
The Effect of Dietary Protein Imbalance during Pregnancy on the Growth, Metabolism and Circulatory Metabolome of Neonatal and Weaned Juvenile Porcine Offspring

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Background
2. Methods
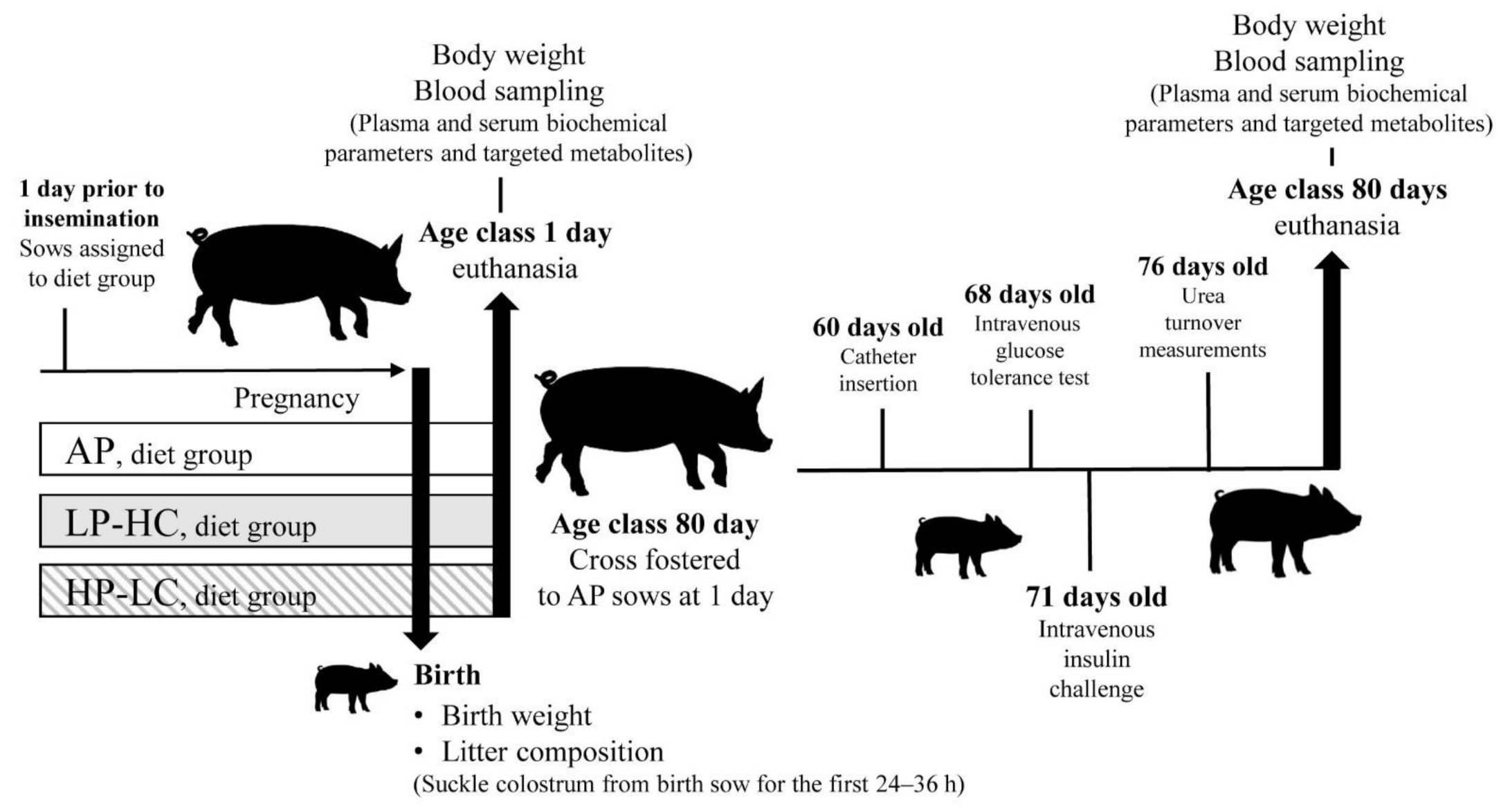
2.1. Experimental Pregnancy Diets
2.2. Animals and Postnatal Offspring Diets
2.3. Blood Sampling
2.4. Plasma and Serum Biochemical Parameters
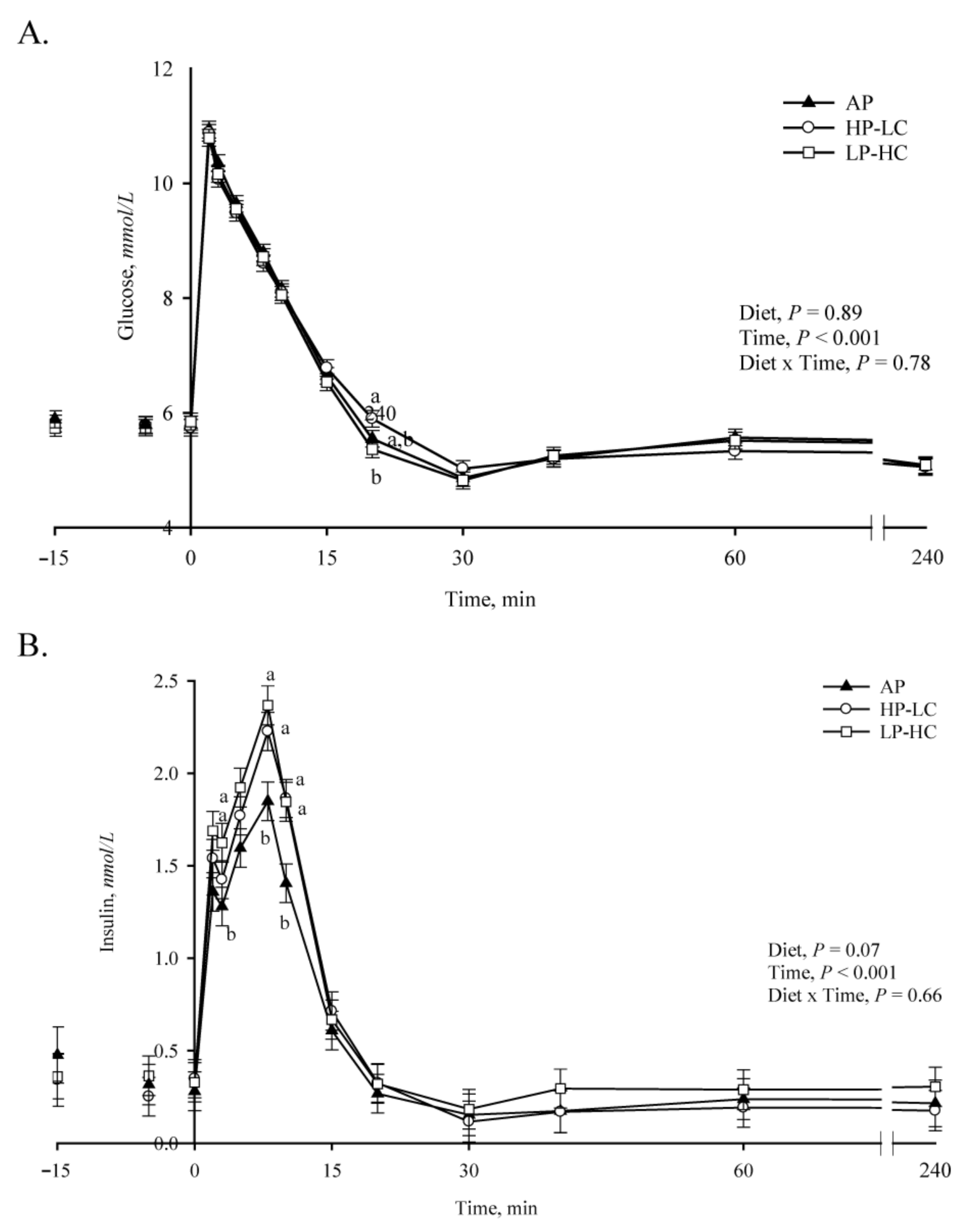
2.5. Intravenous Glucose Tolerance Test/Insulin Challenge and Glucose Turnover Measurements
2.6. Urea Turnover Measurements
2.7. Targeted Metabolomics
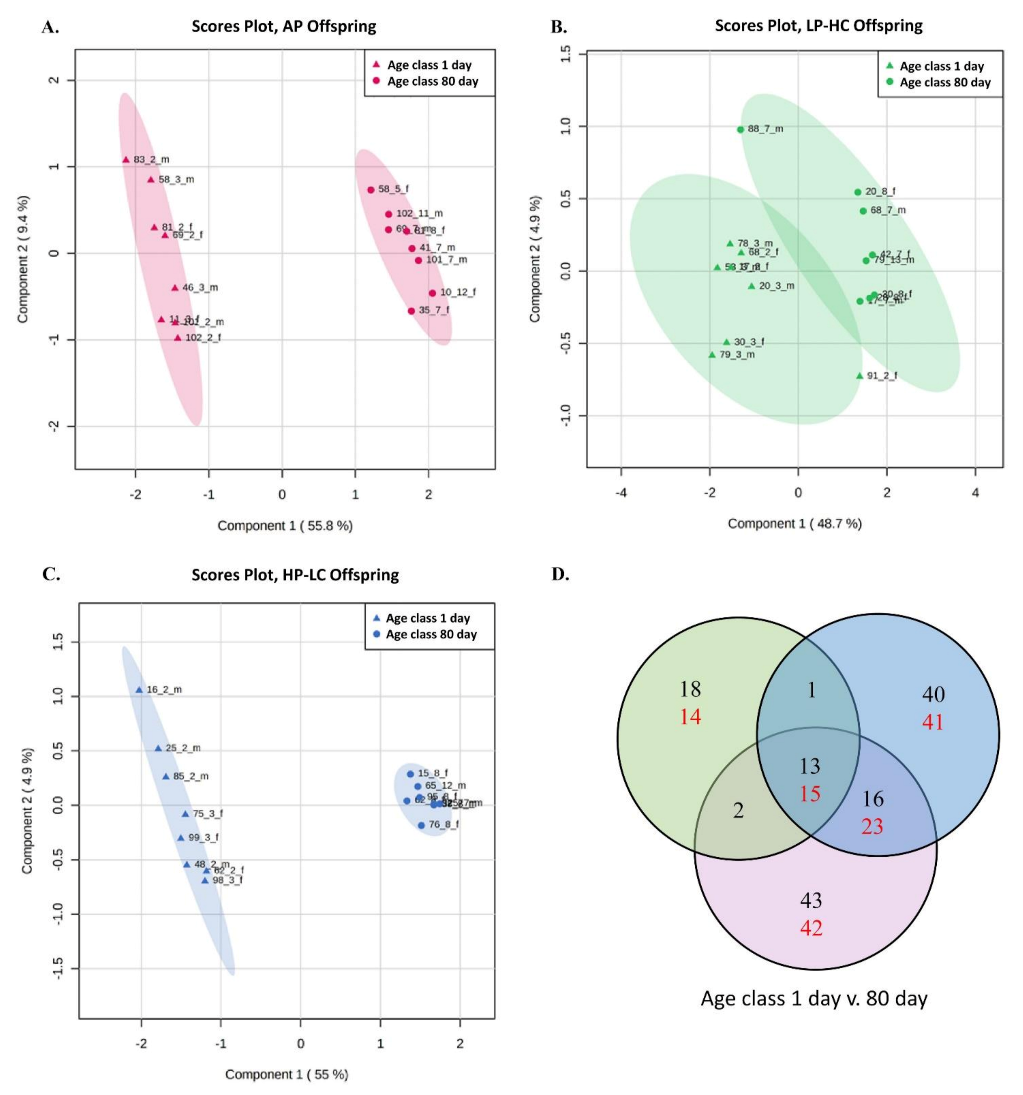
2.8. Data Evaluation and Statistical Analysis
3. Results
3.1. Litter Characteristics and Offspring Body Weight
3.2. Plasma and Serum Biochemical Parameters
3.3. Intravenous Glucose Tolerance, Insulin Challenge and Turnover Studies
3.4. Targeted Metabolite Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.; Eriksson, J.G.; Forsen, T.; Osmond, C. Fetal origins of adult disease: Strength of effects and biological basis. Int. J. Epidemiol. 2002, 31, 1235–1239. [Google Scholar] [CrossRef] [Green Version]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Huffman, S.L. Nutrition in pregnancy and early childhood and associations with obesity in developing countries. Matern. Child Nutr. 2013, 9 (Suppl. S1), 105–119. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. Protein Quality in the First Thousand Days of Life. Food Nutr. Bull. 2016, 37 (Suppl. S1), S14–S21. [Google Scholar] [CrossRef] [Green Version]
- Metzler-Zebeli, B.U.; Lang, I.S.; Gors, S.; Brussow, K.P.; Hennig, U.; Nurnberg, G.; Rehfeldt, C.; Otten, W.; Metges, C.C. High-protein-low-carbohydrate diet during pregnancy alters maternal plasma amino acid concentration and placental amino acid extraction but not fetal plasma amino acids in pigs. Br. J. Nutr. 2012, 108, 2176–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanitz, E.; Otten, W.; Tuchscherer, M.; Grabner, M.; Brussow, K.P.; Rehfeldt, C.; Metges, C.C. High and low proteinratio carbohydrate dietary ratios during gestation alter maternal-fetal cortisol regulation in pigs. PLoS ONE 2012, 7, e52748. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Meng, Q.; Blencowe, M.; Lagarrigue, S.; Xiao, S.; Pan, C.; Wier, J.; Temple, W.C.; Devaskar, S.U.; Lusis, A.J.; et al. Maternal High-Protein and Low-Protein Diets Perturb Hypothalamus and Liver Transcriptome and Metabolic Homeostasis in Adult Mouse Offspring. Front. Genet. 2018, 9, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdge, G.C.; Delange, E.; Dubois, L.; Dunn, R.L.; Hanson, M.A.; Jackson, A.A.; Calder, P.C. Effect of reduced maternal protein intake in pregnancy in the rat on the fatty acid composition of brain, liver, plasma, heart and lung phospholipids of the offspring after weaning. Br. J. Nutr. 2003, 90, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Say, L.; Gulmezoglu, A.M.; Hofmeyr, G.J. Maternal nutrient supplementation for suspected impaired fetal growth. Cochrane Database Syst. Rev. 2003. [Google Scholar] [CrossRef]
- Carter, A.M. Animal models of human pregnancy and placentation: Alternatives to the mouse. Reproduction 2020, 160, R129–R143. [Google Scholar] [CrossRef] [PubMed]
- Sutton, G.M.; Centanni, A.V.; Butler, A.A. Protein malnutrition during pregnancy in C57BL/6J mice results in offspring with altered circadian physiology before obesity. Endocrinology 2010, 151, 1570–1580. [Google Scholar] [CrossRef]
- Mortensen, O.H.; Olsen, H.L.; Frandsen, L.; Nielsen, P.E.; Nielsen, F.C.; Grunnet, N.; Quistorff, B. A maternal low protein diet has pronounced effects on mitochondrial gene expression in offspring liver and skeletal muscle; protective effect of taurine. J. Biomed. Sci. 2010, 17 (Suppl. S1), S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Xiao, X.; Zhang, Q.; Wang, T.; Yu, M.; Xu, J. Maternal Low-Protein Diet Modulates Glucose Metabolism and Hepatic MicroRNAs Expression in the Early Life of Offspring dagger. Nutrients 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Aubert, P.; Oleynikova, E.; Rizvi, H.; Ndjim, M.; Le Berre-Scoul, C.; Grohard, P.A.; Chevalier, J.; Segain, J.P.; Le Drean, G.; Neunlist, M.; et al. Maternal protein restriction induces gastrointestinal dysfunction and enteric nervous system remodeling in rat offspring. FASEB J. 2019, 33, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Twinn, D.S.; Wayman, A.; Ekizoglou, S.; Martin, M.S.; Hales, C.N.; Ozanne, S.E. Maternal protein restriction leads to hyperinsulinemia and reduced insulin-signaling protein expression in 21-mo-old female rat offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R368–R373. [Google Scholar] [CrossRef]
- de Oliveira, J.C.; Gomes, R.M.; Miranda, R.A.; Barella, L.F.; Malta, A.; Martins, I.P.; Franco, C.C.; Pavanello, A.; Torrezan, R.; Natali, M.R.; et al. Protein Restriction During the Last Third of Pregnancy Malprograms the Neuroendocrine Axes to Induce Metabolic Syndrome in Adult Male Rat Offspring. Endocrinology 2016, 157, 1799–1812. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.; Sangild, P.T.; Stoll, B.; Thymann, T.; Buddington, R.; Marini, J.; Olutoye, O.; Shulman, R.J. Translational Advances in Pediatric Nutrition and Gastroenterology: New Insights from Pig Models. Annu. Rev. Anim. Biosci. 2020, 8, 321–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roura, E.; Koopmans, S.J.; Lalles, J.P.; Le Huerou-Luron, I.; de Jager, N.; Schuurman, T.; Val-Laillet, D. Critical review evaluating the pig as a model for human nutritional physiology. Nutr. Res. Rev. 2016, 29, 60–90. [Google Scholar] [CrossRef]
- Chavatte-Palmer, P.; Tarrade, A.; Rousseau-Ralliard, D. Diet before and during Pregnancy and Offspring Health: The Importance of Animal Models and What Can Be Learned from Them. Int. J. Environ. Res. Public Health 2016, 13, 586. [Google Scholar] [CrossRef]
- Puiman, P.; Stoll, B. Animal models to study neonatal nutrition in humans. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 601–606. [Google Scholar] [CrossRef]
- Sciascia, Q.; Daş, G.; Metges, C.C. REVIEW: The pig as a model for humans: Effects of nutritional factors on intestinal function and health. J. Anim. Sci. 2016, 94 (Suppl. S3), 441–452. [Google Scholar] [CrossRef] [Green Version]
- Rehfeldt, C.; Lang, I.S.; Gors, S.; Hennig, U.; Kalbe, C.; Stabenow, B.; Brussow, K.P.; Pfuhl, R.; Bellmann, O.; Nurnberg, G.; et al. Limited and excess dietary protein during gestation affects growth and compositional traits in gilts and impairs offspring fetal growth. J. Anim. Sci. 2011, 89, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Altmann, S.; Murani, E.; Schwerin, M.; Metges, C.C.; Wimmers, K.; Ponsuksili, S. Somatic cytochrome c (CYCS) gene expression and promoter-specific DNA methylation in a porcine model of prenatal exposure to maternal dietary protein excess and restriction. Br. J. Nutr. 2012, 107, 791–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, S.; Murani, E.; Metges, C.C.; Schwerin, M.; Wimmers, K.; Ponsuksili, S. Effect of gestational protein deficiency and excess on hepatic expression of genes related to cell cycle and proliferation in offspring from late gestation to finishing phase in pig. Mol. Biol. Rep. 2012, 39, 7095–7104. [Google Scholar] [CrossRef] [PubMed]
- Sarr, O.; Louveau, I.; Kalbe, C.; Metges, C.C.; Rehfeldt, C.; Gondret, F. Prenatal exposure to maternal low or high protein diets induces modest changes in the adipose tissue proteome of newborn piglets. J. Anim. Sci. 2010, 88, 1626–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oster, M.; Murani, E.; Metges, C.C.; Ponsuksili, S.; Wimmers, K. Transcriptional response of skeletal muscle to a low-protein gestation diet in porcine offspring accumulates in growth- and cell cycle-regulating pathways. Physiol. Genom. 2012, 44, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oster, M.; Murani, E.; Metges, C.C.; Ponsuksili, S.; Wimmers, K. A low protein diet during pregnancy provokes a lasting shift of hepatic expression of genes related to cell cycle throughout ontogenesis in a porcine model. BMC Genom. 2012, 13, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oster, M.; Murani, E.; Metges, C.C.; Ponsuksili, S.; Wimmers, K. A high protein diet during pregnancy affects hepatic gene expression of energy sensing pathways along ontogenesis in a porcine model. PLoS ONE 2011, 6, e21691. [Google Scholar] [CrossRef]
- Oster, M.; Murani, E.; Metges, C.C.; Ponsuksili, S.; Wimmers, K. A gestational high protein diet affects the abundance of muscle transcripts related to cell cycle regulation throughout development in porcine progeny. PLoS ONE 2012, 7, e34519. [Google Scholar]
- Whitfield, P.D.; German, A.J.; Noble, P.J. Metabolomics: An emerging post-genomic tool for nutrition. Br. J. Nutr. 2004, 92, 549–555. [Google Scholar] [CrossRef]
- Bovo, S.; Mazzoni, G.; Galimberti, G.; Calo, D.G.; Fanelli, F.; Mezzullo, M.; Schiavo, G.; Manisi, A.; Trevisi, P.; Bosi, P.; et al. Metabolomics evidences plasma and serum biomarkers differentiating two heavy pig breeds. Animal 2016, 10, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Poupin, N.; Tremblay-Franco, M.; Amiel, A.; Canlet, C.; Remond, D.; Debrauwer, L.; Dardevet, D.; Thiele, I.; Aurich, M.K.; Jourdan, F.; et al. Arterio-venous metabolomics exploration reveals major changes across liver and intestine in the obese Yucatan minipig. Sci. Rep. 2019, 9, 12527. [Google Scholar] [CrossRef] [Green Version]
- Bovo, S.; Mazzoni, G.; Calo, D.G.; Galimberti, G.; Fanelli, F.; Mezzullo, M.; Schiavo, G.; Scotti, E.; Manisi, A.; Samore, A.B.; et al. Deconstructing the pig sex metabolome: Targeted metabolomics in heavy pigs revealed sexual dimorphisms in plasma biomarkers and metabolic pathways. J. Anim. Sci. 2015, 93, 5681–5693. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Bovo, S.; Bosi, P.; Fanelli, F.; Pagotto, U.; Galimberti, G.; Mazzoni, G.; Dall’Olio, S.; Fontanesi, L. Targeted metabolomic profiles of piglet plasma reveal physiological changes over the suckling period. Livest. Sci. 2020, 231, 103890. [Google Scholar] [CrossRef]
- Huang, S.; Li, N.; Liu, C.; Li, T.; Wang, W.; Jiang, L.; Li, Z.; Han, D.; Tao, S.; Wang, J. Characteristics of the gut microbiota colonization, inflammatory profile, and plasma metabolome in intrauterine growth restricted piglets during the first 12 hours after birth. J. Microbiol. 2019, 57, 748–758. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Fiorotto, M.L.; Burrin, D.G.; Pond, W.G.; Nguyen, H.V. Intrauterine growth restriction does not alter response of protein synthesis to feeding in newborn pigs. Am. J. Physiol. 1997, 272 Pt 1, E877–E884. [Google Scholar] [CrossRef]
- Thone-Reineke, C.; Kalk, P.; Dorn, M.; Klaus, S.; Simon, K.; Pfab, T.; Godes, M.; Persson, P.; Unger, T.; Hocher, B. High-protein nutrition during pregnancy and lactation programs blood pressure, food efficiency, and body weight of the offspring in a sex-dependent manner. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1025–R1030. [Google Scholar] [CrossRef] [Green Version]
- Schoknecht, P.A.; Pond, W.G.; Mersmann, H.J.; Maurer, R.R. Protein restriction during pregnancy affects postnatal growth in swine progeny. J. Nutr. 1993, 123, 1818–1825. [Google Scholar] [CrossRef]
- Flachowsky, G.; Pallauf, J.; Pfeffer, E.; Rodehutscord, M.; Schenkel, H.; Staudacher, W.; Susenbeth, A. Empfehlungen zur Energie-und Nährstoffversorgung von Schweinen: 2006; DLG-Verlag: Frankfurt, Germany, 2006. [Google Scholar]
- Rehfeldt, C.; Stabenow, B.; Pfuhl, R.; Block, J.; Nurnberg, G.; Otten, W.; Metges, C.C.; Kalbe, C. Effects of limited and excess protein intakes of pregnant gilts on carcass quality and cellular properties of skeletal muscle and subcutaneous adipose tissue in fattening pigs. J. Anim. Sci. 2012, 90, 184–196. [Google Scholar] [CrossRef]
- Haid, M.; Muschet, C.; Wahl, S.; Romisch-Margl, W.; Prehn, C.; Moller, G.; Adamski, J. Long-Term Stability of Human Plasma Metabolites during Storage at −80 degrees C. J. Proteome Res. 2018, 17, 203–211. [Google Scholar] [CrossRef]
- Metges, C.C.; Lang, I.S.; Hennig, U.; Brussow, K.P.; Kanitz, E.; Tuchscherer, M.; Schneider, F.; Weitzel, J.M.; Steinhoff-Ooster, A.; Sauerwein, H.; et al. Intrauterine growth retarded progeny of pregnant sows fed high protein:low carbohydrate diet is related to metabolic energy deficit. PLoS ONE 2012, 7, e31390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corson, A.M.; Laws, J.; Litten, J.C.; Dodds, P.F.; Lean, I.J.; Clarke, L. Effect of dietary supplementation of different oils during the first or second half of pregnancy on the glucose tolerance of the sow. Animal 2008, 2, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Uken, K.L.; Vogel, L.; Gnott, M.; Gors, S.; Schaff, C.T.; Tuchscherer, A.; Hoeflich, A.; Weitzel, J.M.; Kanitz, E.; Troscher, A.; et al. Effect of maternal supplementation with essential fatty acids and conjugated linoleic acid on metabolic and endocrine development in neonatal calves. J. Dairy Sci. 2021, 104, 7295–7314. [Google Scholar] [CrossRef] [PubMed]
- Otten, W.; Kanitz, E.; Tuchscherer, M.; Grabner, M.; Nurnberg, G.; Bellmann, O.; Hennig, U.; Rehfeldt, C.; Metges, C.C. Effects of low and high protein:carbohydrate ratios in the diet of pregnant gilts on maternal cortisol concentrations and the adrenocortical and sympathoadrenal reactivity in their offspring. J. Anim. Sci. 2013, 91, 2680–2692. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.B.M.; Gors, S.; Derno, M.; Tuchscherer, A.; Wimmers, K.; Zeyner, A.; Kuhla, B. Differences between Holstein dairy cows in renal clearance rate of urea affect milk urea concentration and the relationship between milk urea and urinary nitrogen excretion. Sci. Total Environ. 2021, 755 Pt 2, 143198. [Google Scholar] [CrossRef]
- Zukunft, S.; Sorgenfrei, M.; Prehn, C.; Möller, G.; Adamski, J. Targeted Metabolomics of Dried Blood Spot Extracts. Chromatographia 2013, 76, 1295–1305. [Google Scholar] [CrossRef]
- European Medicines Agency. Guideline on Bioanalytical Method Validation; European Medicines Agency (EMEA/CHMP/EWP/192217/2009 Rev.1 Corr. 2): London, UK, 2011. [Google Scholar]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Kucia, M.; Langhammer, M.; Gors, S.; Albrecht, E.; Hammon, H.M.; Nurnberg, G.; Metges, C.C. High-protein diet during gestation and lactation affects mammary gland mRNA abundance, milk composition and pre-weaning litter growth in mice. Animal 2011, 5, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Vanselow, J.; Kucia, M.; Langhammer, M.; Koczan, D.; Metges, C.C. Maternal high-protein diet during pregnancy, but not during suckling, induced altered expression of an increasing number of hepatic genes in adult mouse offspring. Eur. J. Nutr. 2016, 55, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Metges, C.C.; Gors, S.; Lang, I.S.; Hammon, H.M.; Brussow, K.P.; Weitzel, J.M.; Nurnberg, G.; Rehfeldt, C.; Otten, W. Low and high dietary protein:carbohydrate ratios during pregnancy affect materno-fetal glucose metabolism in pigs. J. Nutr. 2014, 144, 155–163. [Google Scholar] [CrossRef]
- Picone, G.; Zappaterra, M.; Luise, D.; Trimigno, A.; Capozzi, F.; Motta, V.; Davoli, R.; Nanni Costa, L.; Bosi, P.; Trevisi, P. Metabolomics characterization of colostrum in three sow breeds and its influences on piglets’ survival and litter growth rates. J. Anim. Sci. Biotechnol. 2018, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Dividich, J.; Charneca, R.; Thomas, F. Relationship between birth order, birth weight, colostrum intake, acquisition of passive immunity and pre-weaning mortality of piglets. Span. J. Agric. Res. 2017, 15, e0603. [Google Scholar] [CrossRef] [Green Version]
- Loisel, F.; Farmer, C.; Ramaekers, P.; Quesnel, H. Colostrum yield and piglet growth during lactation are related to gilt metabolic and hepatic status prepartum. J. Anim. Sci. 2014, 92, 2931–2941. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.R.; Gottheil, S.K.; Arany, E.J.; Hill, D.J. The effects of low protein during gestation on mouse pancreatic development and beta cell regeneration. Pediatric Res. 2010, 68, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Rehfeldt, C.; Lefaucheur, L.; Block, J.; Stabenow, B.; Pfuhl, R.; Otten, W.; Metges, C.C.; Kalbe, C. Limited and excess protein intake of pregnant gilts differently affects body composition and cellularity of skeletal muscle and subcutaneous adipose tissue of newborn and weanling piglets. Eur. J. Nutr. 2012, 51, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Doring, F.; Luersen, K.; Schmelzer, C.; Hennig, S.; Lang, I.S.; Gors, S.; Rehfeldt, C.; Otten, W.; Metges, C.C. Influence of maternal low protein diet during pregnancy on hepatic gene expression signature in juvenile female porcine offspring. Mol. Nutr. Food Res. 2013, 57, 277–290. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Maternal Diet 1 | p Value, ≤ 2 | ||||
|---|---|---|---|---|---|
| Parameters | Age Class 3 | AP | LP-HC | HP-LC | Age Class |
| Body weight, kg | |||||
| Day 1 | 1 and 80 day | 1.44 ± 0.05 a | 1.25 ± 0.05 b | 1.33 ± 0.05 a,b | - |
| Day 80 | 80 day | 31.9 ± 0.87 a | 30.6 ± 0.78 a,b | 28.1 ± 1.10 b | |
| Metabolites, mmol/L | |||||
| Triglycerides | 1 day | 0.78 ± 0.11 | 0.79 ± 0.11 | 0.95 ± 0.11 c | <0.001 |
| 80 day | 0.42 ± 0.12 | 0.41 ± 0.12 | 0.50 ± 0.12 d | ||
| Urea | 1 day | 5.05 ± 0.37 a,b,c | 4.01 ± 0.36 a,c | 5.61 ± 0.35 b,c | <0.001 |
| 80 day | 2.22 ± 0.40 d | 2.20 ± 0.42 d | 2.17 ± 0.41 d | ||
| Hormones | |||||
| Glucagon, pg/mL | 1 day | 243 ± 35 c | 198 ± 35 | 296 ± 34 c | <0.001 |
| 80 day | 68 ± 41 d | 50 ± 40 | 52 ± 42 d | ||
| Insulin, µU/mL | 1 day | 6.97 ± 1.60 | 3.31 ± 1.57 | 9.02 ± 1.52 | 0.86 |
| 80 day | 6.41 ± 1.82 | 6.92 ± 1.87 | 6.69 ± 1.85 | ||
| Glucagon:Insulin 4 | 1 day | 3.17 ± 0.41 c | 3.63 ± 0.40 c | 2.42 ± 0.39 c | <0.001 |
| 80 day | 0.73 ± 0.53 d | 0.24 ± 0.50 d | 0.50 ± 0.53 d | ||
| Comparison | KEGG Pathway | Metabolites | |||
|---|---|---|---|---|---|
| Identified | Total 1 | p Value | FDR | ||
| HP-LC | Aminoacyl-tRNA biosynthesis | 10 | 48 | <0.001 | <0.001 |
| Arginine biosynthesis | 4 | 14 | <0.001 | <0.001 | |
| Histidine metabolism | 4 | 16 | <0.001 | <0.001 | |
| beta-Alanine metabolism | 4 | 21 | <0.001 | 0.01 | |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 2 | 4 | <0.001 | 0.02 | |
| AP | Aminoacyl-tRNA biosynthesis | 8 | 48 | <0.001 | <0.001 |
| Arginine biosynthesis | 3 | 14 | <0.001 | 0.03 | |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 2 | 4 | <0.001 | 0.03 | |
| Histidine metabolism | 3 | 16 | <0.001 | 0.03 | |
| beta-Alanine metabolism | 3 | 21 | <0.001 | 0.05 | |
| Shared by both | Aminoacyl-tRNA biosynthesis | 6 | 48 | <0.001 | <0.001 |
| AP and HP-LC | Histidine metabolism | 3 | 16 | <0.001 | 0.02 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 2 | 4 | <0.001 | 0.02 | |
| beta-Alanine metabolism | 3 | 21 | <0.001 | 0.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciascia, Q.L.; Prehn, C.; Adamski, J.; Daş, G.; Lang, I.S.; Otten, W.; Görs, S.; Metges, C.C. The Effect of Dietary Protein Imbalance during Pregnancy on the Growth, Metabolism and Circulatory Metabolome of Neonatal and Weaned Juvenile Porcine Offspring. Nutrients 2021, 13, 3286. https://doi.org/10.3390/nu13093286
Sciascia QL, Prehn C, Adamski J, Daş G, Lang IS, Otten W, Görs S, Metges CC. The Effect of Dietary Protein Imbalance during Pregnancy on the Growth, Metabolism and Circulatory Metabolome of Neonatal and Weaned Juvenile Porcine Offspring. Nutrients. 2021; 13(9):3286. https://doi.org/10.3390/nu13093286
Chicago/Turabian StyleSciascia, Quentin L., Cornelia Prehn, Jerzy Adamski, Gürbüz Daş, Iris S. Lang, Winfried Otten, Solvig Görs, and Cornelia C. Metges. 2021. "The Effect of Dietary Protein Imbalance during Pregnancy on the Growth, Metabolism and Circulatory Metabolome of Neonatal and Weaned Juvenile Porcine Offspring" Nutrients 13, no. 9: 3286. https://doi.org/10.3390/nu13093286