Pediococcus argentinicus | 0.5 ± 0.1 | 0.08 ± 0.04 | 0.2 ± 0.05 | 0.009 | NS | NS | |
| Erysipelotrichaceae | Erysipelhotrix muris | 0.1 ± 0.02 | 8.9 ± 3.8 | 0.2 ± 0.1 | 0.020 | NS | 0.046 |
| Prevotellaceae | Prevotella dentasini | 16.0 ± 3.7 | 1.6 ± 0.4 | 0.7 ± 0.2 | 0.005 | 0.005 | 0.044 |
| Bacteroidaceae | Bacteroides xylanisolvens | 2.9 ± 0.4 | 6.6 ± 1.2 | 8.7 ± 1.3 | 0.031 | 0.002 | NS |
| Bacteroides rodentium | 2.7 ± 0.6 | 3.2 ± 0.9 | 5.6 ± 1.3 | NS | NS | NS | |
| Bacteroides acidifaciens | 0.5 ± 0.1 | 4.1 ± 0.7 | 4.6 ± 0.9 | 0.001 | 0.001 | NS | |
| Bacteroides sartorii | 0.3 ± 0.1 | 1.1 ± 0.3 | 1.3 ± 0.1 | 0.011 | 0.002 | NS | |
| Bacteroides denticanum | 0.9 ± 0.1 | 6.5 ± 1.1 | 3.1 ± 0.6 | 0.001 | 0.009 | 0.015 | |
| Porphyromonadaceae | Parabacteroides goldsteinii | 4.9 ± 0.6 | 6.0 ± 0.7 | 10.6 ± 1.3 | NS | 0.005 | 0.013 |
| Parabacteroides gordonii | 0.0 ± 0.0 | 3.6 ± 0.8 | 1.7 ± 0.7 | <0.001 | 0.014 | 0.070 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosell-Cardona, C.; Amat, C.; Griñán-Ferré, C.; Polo, J.; Pallàs, M.; Pérez-Bosque, A.; Moretó, M.; Miró, L. The Neuroprotective Effects of Spray-Dried Porcine Plasma Supplementation Involve the Microbiota−Gut−Brain Axis. Nutrients 2022, 14, 2211. https://doi.org/10.3390/nu14112211
Rosell-Cardona C, Amat C, Griñán-Ferré C, Polo J, Pallàs M, Pérez-Bosque A, Moretó M, Miró L. The Neuroprotective Effects of Spray-Dried Porcine Plasma Supplementation Involve the Microbiota−Gut−Brain Axis. Nutrients. 2022; 14(11):2211. https://doi.org/10.3390/nu14112211
Chicago/Turabian StyleRosell-Cardona, Cristina, Concepció Amat, Christian Griñán-Ferré, Javier Polo, Mercè Pallàs, Anna Pérez-Bosque, Miquel Moretó, and Lluïsa Miró. 2022. "The Neuroprotective Effects of Spray-Dried Porcine Plasma Supplementation Involve the Microbiota−Gut−Brain Axis" Nutrients 14, no. 11: 2211. https://doi.org/10.3390/nu14112211