
Creatine Supplementation for Patients with Inflammatory Bowel Diseases: A Scientific Rationale for a Clinical Trial


{kind=link}
{kind=link}
{kind=link}
Abstract
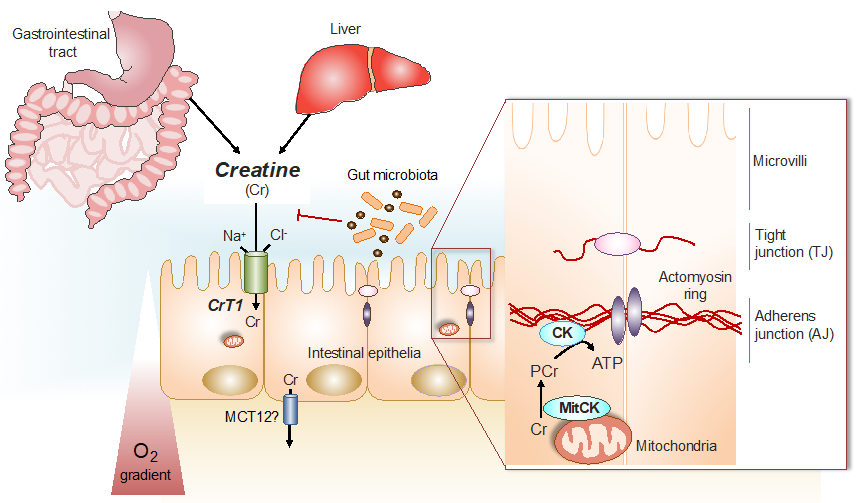
:1. Introduction
1.1. The Phospho-Creatine Creatine Kinase System in Intestinal Epithelial Cells
1.2. Creatine for Cytoprotection against Ischemia, Hypoxia, Oxidative Stress and Acidosis
1.3. Creatine Stimulates Mitochondrial Respiration and Serves as an Anti-Apoptotic Effector
1.4. Creatine as Anti-Inflammatory, Nociceptive and Immune Modulatory Compound
1.5. Creatine Affords Anti-Depressant Effects
2. Scientific Rationale Specifically for Intestinal Tissue
2.1. HIF Controls Creatine Kinase (CK) Expression and CK Together with Creatine Are Involved in the Energetics of Mucosal Barrier Regulation
2.2. Creatine Supplementation Regulates the Energy Balance of Intestinal Epithelial Cells, Epithelial Integrity and Barrier Function
2.3. Creatine Supplementation Maintains Intestinal Epithelial Energy Homeostasis and Protects against Colitis in Animal Models
2.4. Creatine-Loading Preserves Intestinal Barrier Function during Intestine Organ Preservation by Static Cold Storage
2.5. A Genetic Screen with Mice Susceptible for Colitis Reveals a Link to Creatine Metabolism
2.6. Involvement of AMPK Activation to Restore Adherence Junction Assembly in Intestinal Epithelium
2.7. Creatine Supplementation in One Single Case of Crohn’s Disease Improved Both Symptomatic and Endoscopic Characteristics of Ulcerative Colitis
3. Proposal, Methodology and Clinical End-Points
4. Anticipated Outcome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jin, L.; Li, L.; Hu, C.; Paez-Cortez, J.; Bi, Y.; Macoritto, M.; Cao, S.; Tian, Y. Integrative Analysis of Transcriptomic and Proteomic Profiling in Inflammatory Bowel Disease Colon Biopsies. Inflamm. Bowel Dis. 2019, 25, 1906–1918. [Google Scholar] [CrossRef]
- Wang, Q.; Mi, S.; Yu, Z.; Li, Q.; Lei, J. Opening a Window on Attention: Adjuvant Therapies for Inflammatory Bowel Disease. Can. J. Gastroenterol. Hepatol. 2020, 7397523. [Google Scholar] [CrossRef]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [Green Version]
- Kreider, R.B.; Stout, J.R. Creatine in Health and Disease. Nutrients 2021, 13, 447. [Google Scholar] [CrossRef]
- Tarnopolsky, M.A. Clinical use of creatine in neuromuscular and neurometabolic disorders. Subcell Biochem. 2007, 46, 183–204. [Google Scholar] [CrossRef]
- Kreider, R.B.; Kalman, D.S.; Antonio, J.; Ziegenfuss, T.N.; Wildman, R.; Collins, R.; Candow, D.G.; Kleiner, S.M.; Almada, A.L.; Lopez, H.L. International Society of Sports Nutrition position stand: Safety and efficacy of creatine supplementation in exercise, sport, and medicine. J. Int. Soc. Sports Nutr. 2017, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Harris, R. Creatine: A miserable life without it. Amino Acids. 2016, 48, 1739–1750. [Google Scholar] [CrossRef]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H.M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuation energy demands: The phosphocreatine circuit for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessman, S.P.; Carpenter, C.L. The creatine-creatine phosphate energy shuttle. Annu. Rev. Biochem. 1985, 54, 831–862. [Google Scholar] [CrossRef]
- Wallimann, T.; Hemmer, W. Creatine kinase in non-muscle tissues and cells. Mol. Cell Biochem. 1994, 133–134, 193–220. [Google Scholar] [CrossRef]
- Peral, M.J.; García-Delgado, M.; Calonge, M.L.; Durán, J.M.; De La Horra, M.C.; Wallimann, T.; Speer, O.; Ilundáin, A. Human, rat and chicken small intestinal Na+-Cl−-creatine transporter: Functional, molecular characterization and localization. J. Physiol. 2002, 545, 133–144. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol Rev. 2000, 80, 1107–1121. [Google Scholar] [CrossRef] [PubMed]
- Okuma, D.O.; Aponte-Collazo, L.J.; Dewar, B.J.; Cox, N.J.; East, M.P.; Tech, K.; McDonald, I.M.; Tikunov, A.P.; Holmuhamedov, E.; Macdonald, J.M.; et al. Lyn regulates creatine uptake in an imatinib-resistant CML cell line. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129507. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.H.T.; Lee, J.S.; Murphy, E.M.; Gerich, M.E.; Dran, R.; Glover, L.E.; Abdulla, Z.I.; Skelton, M.R.; Colgan, S.P. Creatine Transporter, Reduced in Colon Tissues from Patients With Inflammatory Bowel Diseases, Regulates Energy Balance in In-testinal Epithelial Cells, Epithelial Integrity, and Barrier Function. Gastroenterology 2020, 159, 984–998. [Google Scholar] [CrossRef]
- Takahashi, M.; Kishimoto, H.; Shirasaka, Y.; Inoue, K. Functional characterization of monocarboxylate transporter 12 (SLC16A12/MCT12) as a facilitative creatine transporter. Drug Metab. Pharmacokinet. 2020, 35, 281–287. [Google Scholar] [CrossRef]
- Glover, L.E.; Bowers, B.E.; Saeedi, B.; Ehrentraut, S.F.; Campbell, E.L.; Bayless, A.J.; Dobrinskikh, E.; Kendrick, A.A.; Kelly, C.J.; Burgess, A.; et al. Control of creatine metabolism by HIF is an endogenous mechanism of barrier regulation in colitis. Proc. Natl. Acad. Sci. USA 2013, 110, 19820–19825. [Google Scholar] [CrossRef] [Green Version]
- Kitzenberg, D.; Colgan, S.P.; Glover, L.E. Creatine kinase in ischemic and inflammatory disorders. Clin. Transl. Med. 2016, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Brewer, G.J.; Wallimann, T. Protective effect of the energy precursor creatine against toxicity of glutamate and beta-amyloid in rat hippocampal neurons. J. Neurochem. 2000, 74, 1968–1978. [Google Scholar] [CrossRef]
- Adcock, K.H.; Nedelcu, J.; Loenneker, T.; Martin, E.; Wallimann, T.; Wagner, B.P. Neuroprotection of creatine supplementation in neonatal rats with transient cerebral hypoxia-ischemia. Dev. Neurosci. 2002, 24, 382–388. [Google Scholar] [CrossRef]
- Prass, K.; Royl, G.; Lindauer, U.; Freyer, D.; Megow, D.; Dirnagl, U.; Stöckler-Ipsiroglu, G.; Wallimann, T.; Priller, J. Improved reperfustion and neuroprotection by creatine in a mouse model of stroke. J. Cereb. Blood Flow Metab. 2007, 27, 452–459. [Google Scholar] [CrossRef]
- Balestrino, M.; Sarocchi, M.; Adriano, E.; Spallarossa, P. Potential of creatine or phosphocreatine supplementation in cere-brovascular disease and in ischemic heart disease. Amino Acids 2016, 48, 1955–1967. [Google Scholar] [CrossRef]
- Vavricka, S.; Ruiz, P.A.; Scharl, S.; Biedermann, L.; Scharl, M.; de Vallière, C.; Lundby, C.; Wenger, R.H.; Held, L.; Merz, T.M.; et al. Protocol for a prospective, controlled, observational study to evaluate the influence of hypoxia on healthy volunteers and patients with inflammatory bowel disease: The Altitude IBD-Study. BMJ Open 2017, 7, e013477. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, E.; Guescini, M.; Calcabrini, C.; Vallorani, L.; Diaz, A.R.; Fimognari, C.; Canonico, B.; Luchetti, F.; Papa, S.; Battistelli, M.; et al. Creatine prevents the structural and functional damage to mitochondria in myogenic oxidatively stressed C2C12 cells and restores their differentiation capacity. Oxid. Med. Cell Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Hemati, F.; Rahmani, A.; Asadollahi, K.; Soleimannejad, K.; Khalighi, Z. Effects of Complimentary Creatine Monohydrate and Physical Training on Inflammatory and Endothelial Dysfunction Markers Among Heart Failure Patients. Asian J. Sports Med. 2016, 7, e28578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santacuz, L.; Arciniegas, A.J.L.; Darrabie, M.; Mantilla, J.G.; Baron, R.M.; Bowles, D.E.; Mishra, R.; Jacobs, D.O. Hypoxia decreases creatine uptake in cardiomyocytes, while creatine supplementation enhances HIF activation. Physiol. Rep. 2017, 5, e13382. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Wang, R.X.; Alexeev, E.E.; Lanis, J.M.; Battista, K.D.; Glover, L.E.; Colgan, S.P. Hypoxanthine is a checkpoint stress metabolite in colonic epithelial energy modulation and barrier function. J. Biol. Chem. 2018, 293, 6039–6051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestili, P.; Martinelli, C.; Colombo, E.; Barbieri, E.; Potenza, L.; Sartini, S.; Fimognari, C. Creatine as an antioxidant. Amino Acids 2011, 40, 1385–1396. [Google Scholar] [CrossRef]
- Ellery, S.J.; Dickinson, H.; McKenzie, M.; Walker, D.W. Dietary interventions designed to protect the perinatal brain from hypoxic-ischemic encephalopathy—Creatine prophylaxis and the need for multi-organ protection. Neurochem. Int. 2016, 95, 15–23. [Google Scholar] [CrossRef]
- Whittington, H.J.; Ostrowski, P.J.; McAndrew, D.J.; Cao, F.; Shaw, A.; Eykyn, T.R.; Lake, H.; Tyler, J.; Schneider, J.E.; Neubauer, S.; et al. Over-expression of mitochondrial creatine kinase in the murine heart improves functional recovery and protects against injury following ischemia-reperfusion. Cardiovasc. Res. 2018, 114, 858–869. [Google Scholar] [CrossRef]
- Whittington, H.J.; McAndrew, D.J.; Cross, R.L.; Neubauer, S.; Lygate, C.A. Protective effect of creatine elevation against ischaemia reperfusion injury is retained in the presence of co-morbidities and during cardioplegia. PLoS ONE 2016, 11, e0146429. [Google Scholar] [CrossRef] [Green Version]
- Giatromanolaki, A.; Sivridis, E.; Maltezos, E.; Papazoglou, D.; Simopoulos, C.; Gatter, K.C.; Harris, A.L.; Koukourakis, M.I. Hypoxia inducible factor 1alpha and 2alpha overexpression in inflammatory bowel disease. J. Clin. Pathol. 2003, 56, 209–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, C.T.; Dzus, A.L.; Colgan, S.P. Autocrine regulation of epithelial permeability by hypoxia: Role for polarized release of tumor necrosis factor alpha. Gastroenterology 1998, 114, 657–668. [Google Scholar] [CrossRef]
- Pedoto, A.; Nandi, J.; Oler, A.; Camporesi, E.M.; Hakim, T.S.; Levine, R.A. Role of Nitric Oxide in Acidosis-Induced Intestinal Injury in Anesthetized Rats. J. Lab. Clin. Med. 2001, 138, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Sironi, C.; Bodega, F.; Zocchi, L.; Porta, C. Effects of creatine treatment on jejunial phenotypes in a rat model of acidosis. Antioxidants 2019, 8, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, L.; Nicolay, K.; Wieringa, B.; Saks, V.; Wallimann, T. Direct evidence for the control of mitochondrial respiration by mitochondrial creatine kinase in oxidative muscle cells in situ. J. Biol. Chem. 2000, 275, 6937–6944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Gorman, E.; Beutner, G.; Dolder, M.; Koretsky, A.P.; Brdiczka, D.; Wallimann, T. The role of creatine kinase and creatine in inhibition of mitochondrial permeability transition. FEBS Lett. 1997, 414, 253–257. [Google Scholar] [CrossRef]
- Dolder, M.; Walzel, B.; Speer, O.; Schlattner, U.; Wallimann, T. Inhibition of the mitochondrial permeability transition by creatine kinase substrates. Requirement for micro-compartmentation. J. Biol. Chem. 2003, 278, 17760–17766. [Google Scholar] [CrossRef] [Green Version]
- Caretti, A.; Bianciardi, P.; Sala, G.; Terruzzi, C.; Lucchina, F.; Samaja, M. Supplementation of creatine and ribose prevents apoptosis in ischemic cardiomyocytes. Cell Physiol. Biochem. 2010, 26, 831–838. [Google Scholar] [CrossRef]
- Sun, Z.; Lan, X.; Ahsan, A.; Xi, Y.; Liu, S.; Zhang, Z.; Chu, P.; Song, Y.; Piao, F.; Peng, J.; et al. Phosphocreatine protects against LPS-induced human umbilical vein endothelial cell apoptosis by regulating mitochondrial oxidative phosphorylation. Apoptosis 2016, 21, 283–297. [Google Scholar] [CrossRef]
- Busanello, E.N.B.; Marques, A.C.; Lander, N.; de Oliveira, D.N.; Catharino, R.R.; Oliveira, H.C.F.; Vercesi, A.E. Pravastatin chronic treatment sensitizes hypercholesterolemic mice muscle to mitochondrial permeability transition: Protection by creatine or coenzyme Q10. Front. Pharmacol. 2017, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Speer, O.; Bäck, N.; Buerklen, T.; Brdiczka, D.; Koretsky, A.; Wallimann, T.; Eriksson, O. Octameric mitochondrial creatine kinase induces and stabilizes contact sites between the inner and outer mitochondrial membrane. Biochem. J. 2005, 385, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Wallimann, T.; Riek, U.; Möddel, M. Intradialytic creatine supplementation: A scientific rationale for improving the health and quality of life of dialysis patients. Med. Hypotheses 2017, 99, 1–14. [Google Scholar] [CrossRef]
- Qiu, W.; Wu, B.; Wang, X.; Buchanan, M.E.; Regueiro, M.D.; Hartman, D.J.; Schoen, R.E.; Yu, J.; Zhang, L. PUMA-mediated intestinal epithelial apoptosis contributes to ulcerative colitis in humans and mice. J. Clin. Investig. 2011, 121, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.; Martini, E.; Nadine Wittkopf, N.; Amann, K.; Weigmann, B.; Neumann, H.; Waldner, M.; Hedrick, S.M.; Tenzer, S.; Neurath, M.F.; et al. Caspase-8 regulates TNF-alpha induced epithelial necroptosis and terminal ileitis. Nature 2011, 477, 335–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassit, R.A.; Pinheiro, C.H.; Vitzel, K.F.; Sproesser, A.J.; Silveira, L.R.; Curi, R. Effect of short-term creatine supplementation on markers of skeletal muscle damage after strenuous contractile activity. Eur. J. Appl. Physiol. 2010, 108, 945–955. [Google Scholar] [CrossRef]
- Izurieta Munoz, H.; Gonzales, E.B.; Sumien, N. Effects of creatine supplementation on nociception in young male and female mice. Pharmacol. Rep. 2018, 70, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Gajula, P.; Quigley, E.M. Overlapping irritable bowel syndrome and inflammatory bowel disease. Minerva Gastrolenterologica Diabetol. 2019, 65, 107–115. [Google Scholar] [CrossRef]
- Kazak, L.; Cohen, P. Creatine metabolism: Energy homeostasis, immunity and cancer biology. Nat. Rev. Endocrinol. 2020, 16, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Taii, A.; Tachikawa, M.; Ohta, Y.; Hosoya, K.I.; Terasaki, T. Determination of Intrinsic Creatine Transporter (Slc6a8) Activity and Creatine Transport Function of Leukocytes in Rats. Biol. Pharm. Bull. 2020, 43, 474–479. [Google Scholar] [CrossRef]
- Venter, G.; Polling, S.; Pluk, H.; Venselaar, H.; Wijers, M.; Willemse, M.; Fransen, J.A.M.; Wieringa, B. Submembranous recruitment of creatine kinase B supports formation of dynamic actin-based protrusions of macrophages and relies on its C-terminal flexible loop. Eur. J. Cell Biol. 2015, 94, 114–127. [Google Scholar] [CrossRef]
- Ji, L.; Zhao, X.; Zhang, B.; Kang, L.; Song, W.; Zhao, B.; Xie, W.; Chen, L.; Hu, X. Slc6a8-Mediated Creatine Uptake and Accumulation Reprogram Macrophage Polarization via Regulating Cytokine Responses. Immunity 2019, 51, 272–284.e7. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Wang, X.; Gao, X.; Liu, X. Regulation of T Cell Development and Activation by Creatine Kinase B. PLoS ONE 2009, 4, e5000. [Google Scholar] [CrossRef]
- Di Biase, S.; Ma, X.; Wang, X.; Yu, J.; Wang, Y.C.; Smith, D.J.; Zhou, Y.; Li, Z.; Kim, Y.J.; Clarke, N.; et al. Creatine uptake regulates CD8 T cell antitumor immunity. J. Exp. Med. 2019, 216, 2869–2882. [Google Scholar] [CrossRef] [Green Version]
- Bredahl, E.C.; Eckerson, J.M.; Tracy, S.M.; McDonald, T.L.; Drescher, K.M. The Role of Creatine in the Development and Activation of Immune Responses. Nutrients 2021, 13, 751. [Google Scholar] [CrossRef] [PubMed]
- Andres, D.; Ducray, A.D.; Schlattner, U.; Wallimann, T.; Widmer, H.R. Functions and effects of creatine in the central nervous system. Brain Res. Bull. 2008, 76, 329–343. [Google Scholar] [CrossRef]
- Kious, B.M.; Kondo, D.G.; Renshaw, P.E. Creatine for the treatment of depression. Biomolecules 2019, 9, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, D.G.; Forrest, L.N.; Shi, X.; Sung, Y.H.; Hellem, T.L.; Huber, R.S.; Renshaw, P.F. Creatine target engagement with brain bioenergetics: A dose-range phosphorus-31-magnetic resonance spectroscopy study of adolescent females with SSRI-resistant depression. Amino Acids 2016, 48, 1941–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kious, B.M.; Sabic, H.; Sung, Y.H.; Kondo, D.G.; Renshaw, P. An open-label pilot study of combined augmentation with creatine monohydrate and 5-hydroxytryptophan for selective serotonin reuptake inhibitor- or serotonin-norepinephrine reuptake inhibitor-resistant depression in adult women. J. Clin. Psychopharmacol. 2017, 37, 578–583. [Google Scholar] [CrossRef]
- Cunha, M.P.; Pazini, F.L.; Rosa, J.M.; Ramos-Hryb, A.B.; Oliveira, A.; Kaster, M.P.; Rodrigues, A.L. Creatine, similarly to ketamine, affords anti-depressant-like effects in the tail suspension test via adenosine A1 and A2A receptor activities. Purinergic Signal. 2015, 11, 215–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazini, F.L.; Cunha, M.P.; Rosa, J.M.; Colla, A.R.; Lieberknecht, V.; Oliveira, Á.; Rodrigues, A.L. Creatine, similar to keta-mine, counteracts depressive-like behavior induced by corticosterone via PI3K/Akt/mTOR pathway. Mol. Neurobiol. 2016, 53, 6818–6834. [Google Scholar] [CrossRef]
- Zhang, C.K.; Hewett, J.; Hemming, J.; Grant, T.; Zhao, H.; Abraham, C.; Oikonomou, I.; Berkowitz, M.; Cho, J.H.; Proctor, D.D. The Influence of Depression on Quality of Life in Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2013, 19, 1732–1739. [Google Scholar] [CrossRef] [Green Version]
- Glover, L.E.; Colgan, S.P. Epithelial barrier regulation by hypoxia inducible factor. Ann. Am. Thorac Soc. 2017, 14, S233–S236. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Wang, R.X.; Alexeev, E.E.; Colgan, S.P. Intestinal Inflammation as a Dysbiosis of Energy Procurement: New Insights into an Old Topic. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Colgan, S.P.; Curtis, V.F.; Lanis, J.M.; Glover, L.E. Metabolic regulation of intestinal epithelial barrier during inflammation. Tissue Barriers 2015, 3, e970936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, K.; Kokotilo, M.S.; Carter, J.; Thiesen, A.; Madsen, K.L.; Studzinski, J.; Khadaroo, R.G.; Churchill, T.A. Creatine-loading preserves intestinal barrier function during organ preservation. Cryobiology 2018, 84, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Turer, E.; McAlpine, W.; Wang, K.W.; Lu, T.; Li, X.; Tang, M.; Zhan, X.; Wang, T.; Zhan, X.; Bu, C.H.; et al. Creatine maintains intestinal homeostasis and protects against colitis. Proc. Natl. Acad. Sci. USA 2017, 114, E1273–E1281. [Google Scholar] [CrossRef] [Green Version]
- Ross, F.A.; MacKintosh, C.; Hardie, D.G. AMP-activated protein kinase: A cellular energy sensor that comes in 12 flavours. FEBS J. 2016, 283, 2987–3001. [Google Scholar] [CrossRef] [PubMed]
- Suter, M.; Riek, U.; Tuerk, R.; Schlattner, U.; Wallimann, T.; Neumann, D. Dissecting the role of 5′-AMPK for allosteric stimulation, activation and deactivation of AMP-activated protein kinase. J. Biol. Chem. 2006, 281, 32207–32216. [Google Scholar] [CrossRef] [Green Version]
- Woods, A.; Johnstone, S.R.; Dickerson, K.; Leiper, F.C.; Fryer, L.G.; Neumann, D.; Schlattner, U.; Wallimann, T.; Carlson, M.; Carling, D. LKB1 is the upstream kinase in the AMP-activated protein kinase cascade. Curr. Biol. 2003, 13, 2004–2008. [Google Scholar] [CrossRef] [Green Version]
- Alessi, D.R.; Sakamoto, K.; Bayascas, J.R. LKB1-dependent signaling pathways. Annu. Rev. Biochem. 2006, 75, 137–163. [Google Scholar] [CrossRef]
- Zheng, B.; Cantley, L.C. Regulation of epithelial tight junction assembly and disassembly by AMP-activated protein kinase. Proc. Natl. Acad. Sci. USA 2007, 104, 819–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, J.; You, Q.; He, S.; Meng, Q.; Gao, J.; Wu, X.; Shen, Y.; Sun, Y.; Wu, X.; et al. Activating AMPK to Restore Tight Junction Assembly in Intestinal Epithelium and to Attenuate Experimental Colitis by Metformin. Front. Pharmacol. 2018, 9, 761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.H. Metformin use is associated with a lower risk of inflammatory bowel disease in patients with type 2 diabetes mellitus. J. Crohns Colitis 2021, 15, 64–73. [Google Scholar] [CrossRef]
- Roy, A.; Lee, D. Dietary Creatine as a possible novel treatment for Crohn’s ileitis. ACG Case Rep. J. 2016, 3, e173. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, Z.; Yan, H.; Wang, W.; Wu, Z.; Zhang, F.; Zhang, Q.; Shi, G.; Du, J.; Cai, H.; et al. Creatine promotes cancer metastasis through activation of Smad2/3. Cell Metab. 2021, S1550-413100116-9. [Google Scholar] [CrossRef]
- Cella, P.S.; Marinello, P.C.; Padilha, C.S.; Testa, M.T.; Guirro, P.B.; Cecchini, R.; Duarte, J.A.; Guarnier, F.A.; Deminice, R. Creatine supplementation does not promote tumor growth or enhance tumor aggressiveness in Walker-256 tumor-bearing rats. Nutrition 2020, 79, 110958. [Google Scholar] [CrossRef]
- Campos-Ferraz, P.L.; Gualano, B.; das Neves, W.; Andrade, I.T.; Hangai, I.; Pereira, R.T.; Bezerra, R.N.; Deminice, R.; Seelaender, M.; Lancha, A.H. Exploratory studies of the potential anti-cancer effects of creatine. Amino Acids 2016, 48, 1993–2001. [Google Scholar] [CrossRef]
- Li, B.; Yang, L. Creatine in T cell antitumor immunity and cancer therapy. Nutrients 2021, (in press). [Google Scholar]
- Fairman, C.M.; Kendall, K.L.; Hart, N.H.; Taaffe, D.R.; Galvão, D.A.; Newton, R.U. The potential therapeutic effects of creatine supplementation on body composition and muscle function in cancer. Crit. Rev. Oncol. Hematol. 2019, 133, 46–57. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallimann, T.; Hall, C.H.T.; Colgan, S.P.; Glover, L.E. Creatine Supplementation for Patients with Inflammatory Bowel Diseases: A Scientific Rationale for a Clinical Trial. Nutrients 2021, 13, 1429. https://doi.org/10.3390/nu13051429
Wallimann T, Hall CHT, Colgan SP, Glover LE. Creatine Supplementation for Patients with Inflammatory Bowel Diseases: A Scientific Rationale for a Clinical Trial. Nutrients. 2021; 13(5):1429. https://doi.org/10.3390/nu13051429
Chicago/Turabian StyleWallimann, Theo, Caroline H. T. Hall, Sean P. Colgan, and Louise E. Glover. 2021. "Creatine Supplementation for Patients with Inflammatory Bowel Diseases: A Scientific Rationale for a Clinical Trial" Nutrients 13, no. 5: 1429. https://doi.org/10.3390/nu13051429