
Three-Generation Study of Male Rats Gestationally Exposed to High Butterfat and Bisphenol A: Impaired Spermatogenesis, Penetrance with Reduced Severity

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Hormone Treatment SD Rat Model
2.3. Natural Aging Model
2.4. Tissue Collection and Immunohistochemistry
2.5. Quantitative Real-Time PCR (qPCR)
2.6. Statistical Analysis
3. Results
3.1. Prenatal HFB + BPA Exposure Induces Significant Spermatogenesis Arrest in T + E2 Implanted Offspring
3.2. CYP19A1/Aromatase Expression during Spermatogenesis in T + E2-Implanted Offspring
3.3. Decrease in Cytoplasmic ERbeta Expression in Round Spermatids of T + E2-Implanted Offspring
3.4. Nuclear ERbeta Localization in Round Spermatids in T + E2-Implanted Offspring
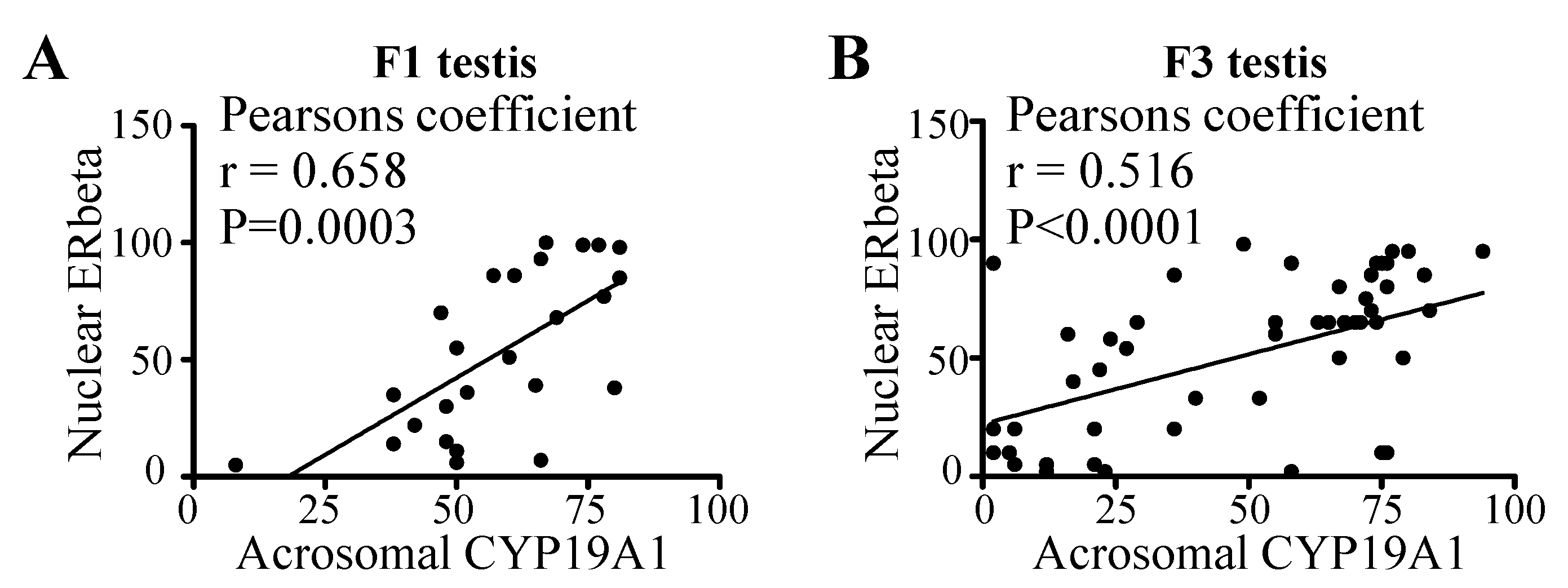
3.5. Correlation between Acrosomal CYP19A1 and Nuclear ERbeta Localization in Round Spermatids
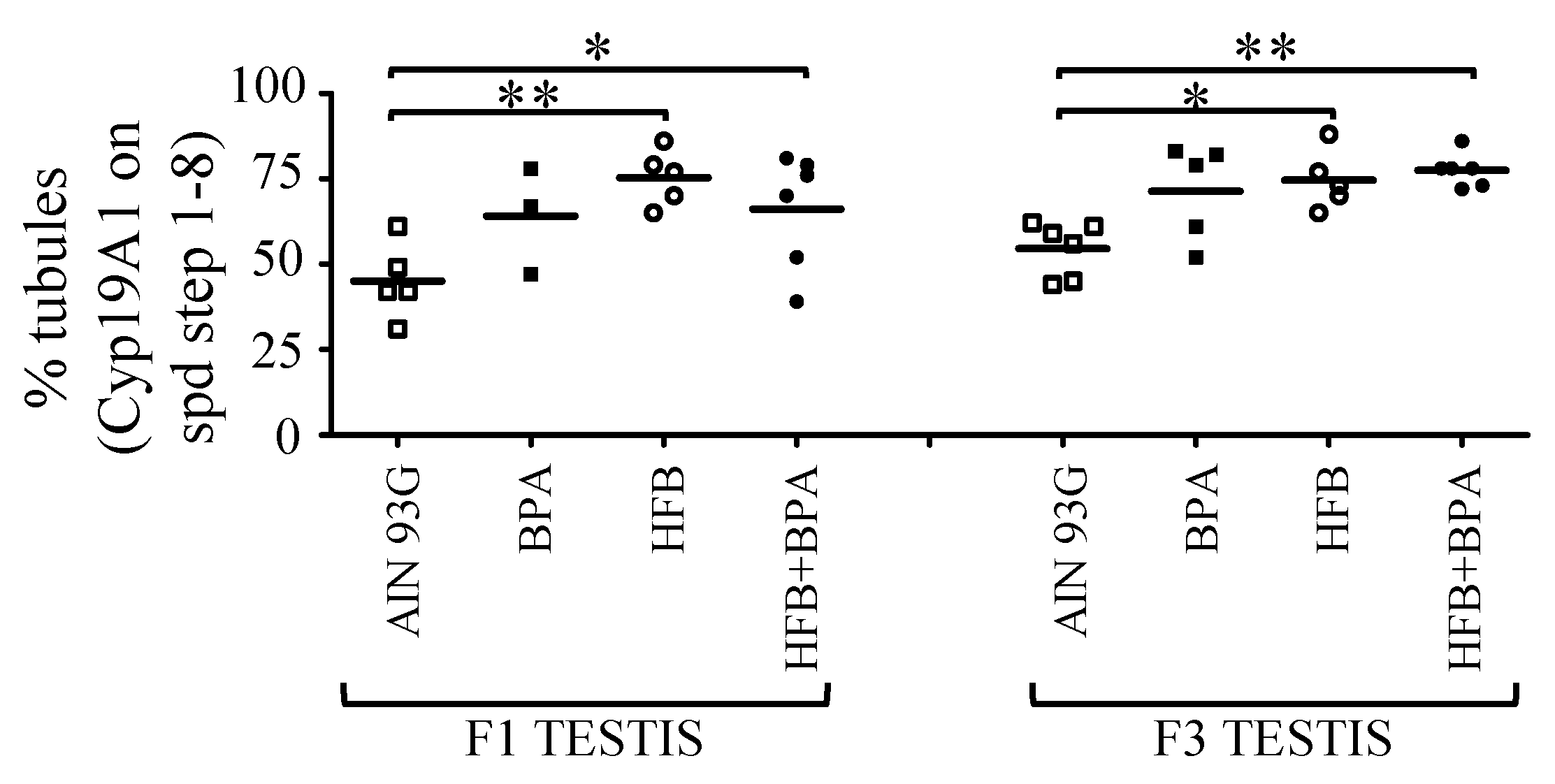
3.6. 18-Month-Old F3-Generation Testes Exhibited Decreased Spermatozoa Numbers
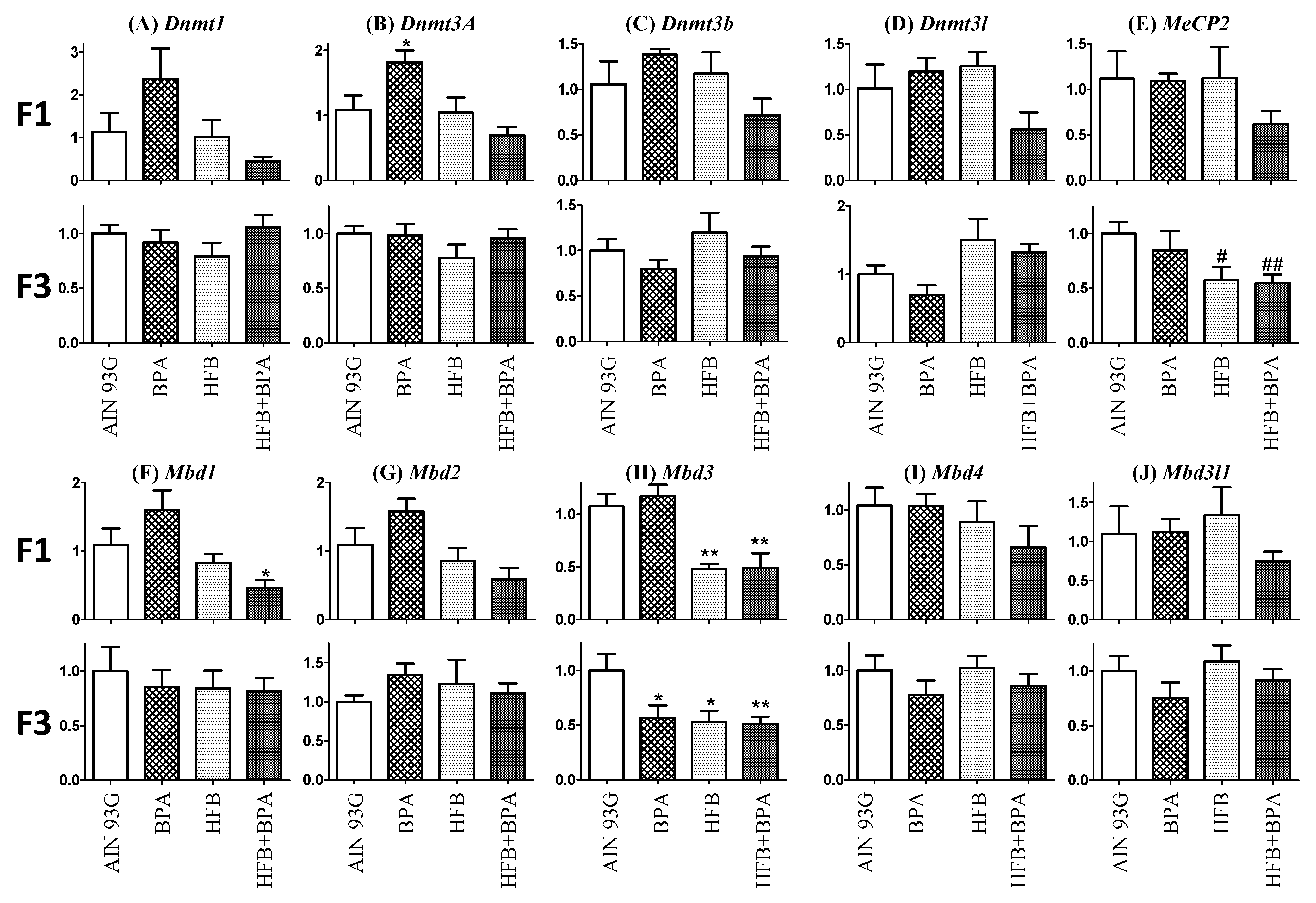
3.7. Methyl-CpG-Binding Domain (MBD3) Levels Are Reduced in HFB and HFB + BPA Group Offspring (T + E2-Treated)
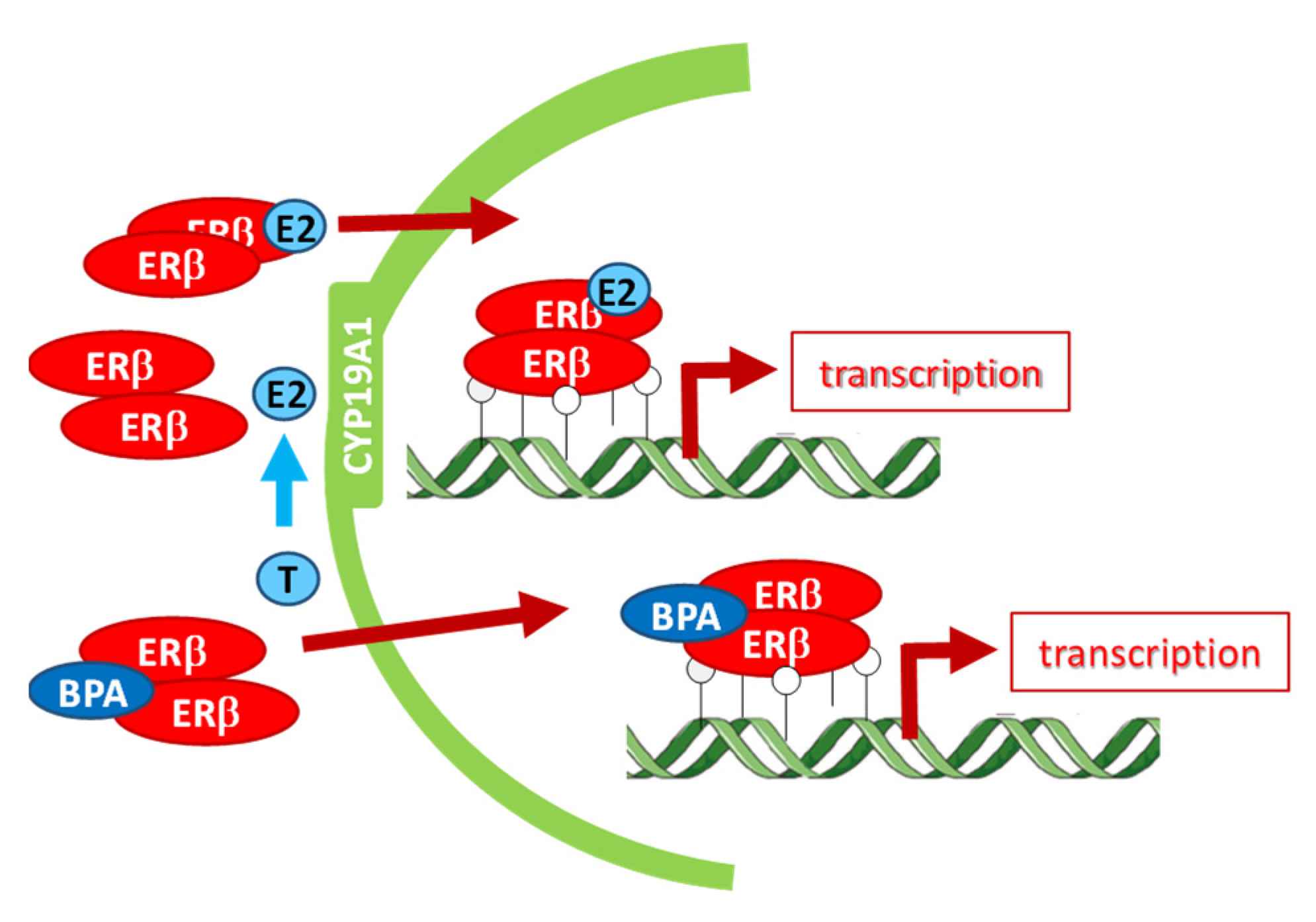
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanson, M.A.; Gluckman, P.D. Early developmental conditioning of later health and disease: Physiology or pathophysiology? Physiol. Rev. 2014, 94, 1027–1076. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A. Maternal constraint of fetal growth and its consequences. Semin. Fetal. Neonatal Med. 2004, 9, 419–425. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in England and Wales. Lancet 1986, 1, 1077–1081. [Google Scholar] [CrossRef]
- Roseboom, T.J.; van der Meulen, J.H.; Ravelli, A.C.; Osmond, C.; Barker, D.J.; Bleker, O.P. Effects of prenatal exposure to the Dutch famine on adult disease in later life: An overview. Mol. Cell Endocrinol. 2001, 185, 93–98. [Google Scholar] [CrossRef]
- Sun, B.; Purcell, R.H.; Terrillion, C.E.; Yan, J.; Moran, T.H.; Tamashiro, K.L. Maternal high-fat diet during gestation or suckling differentially affects offspring leptin sensitivity and obesity. Diabetes 2012, 61, 2833–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mente, A.; de Koning, L.; Shannon, H.S.; Anand, S.S. A systematic review of the evidence supporting a causal link between dietary factors and coronary heart disease. Arch. Intern. Med. 2009, 169, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Madeo, F.; Carmona-Gutierrez, D.; Hofer, S.J.; Kroemer, G. Caloric Restriction Mimetics against Age-Associated Disease: Targets, Mechanisms, and Therapeutic Potential. Cell Metab. 2019, 29, 592–610. [Google Scholar] [CrossRef] [Green Version]
- Maffini, M.V.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Endocrine disruptors and reproductive health: The case of bisphenol-A. Mol. Cell Endocrinol. 2006, 254-255, 179–186. [Google Scholar] [CrossRef]
- Murono, E.P.; Derk, R.C. The reported active metabolite of methoxychlor, 2,2-bis(p-hydroxyphenyl)-1,1,1-trichloroethane, inhibits testosterone formation by cultured Leydig cells from neonatal rats. Reprod. Toxicol. 2005, 20, 503–513. [Google Scholar] [CrossRef]
- Gray, L.E., Jr.; Ostby, J.; Monosson, E.; Kelce, W.R. Environmental antiandrogens: Low doses of the fungicide vinclozolin alter sexual differentiation of the male rat. Toxicol. Ind. Health 1999, 15, 48–64. [Google Scholar] [CrossRef]
- Ho, S.M.; Cheong, A.; Adgent, M.A.; Veevers, J.; Suen, A.A.; Tam, N.N.C.; Leung, Y.K.; Jefferson, W.N.; Williams, C.J. Environmental factors, epigenetics, and developmental origin of reproductive disorders. Reprod. Toxicol. 2017, 68, 85–104. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.M. Environmental epigenetics of asthma: An update. J. Allergy Clin. Immunol. 2010, 126, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.M.; Johnson, A.; Tarapore, P.; Janakiram, V.; Zhang, X.; Leung, Y.K. Environmental epigenetics and its implication on disease risk and health outcomes. ILAR J. 2012, 53, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ho, S.M. Epigenetics meets endocrinology. J. Mol. Endocrinol. 2011, 46, R11–R32. [Google Scholar] [CrossRef] [Green Version]
- Tarapore, P.; Hennessy, M.; Song, D.; Ying, J.; Ouyang, B.; Govindarajah, V.; Leung, Y.K.; Ho, S.M. High butter-fat diet and bisphenol A additively impair male rat spermatogenesis. Reprod. Toxicol. 2017, 68, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damstra, T.; Barlow, S.; Bergman, A.; Kavlock, R.; Van Der Kraak, G. Global Assessment of the State-of-the-Science of Endocrine Disruptors; International Programme on Chemical Safety; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Rochester, J.R. Bisphenol A and human health: A review of the literature. Reprod. Toxicol. 2013, 42, 132–155. [Google Scholar] [CrossRef] [PubMed]
- Tomza-Marciniak, A.; Stepkowska, P.; Kuba, J.; Pilarczyk, B. Effect of bisphenol A on reproductive processes: A review of in vitro, in vivo and epidemiological studies. J. Appl. Toxicol. 2018, 38, 51–80. [Google Scholar] [CrossRef]
- Peretz, J.; Vrooman, L.; Ricke, W.A.; Hunt, P.A.; Ehrlich, S.; Hauser, R.; Padmanabhan, V.; Taylor, H.S.; Swan, S.H.; VandeVoort, C.A.; et al. Bisphenol a and reproductive health: Update of experimental and human evidence, 2007–2013. Environ. Health Perspect. 2014, 122, 775–786. [Google Scholar] [CrossRef]
- Sharpe, R.M. Environmental/lifestyle effects on spermatogenesis. Philos. Trans. R. Soc. Lond B Biol. Sci. 2010, 365, 1697–1712. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Calafat, A.M.; Kuklenyik, Z.; Reidy, J.A.; Caudill, S.P.; Ekong, J.; Needham, L.L. Urinary concentrations of bisphenol A and 4-nonylphenol in a human reference population. Environ. Health Perspect. 2005, 113, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Choi, K.; Park, J.; Moon, H.B.; Choi, G.; Lee, J.J.; Suh, E.; Kim, H.J.; Eun, S.H.; Kim, G.H.; et al. Bisphenol A distribution in serum, urine, placenta, breast milk, and umbilical cord serum in a birth panel of mother-neonate pairs. Sci. Total Environ. 2018, 626, 1494–1501. [Google Scholar] [CrossRef]
- Mendonca, K.; Hauser, R.; Calafat, A.M.; Arbuckle, T.E.; Duty, S.M. Bisphenol A concentrations in maternal breast milk and infant urine. Int. Arch. Occup. Environ. Health 2014, 87, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Boudalia, S.; Berges, R.; Chabanet, C.; Folia, M.; Decocq, L.; Pasquis, B.; Abdennebi-Najar, L.; Canivenc-Lavier, M.C. A multi-generational study on low-dose BPA exposure in Wistar rats: Effects on maternal behavior, flavor intake and development. Neurotoxicol. Teratol. 2014, 41, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.K.; Govindarajah, V.; Cheong, A.; Veevers, J.; Song, D.; Gear, R.; Zhu, X.; Ying, J.; Kendler, A.; Medvedovic, M.; et al. Gestational high-fat diet and bisphenol A exposure heightens mammary cancer risk. Endocr. Relat Cancer 2017, 24, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medwid, S.; Guan, H.; Yang, K. Prenatal exposure to bisphenol A disrupts adrenal steroidogenesis in adult mouse offspring. Environ. Toxicol. Pharmacol. 2016, 43, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Bu, P.; Li, F.; Lan, S.; Wu, H.; Yuan, L.; Wang, Y. Neonatal bisphenol A exposure induces meiotic arrest and apoptosis of spermatogenic cells. Oncotarget 2016, 7, 10606–10615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohue, K.M.; Miller, R.L.; Perzanowski, M.S.; Just, A.C.; Hoepner, L.A.; Arunajadai, S.; Canfield, S.; Resnick, D.; Calafat, A.M.; Perera, F.P.; et al. Prenatal and postnatal bisphenol A exposure and asthma development among inner-city children. J. Allergy Clin. Immunol. 2013, 131, 736–742. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, J.T.; Edwards, M.; Shetty, S.R.; Gatewood, J.D.; Taylor, J.A.; Rissman, E.F.; Connelly, J.J. Gestational exposure to bisphenol a produces transgenerational changes in behaviors and gene expression. Endocrinology 2012, 153, 3828–3838. [Google Scholar] [CrossRef] [Green Version]
- Salian, S.; Doshi, T.; Vanage, G. Perinatal exposure of rats to Bisphenol A affects fertility of male offspring—An overview. Reprod. Toxicol. 2011, 31, 359–362. [Google Scholar] [CrossRef]
- Munoz-de-Toro, M.; Markey, C.M.; Wadia, P.R.; Luque, E.H.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Perinatal exposure to bisphenol-A alters peripubertal mammary gland development in mice. Endocrinology 2005, 146, 4138–4147. [Google Scholar] [CrossRef]
- Tarapore, P.; Hennessy, M.; Song, D.; Ying, J.; Ouyang, B.; Govindarajah, V.; Leung, Y.K.; Ho, S.M. Data on spermatogenesis in rat males gestationally exposed to bisphenol A and high fat diets. Data Brief. 2016, 9, 812–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirihagalle, S.; You, T.; Suh, L.; Patel, C.; Gao, L.; Rattan, S.; Qiao, H. Prenatal exposure to di-(2-ethylhexyl) phthalate and high-fat diet synergistically disrupts mouse fetal oogenesis and affects folliculogenesis. Biol. Reprod. 2019, 100, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Oshio, L.T.; Andreazzi, A.E.; Lopes, J.F.; Sá, J.P.; Bolotari, M.; Costa, V.M.G.; Guerra, M.O.; Peters, V.M. A paternal hypercaloric diet affects the metabolism and fertility of F1 and F2 Wistar rat generations. J. Dev. Orig. Health Dis. 2020, 11, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Ramaiyan, B.; Zarei, M.; Acharya, P.; Talahalli, R.R. Dietary n-3 but not n-6 fatty acids modulate anthropometry and fertility indices in high-fat diet fed rats: A two-generation study. J. Food Sci. Technol. 2021, 58, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Bertoldo, M.J.; Uddin, G.M.; Youngson, N.A.; Agapiou, D.; Walters, K.A.; Sinclair, D.A.; Morris, M.J.; Gilchrist, R.B. Multigenerational obesity-induced perturbations in oocyte-secreted factor signalling can be ameliorated by exercise and nicotinamide mononucleotide. Hum. Reprod. Open. 2018, 2018, hoy010. [Google Scholar] [CrossRef] [Green Version]
- Finger, B.J.; Harvey, A.J.; Green, M.P.; Gardner, D.K. Combined parental obesity negatively impacts preimplantation mouse embryo development, kinetics, morphology and metabolism. Hum. Reprod. 2015, 30, 2084–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organisation. Obesity and Overweight. Fact Sheet. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 20 September 2021).
- Chavatte-Palmer, P.; Tarrade, A.; Levy, R. [Developmental origins of health and disease in adults: Role of maternal environment]. Gynecol. Obstet. Fertil. 2012, 40, 517–519. [Google Scholar] [CrossRef]
- Lane, M.; Robker, R.L.; Robertson, S.A. Parenting from before conception. Science 2014, 345, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Blatt, J.; Van, L.L.; Weiner, T.; Sailer, S. Ovarian carcinoma in an adolescent with transgenerational exposure to diethylstilbestrol. J. Pediatr. Hematol. Oncol. 2003, 25, 635–636. [Google Scholar] [CrossRef]
- Drake, A.J.; Walker, B.R.; Seckl, J.R. Intergenerational consequences of fetal programming by in utero exposure to glucocorticoids in rats. Am. J. Physiol Regul. Integr. Comp Physiol 2005, 288, R34–R38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, S.; Li, S.; Feng, R.; Na, L.; Chu, X.; Wu, X.; Niu, Y.; Sun, Z.; Han, T.; et al. Prenatal exposure to famine and the development of hyperglycemia and type 2 diabetes in adulthood across consecutive generations: A population-based cohort study of families in Suihua, China. Am. J. Clin. Nutr. 2017, 105, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumey, L.H.; Stein, A.D. In utero exposure to famine and subsequent fertility: The Dutch Famine Birth Cohort Study. Am. J. Public Health 1997, 87, 1962–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrzynska, M.M.; Tyrkiel, E.J. The effect of preconceptional exposure of F0 male mice to di(2-ethylhexyl)phthalate on the induction of reproductive toxicity in F2 generation. Drug Chem. Toxicol. 2018, 1–6. [Google Scholar] [CrossRef]
- Chen, J.; Wu, S.; Wen, S.; Shen, L.; Peng, J.; Yan, C.; Cao, X.; Zhou, Y.; Long, C.; Lin, T.; et al. The Mechanism of Environmental Endocrine Disruptors (DEHP) Induces Epigenetic Transgenerational Inheritance of Cryptorchidism. PLoS ONE 2015, 10, e0126403. [Google Scholar] [CrossRef]
- Guerrero-Bosagna, C.; Covert, T.R.; Haque, M.M.; Settles, M.; Nilsson, E.E.; Anway, M.D.; Skinner, M.K. Epigenetic transgenerational inheritance of vinclozolin induced mouse adult onset disease and associated sperm epigenome biomarkers. Reprod. Toxicol. 2012, 34, 694–707. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Bosagna, C.M.; Skinner, M.K. Epigenetic transgenerational effects of endocrine disruptors on male reproduction. Semin. Reprod. Med. 2009, 27, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, E.; Martinez-Samayoa, P.M.; Bautista, C.J.; Deas, M.; Guillen, L.; Rodriguez-Gonzalez, G.L.; Guzman, C.; Larrea, F.; Nathanielsz, P.W. Sex differences in transgenerational alterations of growth and metabolism in progeny (F2) of female offspring (F1) of rats fed a low protein diet during pregnancy and lactation. J. Physiol 2005, 566, 225–236. [Google Scholar] [CrossRef]
- Skinner, M.K.; Anway, M.D. Seminiferous cord formation and germ-cell programming: Epigenetic transgenerational actions of endocrine disruptors. Ann. N. Y. Acad. Sci. 2005, 1061, 18–32. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, Y.J.; Tain, Y.L. Maternal Exposure to Bisphenol A Combined with High-Fat Diet-Induced Programmed Hypertension in Adult Male Rat Offspring: Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 4382. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Fan, Y.; Zhao, N.; Yang, H.; Ye, X.; He, D.; Jin, X.; Liu, J.; Tian, C.; Li, H.; et al. High-fat diet aggravates glucose homeostasis disorder caused by chronic exposure to bisphenol A. J. Endocrinol. 2014, 221, 167–179. [Google Scholar] [CrossRef]
- Andreollo, N.A.; Santos, E.F.; Araujo, M.R.; Lopes, L.R. Rat’s age versus human’s age: What is the relationship? Arq Bras. Cir. Dig. 2012, 25, 49–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, S.M.; Tang, W.Y.; Belmonte de, F.J.; Prins, G.S. Developmental exposure to estradiol and bisphenol A increases susceptibility to prostate carcinogenesis and epigenetically regulates phosphodiesterase type 4 variant 4. Cancer Res. 2006, 66, 5624–5632. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Prins, G.S.; Henneberry, M.O.; Grayhack, J.T. Effect of Estradiol on the Rat Prostate in the Presence and Absence of Testosterone and Pituitary. J Androl. 1981, 2, 293–299. [Google Scholar] [CrossRef]
- Clarke, M.; Pearl, C.A. Alterations in the estrogen environment of the testis contribute to declining sperm production in aging rats. Syst. Biol. Reprod. Med. 2014, 60, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.Y.; Morey, L.M.; Cheung, Y.Y.; Birch, L.; Prins, G.S.; Ho, S.M. Neonatal exposure to estradiol/bisphenol A alters promoter methylation and expression of Nsbp1 and Hpcal1 genes and transcriptional programs of Dnmt3a/b and Mbd2/4 in the rat prostate gland throughout life. Endocrinology 2012, 153, 42–55. [Google Scholar] [CrossRef]
- Thompson, C.J.; Tam, N.N.; Joyce, J.M.; Leav, I.; Ho, S.M. Gene expression profiling of testosterone and estradiol-17 beta-induced prostatic dysplasia in Noble rats and response to the antiestrogen ICI 182,780. Endocrinology 2002, 143, 2093–2105. [Google Scholar] [CrossRef]
- Lombo, M.; Fernandez-Diez, C.; Gonzalez-Rojo, S.; Herraez, M.P. Genetic and epigenetic alterations induced by bisphenol A exposure during different periods of spermatogenesis: From spermatozoa to the progeny. Sci. Rep. 2019, 9, 18029. [Google Scholar] [CrossRef] [Green Version]
- Dumasia, K.; Kumar, A.; Deshpande, S.; Sonawane, S.; Balasinor, N.H. Differential roles of estrogen receptors, ESR1 and ESR2, in adult rat spermatogenesis. Mol. Cell Endocrinol. 2016, 428, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Delbes, G.; Levacher, C.; Pairault, C.; Racine, C.; Duquenne, C.; Krust, A.; Habert, R. Estrogen receptor beta-mediated inhibition of male germ cell line development in mice by endogenous estrogens during perinatal life. Endocrinology 2004, 145, 3395–3403. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, Z.; Wang, G.; Li, Z.; Liang, A.; Wang, H.; Dai, Y.; Huang, X.; Chen, X.; Ma, Y.; et al. Overexpression of Human-Derived DNMT3A Induced Intergenerational Inheritance of Active DNA Methylation Changes in Rat Sperm. Front Genet. 2017, 8, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salian, S.; Doshi, T.; Vanage, G. Perinatal exposure of rats to Bisphenol A affects the fertility of male offspring. Life Sci. 2009, 85, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Ziv-Gal, A.; Wang, W.; Zhou, C.; Flaws, J.A. The effects of in utero bisphenol A exposure on reproductive capacity in several generations of mice. Toxicol. Appl. Pharmacol. 2015, 284, 354–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, J.T.; Goldsby, J.A.; Rissman, E.F. Transgenerational effects of prenatal bisphenol A on social recognition. Horm. Behav. 2013, 64, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS ONE 2013, 8, e55387. [Google Scholar] [CrossRef] [PubMed]
- Eddy, E.M.; Washburn, T.F.; Bunch, D.O.; Goulding, E.H.; Gladen, B.C.; Lubahn, D.B.; Korach, K.S. Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinology 1996, 137, 4796–4805. [Google Scholar] [CrossRef]
- Fisher, C.R.; Graves, K.H.; Parlow, A.F.; Simpson, E.R. Characterization of mice deficient in aromatase (ArKO) because of targeted disruption of the cyp19 gene. Proc. Natl. Acad. Sci. USA 1998, 95, 6965–6970. [Google Scholar] [CrossRef] [Green Version]
- Robertson, K.M.; O’Donnell, L.; Jones, M.E.; Meachem, S.J.; Boon, W.C.; Fisher, C.R.; Graves, K.H.; McLachlan, R.I.; Simpson, E.R. Impairment of spermatogenesis in mice lacking a functional aromatase (cyp 19) gene. Proc. Natl. Acad. Sci. USA 1999, 96, 7986–7991. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, H.; Jia, L.; Li, X.; Rahman, N. Oestrogen action and male fertility: Experimental and clinical findings. Cell Mol. Life Sci. 2015, 72, 3915–3930. [Google Scholar] [CrossRef]
- Li, X.; Strauss, L.; Kaatrasalo, A.; Mayerhofer, A.; Huhtaniemi, I.; Santti, R.; Makela, S.; Poutanen, M. Transgenic mice expressing p450 aromatase as a model for male infertility associated with chronic inflammation in the testis. Endocrinology 2006, 147, 1271–1277. [Google Scholar] [CrossRef] [Green Version]
- Carreau, S.; Hess, R.A. Oestrogens and spermatogenesis. Philos. Trans. R. Soc. Lond B Biol. Sci. 2010, 365, 1517–1535. [Google Scholar] [CrossRef] [Green Version]
- Selva, D.M.; Tirado, O.M.; Toran, N.; Suarez-Quian, C.A.; Reventos, J.; Munell, F. Estrogen receptor beta expression and apoptosis of spermatocytes of mice overexpressing a rat androgen-binding protein transgene. Biol. Reprod. 2004, 71, 1461–1468. [Google Scholar] [CrossRef] [Green Version]
- Chimento, A.; Sirianni, R.; Zolea, F.; Bois, C.; Delalande, C.; Ando, S.; Maggiolini, M.; Aquila, S.; Carreau, S.; Pezzi, V. Gper and ESRs are expressed in rat round spermatids and mediate oestrogen-dependent rapid pathways modulating expression of cyclin B1 and Bax. Int. J. Androl. 2011, 34, 420–429. [Google Scholar] [CrossRef]
- Fowler, K.A.; Gill, K.; Kirma, N.; Dillehay, D.L.; Tekmal, R.R. Overexpression of aromatase leads to development of testicular leydig cell tumors: An in vivo model for hormone-mediated TesticularCancer. Am. J. Pathol. 2000, 156, 347–353. [Google Scholar] [CrossRef]
- Laing, L.V.; Viana, J.; Dempster, E.L.; Uren Webster, T.M.; van Aerle, R.; Mill, J.; Santos, E.M. Sex-specific transcription and DNA methylation profiles of reproductive and epigenetic associated genes in the gonads and livers of breeding zebrafish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 222, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Rojo, S.; Lombo, M.; Fernandez-Diez, C.; Herraez, M.P. Male exposure to bisphenol a impairs spermatogenesis and triggers histone hyperacetylation in zebrafish testes. Environ. Pollut. 2019, 248, 368–379. [Google Scholar] [CrossRef]
- Santangeli, S.; Maradonna, F.; Gioacchini, G.; Cobellis, G.; Piccinetti, C.C.; Dalla, V.L.; Carnevali, O. BPA-Induced Deregulation of Epigenetic Patterns: Effects on Female Zebrafish Reproduction. Sci. Rep. 2016, 6, srep21982. [Google Scholar] [CrossRef]
- Chen, J.; Xiao, Y.; Gai, Z.; Li, R.; Zhu, Z.; Bai, C.; Tanguay, R.L.; Xu, X.; Huang, C.; Dong, Q. Reproductive toxicity of low level bisphenol A exposures in a two-generation zebrafish assay: Evidence of male-specific effects. Aquat. Toxicol. 2015, 169, 204–214. [Google Scholar] [CrossRef]
- Dolinoy, D.C.; Huang, D.; Jirtle, R.L. Maternal nutrient supplementation counteracts bisphenol A-induced DNA hypomethylation in early development. Proc. Natl. Acad. Sci. USA 2007, 104, 13056–13061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, G.; Liand, M.; Mauduit, C.; Chehade, H.; Benahmed, M.; Simeoni, U.; Siddeek, B. Maternal Exposure to High-Fat Diet Induces Long-Term Derepressive Chromatin Marks in the Heart. Nutrients 2020, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.C.; Chen, Y.Z.; Wang, C.H.; Lin, F.J. The nonalcoholic fatty liver disease-like phenotype and lowered serum VLDL are associated with decreased expression and DNA hypermethylation of hepatic ApoB in male offspring of ApoE deficient mothers fed a with Western diet. J. Nutr. Biochem. 2020, 77, 108319. [Google Scholar] [CrossRef] [PubMed]
- Keleher, M.R.; Zaidi, R.; Shah, S.; Oakley, M.E.; Pavlatos, C.; El, I.S.; Xing, X.; Li, D.; Wang, T.; Cheverud, J.M. Maternal high-fat diet associated with altered gene expression, DNA methylation, and obesity risk in mouse offspring. PLoS ONE 2018, 13, e0192606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, E.J.; Lim, D.S.; Lee, J.S. A novel role for methyl CpG-binding domain protein 3, a component of the histone deacetylase complex, in regulation of cell cycle progression and cell death. Biochem. Biophys. Res. Commun. 2009, 378, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Ozmadenci, D.; Feraud, O.; Markossian, S.; Kress, E.; Ducarouge, B.; Gibert, B.; Ge, J.; Durand, I.; Gadot, N.; Plateroti, M.; et al. Netrin-1 regulates somatic cell reprogramming and pluripotency maintenance. Nat. Commun. 2015, 6, 7398. [Google Scholar] [CrossRef] [Green Version]
- Das, P.M.; Ramachandran, K.; Vanwert, J.; Ferdinand, L.; Gopisetty, G.; Reis, I.M.; Singal, R. Methylation mediated silencing of TMS1/ASC gene in prostate cancer. Mol. Cancer 2006, 5, 28. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide/Protein Target | Antigen Sequence | Name of Antibody | Manufacturer, Catalog Number | Species, Monoclonal or Polyclonal | Dilution |
|---|---|---|---|---|---|
| CYP19A1 | 209-503 | H-300 | Santa Cruz Biotechnology, sc-30086 | Rabbit polyclonal | 1:50 |
| ESR2/ERbeta | 17-mer, close to C-terminus | anti-ESR2 | BioGeneX, AR385 | Rabbit polyclonal | 1:100 |
| Primer Name | Primer Sequence | Amplicon Size (bps) | Annealing Temperature | |
|---|---|---|---|---|
| rDNMT1 | Forward: | 5′-GAGGTGGGCGACTGCGTCTC-3′ | 214 | 60 |
| Reverse: | 5′-TGTGGATGTAGGAAAGTTGCA-3′ | |||
| rDNMT3a | Forward: | 5′-CAGAATAGCCAAGTTCAGCAAAGTGA-3′ | 68 | 58 |
| Reverse: | 5′-CTTTGCCCTGCTTTATGGAG-3′ | |||
| rDNMT3b | Forward: | 5′-GTTAAAGAAAGTACAGACAATAACCAC-3′ | 220 | 57 |
| Reverse: | 5′-TCTGATGACTGGCACACTCC-3′ | |||
| rDNMT3l | Forward: | 5’-AATGGCCGAAATCAGCCCCA-3’ | 139 | 60 |
| Reverse: | 5’-CGCTGGTTCACGTTGACTTC-3’ | |||
| rMeCP2 | Forward: | 5′-GTCGCTCTGCTGGAAAGTAT-3′ | 189 | 57 |
| Reverse: | 5′-TGGGCTTCTTAGGTGGTTTC-3′ | |||
| rMBD1 | Forward: | 5′-CAGCAGTCACAACCTTCCTG-3′ | 182 | 58 |
| Reverse: | 5′-GGTGCCAATCCCTCCTATCT-3′ | |||
| rMBD2 | Forward: | 5′-GTCGGCCCAGGTAGTAATGAT-3′ | 195 | 60 |
| Reverse: | 5′-GACTCGCTCTTCCTGTTTCCT-3′ | |||
| rMBD3 | Forward: | 5′-CTGAACACTGCACTGCCTGTA-3′ | 145 | 58 |
| Reverse: | 5′-GTTTCTTCTCCCAGAAAAGCTG-3′ | |||
| rMBD4 | Forward: | 5′-CCTACCGGATCTTTTGTGTCA-3′ | 90 | 58 |
| Reverse: | 5′-GATTTTCCCAAAGCCAGTCAT-3′ | |||
| rMBP3l1 | Forward: | 5’-GCTGGTTGGAGACTGGCAAT-3’ | 96 | 60 |
| Reverse: | 5’-TTGCCCATCTGACTCCGTTC-3’ | |||
| rRpl19 | Forward: | 5′-GCATATGGGCATAGGGAAGA-3′ | 197 | 58 |
| Reverse: | 5′-CCATGAGAATCCGCTTGTTT-3′ |
| Number of Animals | % Animals (Normal) | Odds Ratio | p-Value | |
|---|---|---|---|---|
| AIN | 10/11 | 91% | ||
| BPA | 10/14 | 71% | 4 | 0.340 |
| HFB | 9/14 | 64% | 5.6 | 0.180 |
| HFB + BPA | 6/15 | 40% | 15 | 0.014 * |
| Number of Animals | % Animals (Normal) | Odds Ratio | p-Value | |
|---|---|---|---|---|
| AIN | 6/7 | 85.7% | ||
| BPA25 | 4/5 | 80.0% | 1.5 | 1 |
| HFB | 4/5 | 80.0% | 1.5 | 1 |
| HFB + BPA | 3/6 | 50.0% | 6.0 | 0.265 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, S.-M.; Rao, R.; Ouyang, B.; Tam, N.N.C.; Schoch, E.; Song, D.; Ying, J.; Leung, Y.-K.; Govindarajah, V.; Tarapore, P. Three-Generation Study of Male Rats Gestationally Exposed to High Butterfat and Bisphenol A: Impaired Spermatogenesis, Penetrance with Reduced Severity. Nutrients 2021, 13, 3636. https://doi.org/10.3390/nu13103636
Ho S-M, Rao R, Ouyang B, Tam NNC, Schoch E, Song D, Ying J, Leung Y-K, Govindarajah V, Tarapore P. Three-Generation Study of Male Rats Gestationally Exposed to High Butterfat and Bisphenol A: Impaired Spermatogenesis, Penetrance with Reduced Severity. Nutrients. 2021; 13(10):3636. https://doi.org/10.3390/nu13103636
Chicago/Turabian StyleHo, Shuk-Mei, Rahul Rao, Bin Ouyang, Neville N. C. Tam, Emma Schoch, Dan Song, Jun Ying, Yuet-Kin Leung, Vinothini Govindarajah, and Pheruza Tarapore. 2021. "Three-Generation Study of Male Rats Gestationally Exposed to High Butterfat and Bisphenol A: Impaired Spermatogenesis, Penetrance with Reduced Severity" Nutrients 13, no. 10: 3636. https://doi.org/10.3390/nu13103636