
Adjuvant Effect of Orally Applied Preparations Containing Non-Digestible Polysaccharides on Influenza Vaccination in Healthy Seniors: A Double-Blind, Randomised, Controlled Pilot Trial

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
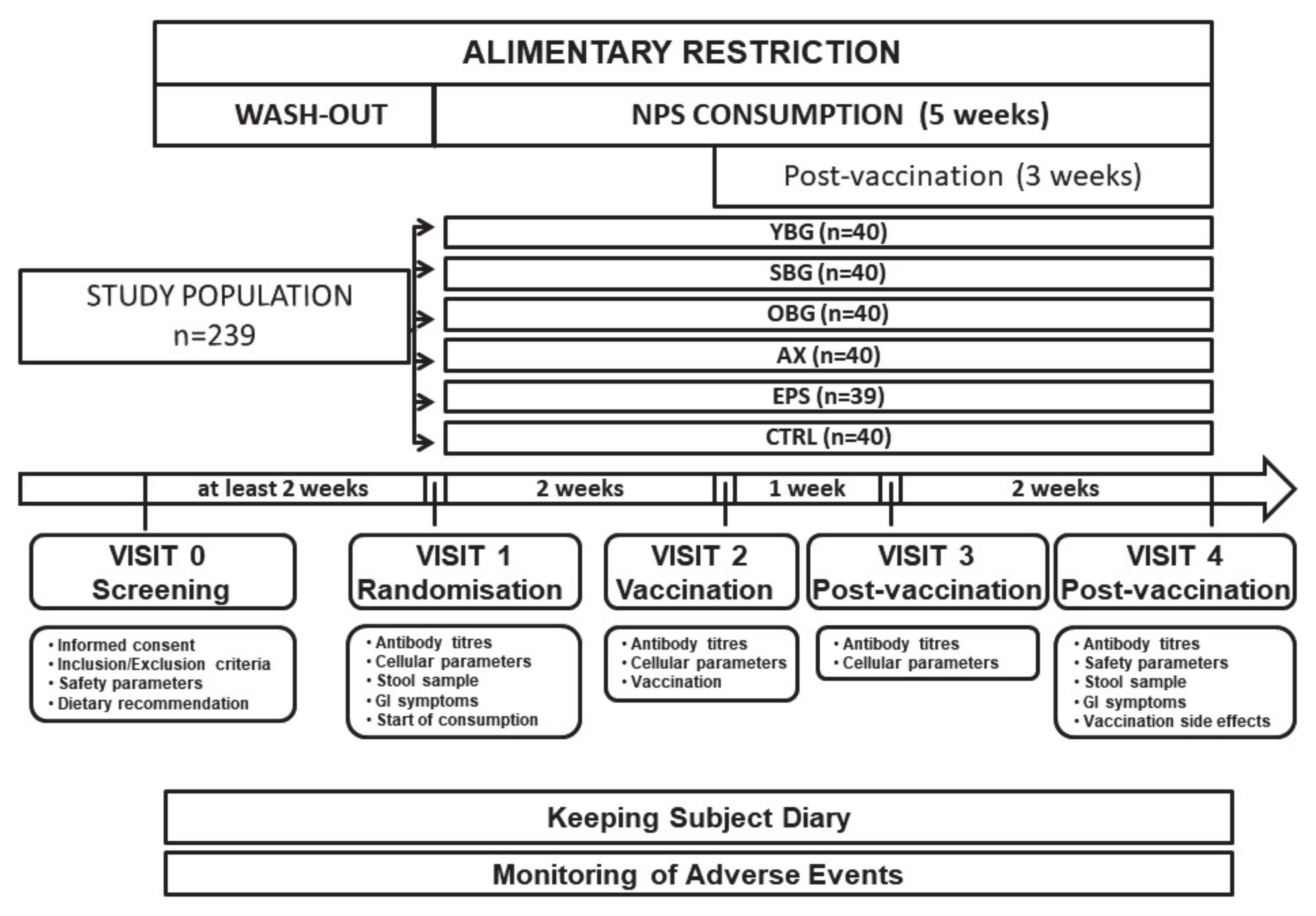
2.1. Study Population
2.2. Study Products
2.3. Study Design
2.4. Compliance Assessment
2.5. Influenza Vaccine
2.6. Acquisition of Blood
2.7. Antibody Titres
2.8. Respiratory Tract Infections
2.8.1. Common Cold (Upper Respiratory Tract Infection)
2.8.2. Influenza or Influenza-Like Illness
2.8.3. Acute Bronchitis and Pneumonia (Lower Respiratory Tract Infection)
2.9. Cellular Immunity
2.10. Microbiota Analysis
2.11. Faecal pH and Short-Chain Fatty Acids
2.12. Gastrointestinal Symptoms, Stool Consistency and Stool Frequency
2.13. Diet
2.14. Safety Laboratory Parameters
2.15. Vaccination Side Effects
2.16. Statistical Analysis
3. Results
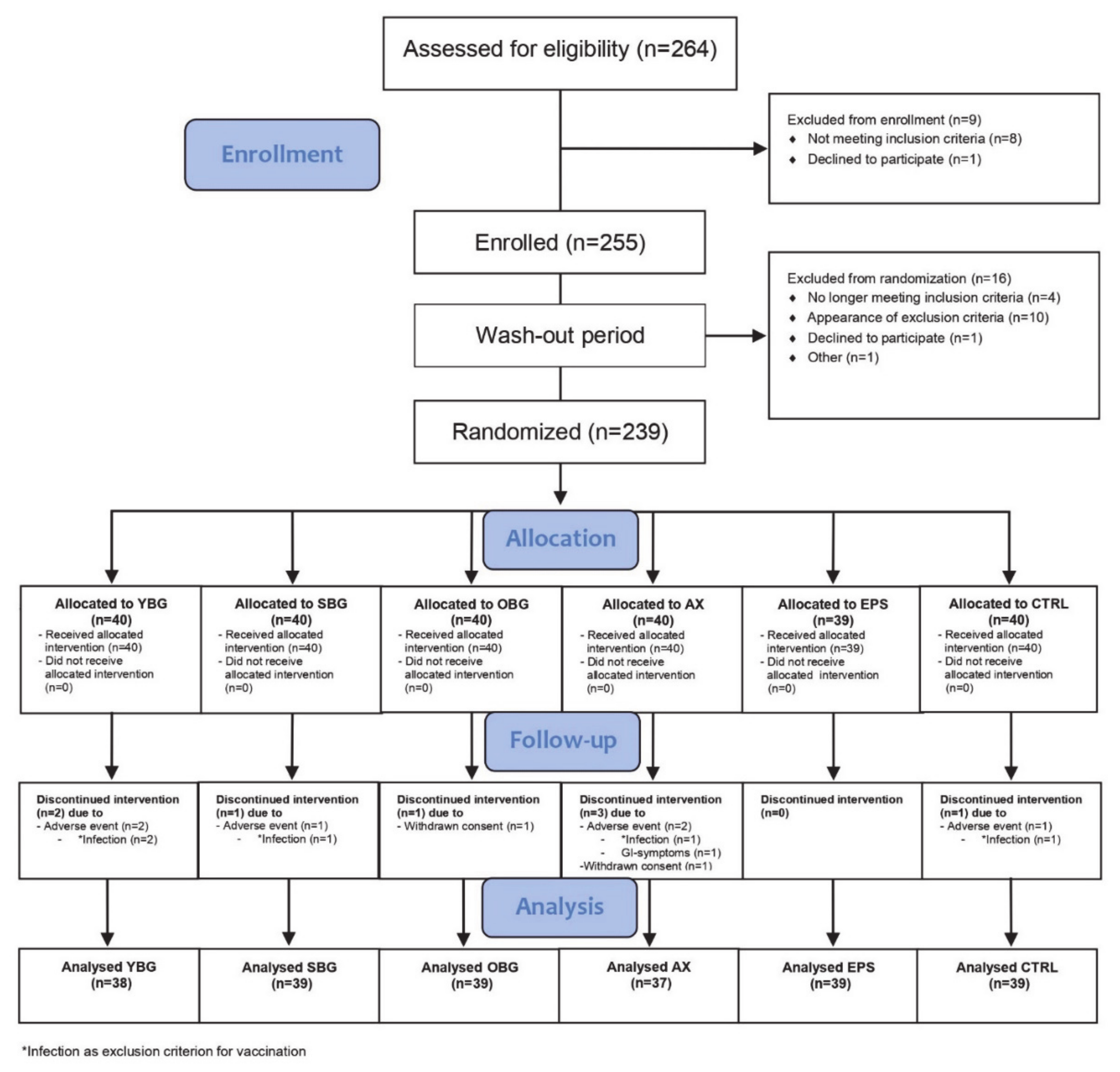
3.1. Study Subjects
3.2. Antibody Titres
3.2.1. HI Antibodies
3.2.2. Micro-Neutralization Antibodies
3.3. Common Cold Incidence
3.4. Cytokine Production
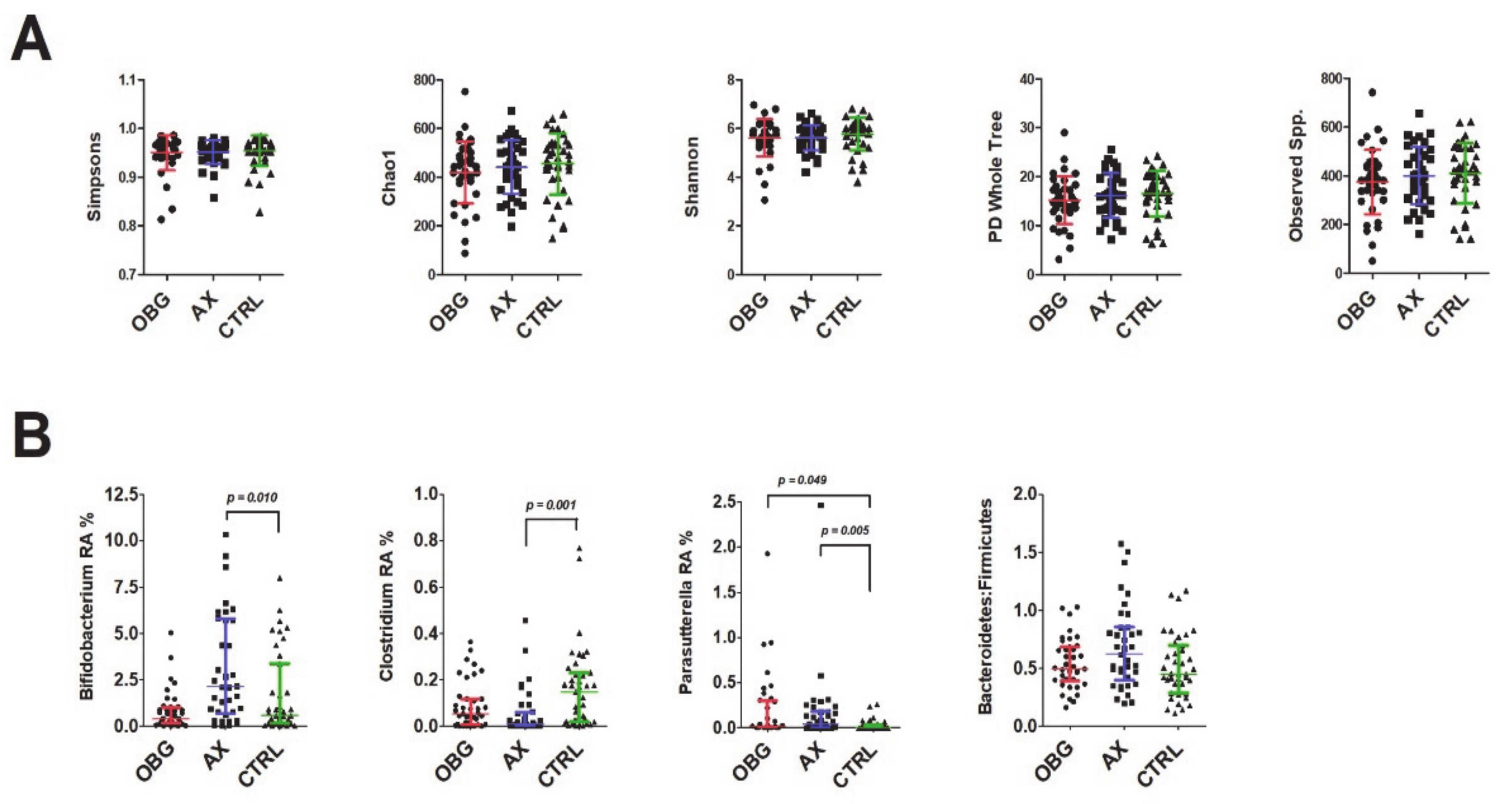
3.5. Microbiota Composition
3.6. Short-Chain Fatty Acids and Faecal pH
3.7. Dietary Intake
3.8. Gastrointestinal Symptom Scores, Stool Frequency and Stool Consistency
3.9. Laboratory Safety Parameters and Adverse Events
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gruver, A.L.; Hudson, L.L.; Sempowski, G.D. Immunosenescence of ageing. J. Pathol. 2007, 211, 144–156. [Google Scholar] [CrossRef]
- Bowdish, D.; Loukov, D.; Naidoo, A. Immunosenescence: Implications for vaccination programs in the elderly. Vaccine: Dev. Ther. 2015, 5, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, K.; Viboud, C.; Simonsen, L. Antibody response to influenza vaccination in the elderly: A quantitative review. Vaccine 2006, 24, 1159–1169. [Google Scholar] [CrossRef]
- Akatsu, H. Exploring the effect of probiotics, prebiotics, and postbiotics in strengthening immune activity in the elderly. Vaccines 2021, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Albers, R.; Antoine, J.-M.; Bourdet-Sicard, R.; Calder, P.C.; Gleeson, M.; LeSourd, B.; Samartín, S.; Sanderson, I.R.; Van Loo, J.; Dias, F.W.V.; et al. Markers to measure immunomodulation in human nutrition intervention studies. Br. J. Nutr. 2005, 94, 452–481. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority Panel on Dietetic Products, Nutrition and Allergies. Guidance on the scientific requirements for health claims related to gut and immune function. EFSA J. 2011, 9, 1984. [Google Scholar] [CrossRef]
- European Food Safety Authority Panel on Nutrition, Novel Foods and Food Allergens. Guidance on the scientific requirements for health claims related to the immune system, the gastrointestinal tract and defence against pathogenic microorganisms. EFSA J. 2016, 14, 4369. [Google Scholar] [CrossRef]
- Lomax, A.R.; Calder, P. Prebiotics, immune function, infection and inflammation: A review of the evidence. Br. J. Nutr. 2008, 101, 633–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokryazdan, P.; Jahromi, M.F.; Navidshad, B.; Liang, J.B. Effects of prebiotics on immune system and cytokine expression. Med. Microbiol. Immunol. 2016, 206, 1–9. [Google Scholar] [CrossRef]
- Petrovsky, N.; Cooper, P.D. Carbohydrate-based immune adjuvants. Expert Rev. Vaccines 2011, 10, 523–537. [Google Scholar] [CrossRef]
- Hald, S.; Schioldan, A.G.; Moore, M.E.; Dige, A.; Lærke, H.N.; Agnholt, J.; Knudsen, K.E.B.; Hermansen, K.; Marco, M.L.; Gregersen, S.; et al. Effects of arabinoxylan and resistant starch on intestinal microbiota and short-chain fatty acids in subjects with metabolic syndrome: A randomised crossover study. PLoS ONE 2016, 11, e0159223. [Google Scholar] [CrossRef]
- Cianci, R.; Franza, L.; Massaro, M.G.; Borriello, R.; De Vito, F.; Gambassi, G. The interplay between immunosenescence and microbiota in the efficacy of vaccines. Vaccines 2020, 8, 636. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Wang, F. Polysaccharides: Candidates of promising vaccine adjuvants. Drug Discov. Ther. 2015, 9, 88–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Liu, X.; Qian, T.; Sun, G.; Guo, Y.; Chang, F.; Zhou, S.; Sun, X. Antitumor and immunomodulatory activity of arabinoxylans: A major constituent of wheat bran. Int. J. Biol. Macromol. 2011, 48, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Agrawal, S. Activation of human monocyte-derived dendritic cells in vitro by the biological response modifier Arabinoxylan rice bran (MGN-3/BIOBRAN). Int. J. Immunopathol. Pharmacol. 2011, 24, 941–948. [Google Scholar] [CrossRef]
- Mendis, M.; Leclerc, E.; Simsek, S. Arabinoxylans, gut microbiota and immunity. Carbohydr. Polym. 2016, 139, 159–166. [Google Scholar] [CrossRef]
- Choi, J.Y.; Paik, D.J.; Kwon, D.Y.; Park, Y. Dietary supplementation with rice bran fermented with Lentinus edodes increases interferon-γ activity without causing adverse effects: A randomized, double-blind, placebo-controlled, parallel-group study. Nutr. J. 2014, 13, 35. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, K.C.; Breslin, W.L.; Davidson, T.; Adams, A.; McFarlin, B.K. Baker′s yeast β-glucan supplementation increases monocytes and cytokines post-exercise: Implications for infection risk? Br. J. Nutr. 2012, 109, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vashishta, A.; Saraswat-Ohri, S.; Vetvickova, J. Immunological effects of yeast- and mushroom-derived β-glucans. J. Med. Food 2008, 11, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vannucci, L.; Sima, P. β-glucan as a new tool in vaccine development. Scand. J. Immunol. 2019, 91, e12833. [Google Scholar] [CrossRef] [PubMed]
- Xiu, L.; Zhang, H.; Hu, Z.; Liang, Y.; Guo, S.; Yang, M.; Du, R.; Wang, X. Immunostimulatory activity of exopolysaccharides from probiotic Lactobacillus casei WXD030 strain as a novel adjuvant in vitro and in vivo. Food Agric. Immunol. 2018, 29, 1086–1105. [Google Scholar] [CrossRef] [Green Version]
- Vinderola, G.; Perdigón, G.; Duarte, J.; Farnworth, E.; Matar, C. Effects of the oral administration of the exopolysaccharide produced by Lactobacillus kefiranofaciens on the gut mucosal immunity. Cytokine 2006, 36, 254–260. [Google Scholar] [CrossRef]
- Jin, Y.; Li, P.; Wang, F. β-glucans as potential immunoadjuvants: A review on the adjuvanticity, structure-activity relationship and receptor recognition properties. Vaccine 2018, 36, 5235–5244. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Carpenter, K.C.; Davidson, T.; McFarlin, M.A. Baker′s yeast beta glucan supplementation increases salivary IgA and decreases cold/flu symptomatic days after intense exercise. J. Diet. Suppl. 2013, 10, 171–183. [Google Scholar] [CrossRef]
- Fuller, R.; Butt, H.; Noakes, P.; Kenyon, J.; Yam, T.S.; Calder, P. Influence of yeast-derived 1,3/1,6 glucopolysaccharide on circulating cytokines and chemokines with respect to upper respiratory tract infections. Nutrition 2012, 28, 665–669. [Google Scholar] [CrossRef]
- Dharsono, T.; Rudnicka, K.; Wilhelm, M.; Schoen, C. Effects of yeast (1,3)-(1,6)-beta-glucan on severity of upper respiratory tract infections: A double-blind, randomized, placebo-controlled study in healthy subjects. J. Am. Coll. Nutr. 2018, 38, 40–50. [Google Scholar] [CrossRef]
- Jin, X.; Liu, X.; Ding, J.; Zhang, L.; Yang, Y.; Wang, X.; Yang, Y.; Liu, M. Lentinan improved the efficacy of vaccine against Trichinella spiralis in an NLRP3 dependent manner. PLoS Neglected Trop. Dis. 2020, 14, e0008632. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, M.; Xu, L.; Yang, X.; Chang, Y.; Zhu, Y. Effect of edible fungal polysaccharides on improving influenza vaccine protection in mice. Food Agric. Immunol. 2017, 28, 981–992. [Google Scholar] [CrossRef]
- Saulnier, L. Types and functionality of polysaccharides in cereal grains. In Food Chemistry, Function and Analysis No. 6 Cereal Grain-Based Functional Foods: Carbohydrate and Phytochemical Components; Beta, T., Camire, M.E., Eds.; The Royal Society of Chemistry: London, UK, 2019; pp. 54–84. [Google Scholar]
- Akhtar, M.; Tariq, A.F.; Awais, M.M.; Iqbal, Z.; Muhammad, F.; Shahid, M.; Hiszczynska-Sawicka, E. Studies on wheat bran Arabinoxylan for its immunostimulatory and protective effects against avian coccidiosis. Carbohydr. Polym. 2012, 90, 333–339. [Google Scholar] [CrossRef]
- Patel, S.; Majumder, A.; Goyal, A. Potentials of Exopolysaccharides from Lactic Acid Bacteria. Indian J. Microbiol. 2011, 52, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Shen, Q.; Wu, R.; Li, P. Structural analysis and mucosal immune regulation of exopolysaccharide fraction from Bifidobacterium animalis RH. Food Agric. Immunol. 2017, 28, 1226–1241. [Google Scholar] [CrossRef] [Green Version]
- Ryan, P.M.; Ross, R.; Fitzgerald, G.F.; Caplice, N.M.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2014, 6, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.M.; Stolte, E.H.; London, L.E.E.; Wells, J.M.; Long, S.L.; Joyce, S.A.; Gahan, C.G.M.; Fitzgerald, G.F.; Ross, R.P.; Caplice, N.M.; et al. Lactobacillus mucosae DPC 6426 as a bile-modifying and immunomodulatory microbe. BMC Microbiol. 2019, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y. Non-Digestible Polysaccharides to Support the Intestinal Immune Barrier: In Vitro Models to Unravel Molecular Mechanisms. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 18 December 2017. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Thomas, J.; Chandler, J.; Cumpstom, M.; Page, M.J.; Welch, V.A. Assessing Risk of Bias in a Randomized Trial. Available online: http://handbook.cochrane.org. (accessed on 29 May 2012).
- Wichmann, B.A.; Hill, I.D. Algorithm AS 183: An efficient and portable pseudo-random number generator. J. R. Stat. Soc. Ser. C Appl. Stat. 1982, 31, 188. [Google Scholar] [CrossRef]
- Morisky, D.E.; Green, L.W.; Levine, D.M. Concurrent and predictive validity of a self-reported measure of medication adherence. Med. Care 1986, 24, 67–74. [Google Scholar] [CrossRef]
- WHO. Recommended composition of influenza virus vaccines for use in the 2012–2013 northern hemisphere influenza season. Wkly. Epidemiol. Rec. 2012, 87, 83–95. [Google Scholar]
- Committee for Proprietary Medicinal Products. Harmonisation of Requirements for Influenza Vaccines; CPMP/BWP/214/96; The European Agency for the Evaluation of Medical Products (EMEA): London, UK, 1996. [Google Scholar]
- Nauta, J. Statistics In Clinical Vaccine Trials, 1st ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Predy, G.N.; Goel, V.; Lovlin, R.; Donner, A.; Stitt, L.; Basu, T.K. Efficacy of an extract of North American ginseng containing poly-furanosyl-pyranosyl-saccharides for preventing upper respiratory tract infections: A randomized controlled trial. Can. Med. Assoc. J. 2005, 173, 1043–1048. [Google Scholar] [CrossRef] [Green Version]
- Jackson, G.G.; Dowling, H.F.; Spiesman, I.G.; Boand, A.V. Transmission of the common cold to volunteers under controlled conditions. A.M.A. Arch. Intern. Med. 1958, 101, 267–278. [Google Scholar] [CrossRef]
- Fouhy, F.; Deane, J.; Rea, M.; O′Sullivan, O.; Ross, R.; O’Callaghan, G.; Plant, B.J.; Stanton, C. The effects of freezing on faecal microbiota as determined using MiSeq sequencing and culture-based investigations. PLoS ONE 2015, 10, e0119355. [Google Scholar] [CrossRef]
- De Weirdt, R.; Possemiers, S.; Vermeulen, G.; Moerdijk-Poortvliet, T.C.; Boschker, H.T.; Verstraete, W.; Van De Wiele, T. Human faecal microbiota display variable patterns of glycerol metabolism. FEMS Microbiol. Ecol. 2010, 74, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, C.Y.; Morris, J.; Whorwell, P. The irritable bowel severity scoring system: A simple method of monitoring irritable bowel syndrome and its progress. Aliment. Pharmacol. Ther. 1997, 11, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Riegler, G.; Esposito, I. Bristol scale stool form. A still valid help in medical practice and clinical research. Tech. Coloproctol. 2001, 5, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Falomir, Z.; Arregui, M.; Madueño, F.; Corella, D.; Coltell, O. Automation of food questionnaires in medical studies: A state-of-the-art review and future prospects. Comput. Biol. Med. 2012, 42, 964–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boge, T.; Rémigy, M.; Vaudaine, S.; Tanguy, J.; Bourdet-Sicard, R.; van der Werf, S. A probiotic fermented dairy drink improves antibody response to influenza vaccination in the elderly in two randomised controlled trials. Vaccine 2009, 27, 5677–5684. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Kim, Y.; Lee, H.; Kim, J.; Han, B.K.; Go, E.; Kwon, S.; Kang, J.-G.; You, S.; Kwon, J. Carrot pomace polysaccharide (CPP) improves influenza vaccine efficacy in immunosuppressed mice via dendritic cell activation. Nutrition 2020, 12, 2740. [Google Scholar] [CrossRef]
- Ghoneum, M.; Agrawal, S. MGN-3/Biobran enhances generation of cytotoxic CD8+ T cells via upregulation of DEC-205 expression on dendritic cells. Int. J. Immunopathol. Pharmacol. 2014, 27, 523–530. [Google Scholar] [CrossRef]
- Ghoneum, M.; Matsuura, M. Augmentation of macrophage phagocytosis by modified arabinoxylan rice bran (MGN-3/Biobran). Int. J. Immunopathol. Pharmacol. 2004, 17, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Govers, C.; Wichers, H.; Mes, J.J. Macrophages treated with non-digestible polysaccharides reveal a transcriptionally unique phenotype. J. Funct. Foods 2017, 36, 280–289. [Google Scholar] [CrossRef]
- Govers, C.; Tang, Y.; Stolte, E.H.; Wichers, H.J.; Mes, J.J. Wheat-derived arabinoxylans reduced M2-macrophage functional activity, but enhanced monocyte-recruitment capacity. Food Funct. 2020, 11, 7073–7083. [Google Scholar] [CrossRef]
- Yeh, T.-L.; Shih, P.-C.; Liu, S.-J.; Lin, C.-H.; Liu, J.-M.; Lei, W.-T.; Lin, C.-Y. The influence of prebiotic or probiotic supplementation on antibody titers after influenza vaccination: A systematic review and meta-analysis of randomized controlled trials. Drug Des. Dev. Ther. 2018, 12, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Trieu, M.-C.; Zhou, F.; Lartey, S.L.; Sridhar, S.; Mjaaland, S.; Cox, R.J. Augmented CD4+ T-cell and humoral responses after repeated annual influenza vaccination with the same vaccine component A/H1N1pdm09 over 5 years. Vaccines 2018, 3, 37. [Google Scholar] [CrossRef]
- Belongia, E.A.; Skowronski, D.M.; McLean, H.Q.; Chambers, C.; Sundaram, M.E.; De Serres, G. Repeated annual influenza vaccination and vaccine effectiveness: Review of evidence. Expert Rev. Vaccines 2017, 16, 723–736. [Google Scholar] [CrossRef] [Green Version]
- WHO. Recommended viruses for influenza vaccines for use in the 2010–2011 northern hemisphere influenza season. Wkly. Epidemiol. Rec. 2010, 85, 81–92. [Google Scholar]
- WHO. Recommended composition of influenza virus vaccines for use in the 2011–2012 northern hemisphere influenza season. Wkly. Epidemiol. Rec. 2011, 86, 81–91. [Google Scholar]
- Veguilla, V.; Hancock, K.; Schiffer, J.; Gargiullo, P.; Lu, X.; Aranio, D.; Branch, A.; Dong, L.; Holiday, C.; Liu, F.; et al. Sensitivity and specificity of serologic assays for detection of human infection with 2009 pandemic H1N1 virus in U.S. Populations. J. Clin. Microbiol. 2011, 49, 2210–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaullier, J.-M.; Sleboda, J.; Ofjord, E.S.; Ulvestad, E.; Nurminiemi, M.; Moe, C.; Albrektsen, T.; Gudmundsen, O. Supplementation with a soluble beta-glucan exported from Shiitake medicinal mushroom, Lentinus edodes (Berk.) Singer Mycelium: A Crossover, placebo-controlled study in healthy elderly. Int. J. Med. Mushrooms 2011, 13, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Ichihashi, K.; Fujii, T.; Omura, K.; Zhu, X.; Anazawa, M.; Tazawa, K. Oral administration of hydrolyzed rice bran prevents the common cold syndrome in the elderly based on its immunomodulatory action. BioFactors 2004, 21, 185–187. [Google Scholar] [CrossRef]
- Mocchegiani, E.; Malavolta, M. NK and NKT cell functions in immunosenescence. Aging Cell 2004, 3, 177–184. [Google Scholar] [CrossRef]
- Elsaid, A.; Shaheen, M.; Ghoneum, M. Biobran/MGN-3, an arabinoxylan rice bran, enhances NK cell activity in geriatric subjects: A randomized, double-blind, placebo-controlled clinical trial. Exp. Ther. Med. 2018, 15, 2313–2320. [Google Scholar] [CrossRef] [Green Version]
- Kak, G.; Raza, M.; Tiwari, B.K. Interferon-gamma (IFN-γ): Exploring its implications in infectious diseases. Biomol. Concepts 2018, 9, 64–79. [Google Scholar] [CrossRef]
- McFarlin, B.K.; Venable, A.S.; Carpenter, K.C.; Henning, A.L.; Ogenstad, S.; McFarlin, B.K.; Venable, A.S.; Carpenter, K.C.; Henning, A.L.; Ogenstad, S. Oral supplementation with Baker’s yeast beta glucan is associated with altered monocytes, t cells and cytokines following a bout of strenuous exercise. Front. Physiol. 2017, 8, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leentjens, J.; Quintin, J.; Gerretsen, J.; Kox, M.; Pickkers, P.; Netea, M.G. The effects of orally administered beta-glucan on innate immune responses in humans, a randomized open-label intervention pilot-study. PLoS ONE 2014, 9, e108794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, D.J.; Pulendran, B. The potential of the microbiota to influence vaccine responses. J. Leukoc. Biol. 2017, 103, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Santamarina, A.; Lamas, A.; Mondragón, A.D.C.; Cardelle-Cobas, A.; Regal, P.; Rodriguez-Avila, J.A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Probiotic effects against virus infections: New weapons for an old war. Foods 2021, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.S.; Lærke, H.N.; Theil, P.K.; Sørensen, J.F.; Saarinen, M.; Forssten, S.; Knudsen, K.E.B. Diets high in resistant starch and arabinoxylan modulate digestion processes and SCFA pool size in the large intestine and faecal microbial composition in pigs. Br. J. Nutr. 2014, 112, 1837–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cloetens, L.; Broekaert, W.F.; Delaedt, Y.; Ollevier, F.; Courtin, C.; Delcour, J.; Rutgeerts, P.; Verbeke, K. Tolerance of arabinoxylan-oligosaccharides and their prebiotic activity in healthy subjects: A randomised, placebo-controlled cross-over study. Br. J. Nutr. 2009, 103, 703–713. [Google Scholar] [CrossRef]
- Yasui, H.; Kiyoshima, J.; Hori, T.; Shida, K. Protection against influenza virus infection of mice fed bifidobacterium breve YIT4064. Clin. Diagn. Lab. Immunol. 1999, 6, 186–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namba, K.; Hatano, M.; Yaeshima, T.; Takase, M.; Suzuki, K. Effects of Bifidobacterium longum BB536 administration on influenza infection, influenza vaccine antibody titer, and cell-mediated immunity in the elderly. Biosci. Biotechnol. Biochem. 2010, 74, 939–945. [Google Scholar] [CrossRef] [Green Version]
- De Vrese, M.; Winkler, P.; Rautenberg, P.; Harder, T.; Noah, C.; Laue, C.; Ott, S.; Hampe, C.; Schreiber, S.; Heller, K.; et al. Effect of Lactobacillus gasseri PA 16/8, Bifidobacterium longum SP 07/3, B. bifidum MF 20/5 on common cold episodes: A double blind, randomized, controlled trial. Clin. Nutr. 2005, 24, 481–491. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; Bird, A.; Conlon, M.A.; Williams, B.A.; Kang, S.; McSweeney, C.; Zhang, D.; Bryden, W.L.; Gidley, M.J.; Topping, D.L. An arabinoxylan-rich fraction from wheat enhances caecal fermentation and protects colonocyte DNA against diet-induced damage in pigs. Br. J. Nutr. 2011, 107, 1274–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, T.; Kong, J.Y.; Stothard, P.; Willing, B.P. Defining the role of Parasutterella, a previously uncharacterized member of the core gut microbiota. ISME J. 2019, 13, 1520–1534. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Liu, L.; Meng, G.; Zhu, S.; Zhou, R.; Jiang, W. IL-18 maintains the homeostasis of mucosal immune system via inflammasome-independent but microbiota-dependent manner. Sci. Bull. 2021. [Google Scholar] [CrossRef]
- Li, Y.; Elmén, L.; Segota, I.; Xian, Y.; Tinoco, R.; Feng, Y.; Fujita, Y.; Muñoz, R.R.S.; Schmaltz, R.; Bradley, L.M.; et al. Prebiotic-induced anti-tumor immunity attenuates tumor growth. Cell Rep. 2020, 30, 1753–1766.e6. [Google Scholar] [CrossRef] [Green Version]
- Christodoulides, S.; Dimidi, E.; Fragkos, K.; Farmer, A.D.; Whelan, K.; Scott, S.M. Systematic review with meta-analysis: Effect of fibre supplementation on chronic idiopathic constipation in adults. Aliment. Pharmacol. Ther. 2016, 44, 103–116. [Google Scholar] [CrossRef]
- Calame, W.; Weseler, A.R.; Viebke, C.; Flynn, C.; Siemensma, A.D. Gum arabic establishes prebiotic functionality in healthy human volunteers in a dose-dependent manner. Br. J. Nutr. 2008, 100, 1269–1275. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Total (n = 239) | YBG (n = 40) | SBG (n = 40) | OBG (n = 40) | AX (n = 40) | EPS (n = 39) | CTRL (n = 40) | p-Value | |
|---|---|---|---|---|---|---|---|---|
| Female/male (n) | 118/121 | 17/23 | 19/21 | 23/17 | 22/18 | 23/16 | 14/26 | 0.212 |
| Age (years) | 67.9 [63.1; 71.4] | 68.2 [62.1; 71.2] | 67.3 [60.2; 70.0] | 68.0 [63.0; 71.3] | 67.5 [63.7; 71.3] | 66.2 [62.2; 72.3] | 69.5 [65.0; 73.3] | 0.271 |
| BMI (kg/m2) | 27.0 ± 3.6 | 27.2 ± 3.9 | 27.0 ± 3.1 | 26.9 ± 3.5 | 26.7 ± 4.0 | 27.3 ± 3.7 | 27.3 ± 3.5 | 0.969 |
| YBG | SBG | OBG | AX | EPS | CTRL | p | ||
|---|---|---|---|---|---|---|---|---|
| GMT (Log2(HI Titre/10)) | ||||||||
| H1N1 | V1 | 1.00 [0.00; 3.00] | 1.75 [0.00; 3.00] | 0.50 [0.00; 2.50] | 0.88 [−0.44; 2.00] | 1.50 [0.00; 2.50] | 1.38 [0.13; 2.94] | 0.557 |
| V4 | 2.88 [1.00; 4.00] | 2.50 [1.00; 3.50] | 2.50 [0.50;3.50] | 2.50 [1.88; 3.63] | 2.00 [1.00; 3.00] | 3.00 [1.50; 4.00] | 0.548 | |
| MLFI | 1.00 [0.00; 1.50] | 0.75 [0.00; 1.50] * | 1.00 [0.25; 2.25] | 2.00 [1.00; 3.00] # | 1.00 [0.50; 2.00] | 1.25 [0.50; 2.00] | 0.010 | |
| H3N2 | V1 | 0.00 [−1.00; 0.69] | 0.00 [−1.00; 0.50] | 0.00 [−1.00; 0.50] | −0.38 [−1.00; 0.50] | −0.50 [−1.00; 0.00] | −0.25 [−1.00; 0.44] | 0.266 |
| V4 | 1.00 [−1.00; 2.00] | 1.00 [0.00; 1.50] | 1.00 [0.25; 1.50] | 1.00 [0.00; 2.00] | 0.50 [0.00; 1.25] | 1.00 [0.50; 1.75] | 0.382 | |
| MLFI | 0.50 [0.00; 1.50] | 0.75 [0.00; 1.25] | 1.00 [0.50; 2.00] | 1.00 [0.38; 1.75] | 1.00 [0.00; 1.75] | 1.00 [0.50; 2.00] | 0.198 | |
| Influenza B | V1 | −0.50 [−1.00; 2.00] | −0.50 [−1.00; 0.50] | 0.00 [−1.00; 0.44] | −0.25 [−1.00; 1.00] | 0.00 [−1.00; 1.00] | 0.13 [−1.00; 1.88] | 0.741 |
| V4 | 1.00 [1.19; 2.06] | 1.00 [0.00; 2.00] | 1.00 [0.00; 1.50] | 1.00 [0.00; 2.13] | 1.00 [−0.25; 2.00] | 1.00 [0.00; 2.00] | 0.786 | |
| MLFI | 1.00 [0.00; 2.06] | 1.00 [0.00; 2.00] | 0.75 [0.00; 1.75] | 1.00 [0.00; 2.00] | 0.75 [0.00; 2.00] | 1.00 [0.00; 1.50] | 0.683 | |
| YBG | SBG | OBG | AX | EPS | CTRL | p | ||
|---|---|---|---|---|---|---|---|---|
| Seroprotection rate (%) | ||||||||
| H1N1 | V1 | 42.5 | 50.0 | 27.5 | 27.5 | 38.5 | 42.5 | 0.230 |
| V4 | 60.5 | 59.0 | 59.0 | 75.7 | 61.5 | 66.7 | 0.633 | |
| V4-V1 | 18.4 | 10.3 | 30.8 | 48.7 # | 23.1 | 25.6 | 0.005 | |
| H3N2 | V1 | 5.0 | 7.5 | 2.5 | 7.5 | 0.0 | 2.5 | 0.476 |
| V4 | 26.3 | 17.9 | 19.7 | 32.4 | 15.4 | 20.5 | 0.468 | |
| V4-V1 | 21.0 | 12.8 | 15.4 | 24.3 | 15.4 | 17.9 | 0.802 | |
| Influenza B | V1 | 15.0 | 7.5 | 7.5 | 10.0 | 10.3 | 25.0 | 0.148 |
| V4 | 36.8 | 28.2 | 17.9 | 37.8 | 25.6 | 35.9 | 0.343 | |
| V4-V1 | 26.3 | 23.1 | 10.3 | 27.0 | 15.4 | 10.3 | 0.180 | |
| Seroconversion rate (%) | ||||||||
| H1N1 | V1 to V4 | 15.8 | 17.9 | 28.2 | 43.2 | 17.9 | 23.1 | 0.056 |
| H3N2 | V1 to V4 | 13.2 | 7.7 | 10.3 | 10.8 | 10.3 | 17.9 | 0.799 |
| Influenza B | V1 to V4 | 21.1 | 17.9 | 7.7 | 21.6 | 17.9 | 15.4 | 0.618 |
| YBG | SBG | OBG | AX | EPS | CTRL | p | ||
|---|---|---|---|---|---|---|---|---|
| GMT (Log2(Titre/10)) | ||||||||
| H1N1 | V1 | 0.49 [−0.46; 3.88] | 2.51 [−0.51; 4.50] | 0.75 [−0.51; 2.18] | 0.49 [−0.51; 2.38] | 1.00 [−0.51; 2.51] | 0.49 [−0.51; 2.00] | 0.408 |
| V4 | 4.50 [2.88; 6.00] | 4.50 [3.00; 6.13] | 4.50 [2.74; 6.00] | 5.50 [3.75; 6.00] | 4.50 [2.00; 5.50] | 5.00 [3.00; 6.50] | 0.437 | |
| MLFI | 2.00 [0.93; 4.26] # | 1.50 [0.51; 3.63] * | 3.51 [1.74; 5.01] | 3.77 [2.50; 5.51] | 2.09 [1.00; 4.50] | 3.50 [1.50; 5.51] | 0.013 | |
| H3N2 | V1 | 1.25 [0.49; 3.88] | 1.49 [0.00; 2.51] | 0.49 [−0.51; 1.49] | 0.49 [−0.51; 2.00] | 0.49 [−0.51; 1.49] | 0.49 [0.00; 2.00] | 0.031 |
| V4 | 4.25 [1.87; 5.63] | 4.50 [2.51; 5.50] | 4.00 [1.49; 6.00] | 4.50 [2.76; 6.00] | 3.50 [2.51; 5.00] | 4.00 [3.00; 5.50] | 0.586 | |
| MLFI | 1.88 [0.51; 3.50] | 2.02 [0.88; 4.01] | 3.01 [1.49; 5.50] | 3.00 [1.00; 4.76] | 3.03 [1.51; 4.50] | 3.03 [1.51; 5.02] | 0.112 | |
| Influenza B | V1 | 1.00 [0.13; 1.50] | 1.00 [0.50; 2.00] | 0.50 [0.00; 1.50] | 1.00 [0.50; 2.00] | 0.50 [0.00; 2.00] | 1.00 [0.50; 2.00] | 0.486 |
| V4 | 3.00 [2.50; 4.50] | 3.25 [2.00; 4.55] | 3.00 [1.50; 4.00] | 3.50 [1.90; 5.15] | 3.50 [2.00; 5.30] | 3.80 [2.00; 4.50] | 0.609 | |
| MLFI | 2.03 [0.95; 3.51] | 1.52 [0.87; 3.01] | 1.81 [1.00; 3.01] | 2.03 [1.49; 3.37] | 2.26 [1.10; 3.52] | 2.00 [0.51; 3.01] | 0.745 | |
| YBG | SBG | OBG | AX | EPS | CTRL | p | ||
|---|---|---|---|---|---|---|---|---|
| Seroprotection rate (%) | ||||||||
| H1N1 | V1 | 40.0 | 53.8 | 25.0 | 35.0 | 33.3 | 30.0 | 0.130 |
| V4 | 86.8 | 84.2 | 84.6 | 89.2 | 79.5 | 89.7 | 0.813 | |
| V4-V1 | 44.7 | 30.8 * | 59.0 | 59.5 | 46.2 | 61.5 | 0.050 | |
| H3N2 | V1 | 37.5 | 38.5 | 15.0 | 32.5 | 20.5 | 27.5 | 0.126 |
| V4 | 76.3 | 86.8 | 74.4 | 81.1 | 84.6 | 97.4 | 0.082 | |
| V4-V1 | 39.5 | 48.7 | 59.0 | 48.6 | 64.1 | 71.8 | 0.053 | |
| Influenza B | V1 | 22.5 | 30.8 | 20.0 | 32.5 | 30.8 | 30.0 | 0.753 |
| V4 | 84.2 | 78.9 | 74.4 | 75.7 | 79.5 | 76.9 | 0.927 | |
| V4-V1 | 60.5 | 50.0 | 53.8 | 43.2 | 48.7 | 46.1 | 0.726 | |
| Seroconversion rate (%) | ||||||||
| H1N1 | V1 to V4 | 52.6 | 39.5 # | 69.2 | 75.7 | 51.3 | 61.5 | 0.019 |
| H3N2 | V1 to V4 | 42.1 | 52.6 | 64.1 | 56.8 | 69.2 | 71.8 | 0.076 |
| Influenza B | V1 to V4 | 60.5 | 47.4 | 48.7 | 56.8 | 53.8 | 51.3 | 0.863 |
| n | YBG | SBG | OBG | AX | EPS | CTRL | p Chi-Square Test | p Fisher Exact Test AX vs. CTRL |
|---|---|---|---|---|---|---|---|---|
| Cold acc. to Predy et al. [43] | 3 | 1 | 3 | 1 | 5 | 5 | 0.317 | 0.113 |
| Cold acc. to Jackson et al. [44] | 7 | 3 | 5 | 1 | 7 | 8 | 0.141 | 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laue, C.; Stevens, Y.; van Erp, M.; Papazova, E.; Soeth, E.; Pannenbeckers, A.; Stolte, E.; Böhm, R.; Gall, S.L.; Falourd, X.; et al. Adjuvant Effect of Orally Applied Preparations Containing Non-Digestible Polysaccharides on Influenza Vaccination in Healthy Seniors: A Double-Blind, Randomised, Controlled Pilot Trial. Nutrients 2021, 13, 2683. https://doi.org/10.3390/nu13082683
Laue C, Stevens Y, van Erp M, Papazova E, Soeth E, Pannenbeckers A, Stolte E, Böhm R, Gall SL, Falourd X, et al. Adjuvant Effect of Orally Applied Preparations Containing Non-Digestible Polysaccharides on Influenza Vaccination in Healthy Seniors: A Double-Blind, Randomised, Controlled Pilot Trial. Nutrients. 2021; 13(8):2683. https://doi.org/10.3390/nu13082683
Chicago/Turabian StyleLaue, Christiane, Yala Stevens, Monique van Erp, Ekaterina Papazova, Edlyn Soeth, Angelika Pannenbeckers, Ellen Stolte, Ruwen Böhm, Sophie Le Gall, Xavier Falourd, and et al. 2021. "Adjuvant Effect of Orally Applied Preparations Containing Non-Digestible Polysaccharides on Influenza Vaccination in Healthy Seniors: A Double-Blind, Randomised, Controlled Pilot Trial" Nutrients 13, no. 8: 2683. https://doi.org/10.3390/nu13082683