
Beneficial Effects of Walnut Oligopeptides on Muscle Loss in Senescence-Accelerated Mouse Prone-8 (SAMP8) Mice: Focusing on Mitochondrial Function



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of WOPs Sample
2.2. Animals and Treatments
2.3. Muscle Mass Measurement
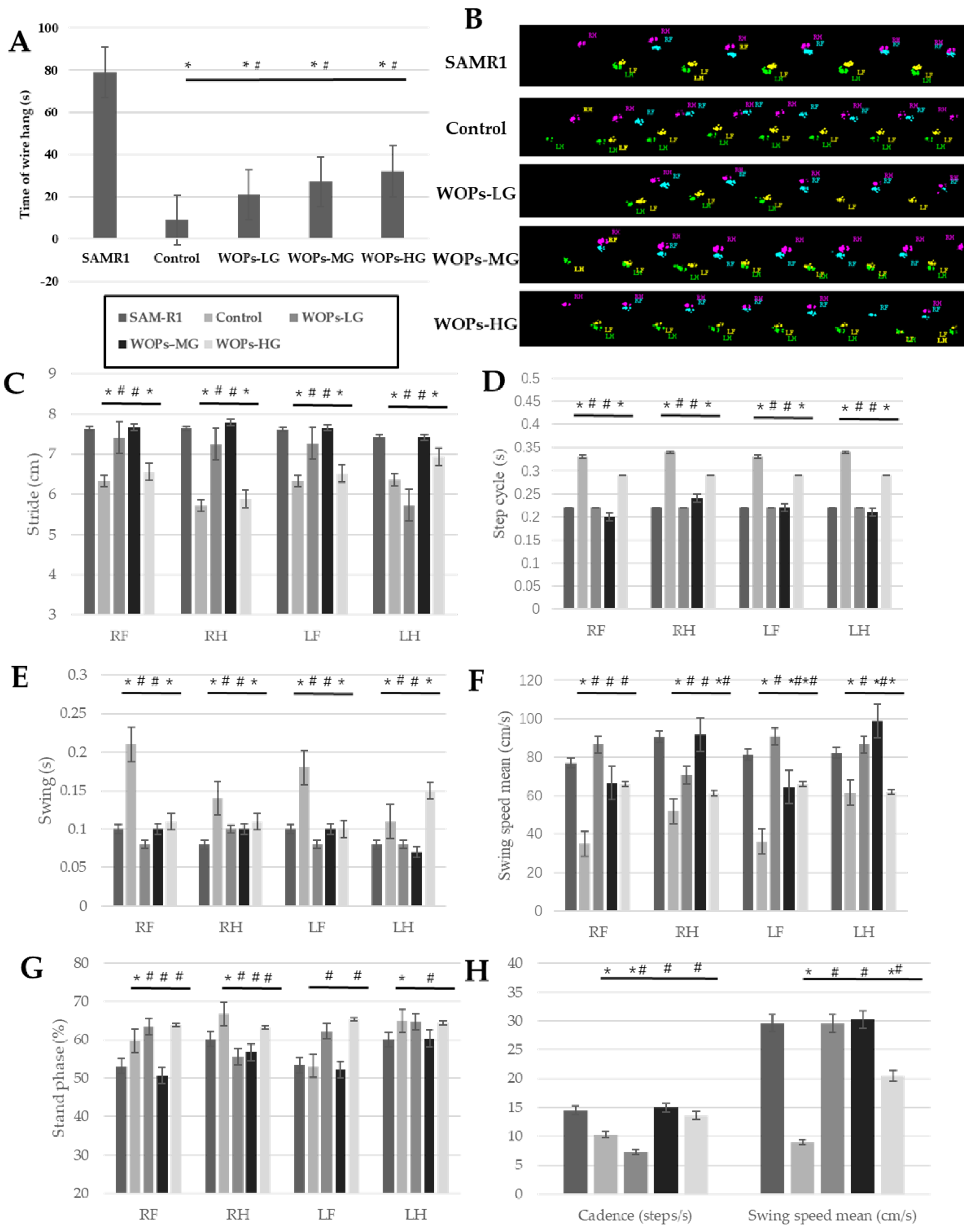
2.4. Wire Hang and Catwalk Test
2.5. Enzyme-Linked Immunosorbent Assay in Serum and Gastrocnemius
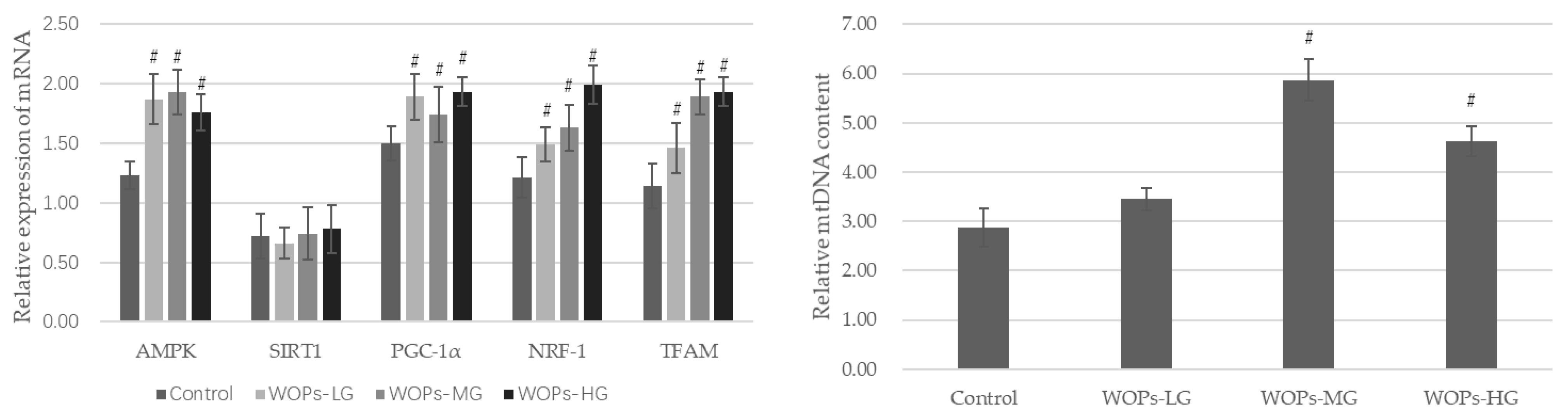
2.6. Quantitative Real-Time PCR and Analyses of mtDNA Content in Gastrocnemius
2.7. Statistical Analysis
3. Results
3.1. Effects of WOPs on Body Composition and Gastrocnemius (%Body Weight) in Mice
3.2. Effects of WOPs on the Physical Performance
3.3. Effects of WOPs on Proinflammatory Factors in Serum and Gastrocnemius
3.4. Effects of WOPs on Mitochondrial Function Factors and mtDNA Content in the Gastrocnemius
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Partridge, L.; Deelen, J.; Slagboom, P.E. Facing up to the global challenges of ageing. Nature 2018, 561, 45–56. [Google Scholar] [CrossRef] [PubMed]
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Ageing 2013; United Nations: New York, NY, USA, 2013. [Google Scholar]
- Chen, H.; Chen, G.; Zheng, X.; Guo, Y. Contribution of specific diseases and injuries to changes in health adjusted life expectancy in 187 countries from 1990 to 2013: Retrospective observational study. BMJ 2019, 364, l969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cawthon, P.M.; Lui, L.Y.; Taylor, B.C.; McCulloch, C.E.; Cauley, J.A.; Lapidus, J.; Orwoll, E.; Ensrud, K.E. Clinical Definitions of Sarcopenia and Risk of Hospitalization in Community-Dwelling Older Men: The Osteoporotic Fractures in Men Study. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, I.; Heymsfield, S.B.; Ross, R. Low Relative Skeletal Muscle Mass (Sarcopenia) in Older Persons Is Associated with Functional Impairment and Physical Disability. J. Am. Geriatr. Soc. 2002, 50, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Landi, F.; Cruz-Jentoft, A.J.; Liperoti, R.; Russo, A.; Giovannini, S.; Tosato, M.; Capoluongo, E.D.; Bernabei, R.; Onder, G. Sarcopenia and mortality risk in frail older persons aged 80 years and older: Results from ilSIRENTE study. Age Ageing 2013, 42, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.K.; Woo, J.; Assantachai, P.; Auyeung, T.-W.; Chou, M.-Y.; Iijima, K.; Jang, H.C.; Kang, L.; Kim, M.; Kim, S.; et al. Asian Working Group for Sarcopenia: 2019 Consensus Update on Sarcopenia Diagnosis and Treatment. J. Am. Med. Dir. Assoc. 2020, 21, 300–307. [Google Scholar] [CrossRef]
- Wu, X.; Li, X.; Xu, M.; Zhang, Z.; He, L.; Li, Y. Sarcopenia prevalence and associated factors among older Chinese population: Findings from the China Health and Retirement Longitudinal Study. PLoS ONE 2021, 16, e0247617. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Landi, F.; Schneider, S.M.; Zúñiga, C.; Arai, H.; Boirie, Y.; Chen, L.-K.; Fielding, R.A.; Martin, F.C.; Michel, J.-P.; et al. Prevalence of and interventions for sarcopenia in ageing adults: A systematic review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing 2014, 43, 748–759. [Google Scholar] [CrossRef]
- Dennison, E.M.; Sayer, A.A.; Cooper, C. Epidemiology of sarcopenia and insigh into possible therapeutic targets. Nat. Rev. Rheumatol. 2017, 13, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zheng, X.; Aihemaitijiang, S.; Wang, R.; Halimulati, M.; Huang, X.; Zhang, Z. Sarcopenia and catastrophic health expenditure by socio-economic groups in China: An analysis of household-based panel data. J. Cachex-Sarcopenia Muscle 2022. [Google Scholar] [CrossRef]
- Arai, H.; Wakabayashi, H.; Yoshimura, Y.; Yamada, M.; Kim, H.; Harada, A. Chapter 4 Treatment of sarcopenia. Geriatr. Gerontol. Int. 2018, 18 (Suppl. S1), 28–44. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Shih, M.-H.; Chen, C.-D.; Yeh, S.-L. Effects of adequate dietary protein with whey protein, leucine, and vitamin D supplementation on sarcopenia in older adults: An open-label, parallel-group study. Clin. Nutr. 2020, 40, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Crosignani, S.; Sedini, C.; Calvani, R.; Marzetti, E.; Cesari, M. Sarcopenia in primary care: Screening, diagnosis, management. J. Frailty Aging 2021, 10, 226–232. [Google Scholar] [PubMed]
- Kramer, I.F.; Verdijk, L.B.; Hamer, H.M.; Verlaan, S.; Luiking, Y.C.; Kouw, I.W.K.; Senden, J.M.; van Kranenburg, J.; Gijsen, A.P.; Bierau, J.; et al. Both basal and post-prandial muscle protein synthesis rates, following the ingestion of a leucine-enriched whey protein supplement, are not impaired in sarcopenic older males. Clin. Nutr. 2017, 36, 1440–1449. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Bukhari, S.S.I.; Phillips, B.E.; Limb, M.C.; Cegielski, J.; Brook, M.S.; Rankin, D.; Mitchell, W.K.; Kobayashi, H.; Williams, J.P.; et al. Effects of leucine-enriched essential amino acid and whey protein bolus dosing upon skeletal muscle protein synthesis at rest and after exercise in older women. Clin. Nutr. 2018, 37, 2011–2021. [Google Scholar] [CrossRef] [Green Version]
- Reitelseder, S.; Dideriksen, K.; Agergaard, J.; Malmgaard-Clausen, N.M.; Bechshoeft, R.L.; Petersen, R.K.; Serena, A.; Mikkelsen, U.R.; Holm, L. Even effect of milk protein and carbohydrate intake but no further effect of heavy resistance exercise on myofibrillar protein synthesis in older men. Eur. J. Nutr. 2019, 58, 583–595. [Google Scholar] [CrossRef]
- Borack, M.S.; Reidy, P.T.; Husaini, S.H.; Markofski, M.M.; Deer, R.R.; Richison, A.B.; Lambert, B.S.; Cope, M.B.; Mukherjea, R.; Jennings, K.; et al. Soy-Dairy Protein Blend or Whey Protein Isolate Ingestion Induces Similar Postexercise Muscle Mechanistic Target of Rapamycin Complex 1 Signaling and Protein Synthesis Responses in Older Men. J. Nutr. 2016, 146, 2468–2475. [Google Scholar] [CrossRef] [Green Version]
- Walrand, S.; Zangarelli, A.; Guillet, C.; Salles, J.; Soulier, K.; Giraudet, C.; Patrac, V.; Boirie, Y. Effect of fast dietary proteins on muscle protein synthesis rate and muscle strength in ad libitum-fed and energy-restricted old rats. Br. J. Nutr. 2011, 106, 1683–1690. [Google Scholar] [CrossRef] [Green Version]
- Kerasioti, E.; Stagos, D.; Priftis, A.; Aivazidis, S.; Tsatsakis, A.M.; Hayes, A.W.; Kouretas, D. Antioxidant effects of whey protein on muscle C2C12 cells. Food Chem. 2014, 155, 271–278. [Google Scholar] [CrossRef]
- Xu, R.; Liu, N.; Xu, X.; Kong, B. Antioxidative effects of whey protein on peroxide-induced cytotoxicity. J. Dairy Sci. 2011, 94, 3739–3746. [Google Scholar] [CrossRef] [Green Version]
- Admassu, H.; Gasmalla, M.A.A.; Yang, R.; Zhao, W. Bioactive Peptides Derived from Seaweed Protein and Their Health Benefits: Antihypertensive, Antioxidant, and Antidiabetic Properties. J. Food Sci. 2017, 83, 6–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmartin, S.; O’Brien, N.; Giblin, L. Whey for Sarcopenia; Can Whey Peptides, Hydrolysates or Proteins Play a Beneficial Role? Foods 2020, 9, 750. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Liu, R.; He, L.-X.; Mao, R.X.; Liu, L.-R.; Zhang, T.; Hao, Y.-T.; Fan, R.; Xu, M.-H.; Li, Y. Radioprotective Effect of Walnut Oligopeptides Against Gamma Radiation-Induced Splenocyte Apoptosis and Intestinal Injury in Mice. Molecules 2019, 24, 1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Ren, J.; Wang, T.; Wu, L.; Liu, P.; Li, Y. Anti-hypoxia effects of walnut oligopeptides (Juglans regia L.) in mice. Am. J. Transl. Res. 2021, 13, 4581–4590. [Google Scholar]
- Ding, T.; Li, Y. Beneficial effect and mechanism of walnut oligopeptide on Lactobacillus plantarum Z7. Food Sci. Nutr. 2021, 9, 672–681. [Google Scholar] [CrossRef]
- Liu, R.; Hao, Y.-T.; Zhu, N.; Liu, X.-R.; Kang, J.-W.; Mao, R.-X.; Hou, C.; Li, Y. The Gastroprotective Effect of Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) against Ethanol-Induced Gastric Mucosal Injury in Rats. Nutrients 2020, 12, 1138. [Google Scholar] [CrossRef]
- Liu, R.; Wu, L.; Du, Q.; Ren, J.-W.; Chen, Q.-H.; Li, D.; Mao, R.-X.; Liu, X.-R.; Li, Y. Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) and Their Anti-Fatigue Effects in Mice. Molecules 2018, 24, 45. [Google Scholar] [CrossRef] [Green Version]
- Brennan, J.L.; Keerati-U-Rai, M.; Yin, H.; Daoust, J.; Nonnotte, E.; Quinquis, L.; St-Denis, T.; Bolster, D.R. Differential Responses of Blood Essential Amino Acid Levels Following Ingestion of High-Quality Plant-Based Protein Blends Compared to Whey Protein—A Double-Blind Randomized, Cross-Over, Clinical Trial. Nutrients 2019, 11, 2987. [Google Scholar] [CrossRef] [Green Version]
- Nabuco, H.; Tomeleri, C.M.; Fernandes, R.R.; Junior, S.P.; Venturini, D.; Barbosa, D.S.; Deminice, R.; Sardinha, L.B.; Cyrino, E.S. Effects of pre- or post-exercise whey protein supplementation on oxidative stress and antioxidant enzymes in older women. Scand. J. Med. Sci. Sports 2019, 29, 1101–1108. [Google Scholar] [CrossRef]
- Corpuz, H.M.; Arimura, M.; Chawalitpong, S.; Miyazaki, K.; Sawaguchi, M.; Nakamura, S.; Katayama, S. Oral Administration of Okara Soybean By-Product Attenuates Cognitive Impairment in a Mouse Model of Accelerated Aging. Nutrients 2019, 11, 2939. [Google Scholar] [CrossRef] [Green Version]
- Vela, S.; Neira, S.; Moreno-Aliaga, M.J.; Solas, M.; Ramírez, M.J. DHA Selectively Protects SAMP-8-Associated Cognitive Deficits Through Inhibition of JNK. Mol. Neurobiol. 2019, 56, 1618–1627. [Google Scholar] [CrossRef] [PubMed]
- Toricelli, M.; Evangelista, S.R.; Buck, H.S.; Viel, T.A. Microdose Lithium Treatment Reduced Inflammatory Factors and Neurodegeneration in Organotypic Hippocampal Culture of Old SAMP-8 Mice. Cell. Mol. Neurobiol. 2021, 41, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- Derave, W.; Eijnde, B.O.; Ramaekers, M.; Hespel, P. Soleus muscles of SAMP8 mice provide an accelerated model of skeletal muscle senescence. Exp. Gerontol. 2005, 40, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Yun, G.A.; Leung, K.S.; Siu, P.M.F.; Qin, J.H.; Chow, S.K.H.; Qin, L.; Li, C.Y.; Cheung, W.H. Muscle mass, structural and functional investigations of senescence-accelerated mouse P8 (SAMP8). Exp. Anim. 2015, 64, 425–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzetti, E.; Calvani, R.; Cesari, M.; Buford, T.B.; Lorenzi, M.; Behnke, B.J.; Leeuwenburgh, C. Mitochondrial dysfunction and sarcopenia of aging: From signaling pathways to clinical trials. Int. J. Biochem. Cell Biol. 2013, 45, 2288–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzetti, E. Musculoskeletal Aging and Sarcopenia in the Elderly. Int. J. Mol. Sci. 2022, 23, 2808. [Google Scholar] [CrossRef]
- Fan, R.; Kang, J.; Hao, Y.; Liu, X.; Hu, J.; Mao, R.; Liu, R.; Zhu, N.; Xu, M.; Li, Y. Undenatured type II collagen prevents and treats osteoarthritis and motor function degradation in T2DM patients and db/db mice. Food Funct. 2021, 12, 4373–4391. [Google Scholar]
- Fiander, M.D.; Stifani, N.; Nichols, M.; Akay, T.; Robertson, G.S. Kinematic gait parameters are highly sensitive measures of motor deficits and spinal cord injury in mice subjected to experimental autoimmune encephalomyelitis. Behav. Brain Res. 2017, 317, 95–108. [Google Scholar] [CrossRef]
- Li, C.-W.; Yu, K.; Shyh-Chang, N.; Li, G.-X.; Jiang, L.-J.; Yu, S.-L.; Xu, L.-Y.; Liu, R.-J.; Guo, Z.-J.; Xie, H.-Y.; et al. Circulating factors associated with sarcopenia during ageing and after intensive lifestyle intervention. J. Cachexia-Sarcopenia Muscle 2019, 10, 586–600. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef]
- Beyer, I.; Mets, T.; Bautmans, I. Chronic low-grade inflammation and age-related sarcopenia. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Aleman, H.; Esparza, J.; Ramirez, F.A.; Astiazaran, H.; Payette, H. Longitudinal evidence on the association between interleukin-6 and C-reactive protein with the loss of total appendicular skeletal muscle in free-living older men and women. Age Ageing 2011, 40, 469–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haren, M.T.; Malmstrom, T.K.; Miller, D.K.; Patrick, P.; Perry, H.M.; Herning, M.M.; Banks, W.A.; Morley, J.E. Higher C-reactive protein and soluble tumor necrosis factor receptor levels are associated with poor physical function and disability: A cross-sectional analysis of a cohort of late middle-aged African Americans. J. Gerontol. Ser. A 2010, 65, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto-Sen, S.; Kawakami, S.; Maruki-Uchida, H.; Ito, R.; Matsui, N.; Komiya, Y.; Mita, Y.; Morisasa, M.; Goto-Inoue, N.; Furuichi, Y.; et al. Effect of antioxidant supplementation on skeletal muscle and metabolic profile in aging mice. Food Funct. 2021, 12, 825–833. [Google Scholar] [CrossRef]
- Kinoshita, H.; Orita, S.; Inage, K.; Yamauchi, K.; Abe, K.; Inoue, M.; Norimoto, M.; Umimura, T.; Eguchi, Y.; Fujimoto, K.; et al. Skeletal Muscle Cell Oxidative Stress as a Possible Therapeutic Target in a Denervation-Induced Experimental Sarcopenic Model. Spine 2019, 44, E446–E455. [Google Scholar] [CrossRef]
- Li, C.; Yu, K.; Shyh-Chang, N.; Li, G.; Yu, S.; Liu, H.; Yang, B.; Li, Z.; Zhao, Y.; Xu, L.; et al. Sterol metabolism and protein metabolism are differentially correlated with sarcopenia in Asian Chinese men and women. Cell Prolif. 2021, 54, e12989. [Google Scholar] [CrossRef]
- Villota-Narvaez, Y.; Garzon-Alvarado, D.A.; Ramirez-Martinez, A.M. A dynamical system for the IGF1-AKT signaling pathway in skeletal muscle adaptation. Biosystems 2021, 202, 104355. [Google Scholar] [CrossRef]
- Mankhong, S.; Kim, S.; Moon, S.; Kwak, H.-B.; Park, D.-H.; Kang, J.-H. Experimental Models of Sarcopenia: Bridging Molecular Mechanism and Therapeutic Strategy. Cells 2020, 9, 1385. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Penna, F.; Busquets, S.; Pin, F.; Toledo, M.; Baccino, F.M.; López-Soriano, F.J.; Costelli, P.; Argilés, J.M. Combined approach to counteract experimental cancer cachexia: Eicosapentaenoic acid and training exercise. J. Cachex-Sarcopenia Muscle 2011, 2, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Scarpulla, R.C. Transcriptional Paradigms in Mammalian Mitochondrial Biogenesis and Function. Physiol. Rev. 2008, 88, 611–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.-W.; Chang, Y.-C.; Chan, Y.-C.; Hu, S.-H.; Liu, M.-Y.; Chang, S.-J. Dysregulations of mitochondrial quality control and autophagic flux at an early age lead to progression of sarcopenia in SAMP8 mice. Biogerontology 2020, 21, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Koo, Y.D.; Lim, S.; Park, Y.J.; Chung, S.S.; Jang, H.C.; Park, K.S. Rg3 Improves mitochondrial function and the expression of key genes involved in mitochondrial biogenesis in C2C12 myotubes. Diabetes Metab. J. 2016, 40, 406–413. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Lean Mass (%Body Weight) | Gastrocnemius (%Body Weight) | |
|---|---|---|
| SAMR1 | 85.41 ± 0.41 | 1.37 ± 0.13 |
| Control | 81.22 ± 0.23 * | 0.98 ± 0.11 * |
| WOPs-LG | 83.12 ± 0.30 # | 1.21 ± 0.14 # |
| WOPs-MG | 82.41 ± 0.12 * | 1.26 ± 0.12 # |
| WOPs-HG | 84.17 ± 0.19 # | 1.19 ± 0.09 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, R.; Hao, Y.; Du, Q.; Kang, J.; Xu, M.; Li, Y. Beneficial Effects of Walnut Oligopeptides on Muscle Loss in Senescence-Accelerated Mouse Prone-8 (SAMP8) Mice: Focusing on Mitochondrial Function. Nutrients 2022, 14, 2051. https://doi.org/10.3390/nu14102051
Fan R, Hao Y, Du Q, Kang J, Xu M, Li Y. Beneficial Effects of Walnut Oligopeptides on Muscle Loss in Senescence-Accelerated Mouse Prone-8 (SAMP8) Mice: Focusing on Mitochondrial Function. Nutrients. 2022; 14(10):2051. https://doi.org/10.3390/nu14102051
Chicago/Turabian StyleFan, Rui, Yuntao Hao, Qian Du, Jiawei Kang, Meihong Xu, and Yong Li. 2022. "Beneficial Effects of Walnut Oligopeptides on Muscle Loss in Senescence-Accelerated Mouse Prone-8 (SAMP8) Mice: Focusing on Mitochondrial Function" Nutrients 14, no. 10: 2051. https://doi.org/10.3390/nu14102051