Osteoprotective Activity and Metabolite Fingerprint via UPLC/MS and GC/MS of Lepidium sativum in Ovariectomized Rats

 ,
,  ,
,  , and
, and
Abstract
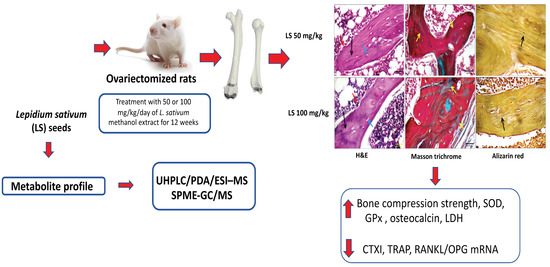
:
1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Extraction of Plant Material
2.3. Biological Study
2.3.1. Determination of Oral Lethal Dose 50 (LD50)
2.3.2. Animals and Animal Treatment
2.3.3. Determination of Relative Weights and Compression Strength
2.3.4. Histological Examination
2.3.5. Assessment of Serum Markers of Bone Metabolism
2.3.6. Preparation of Bone Homogenate
2.3.7. Bone Tissue Oxidative Status Assessment
2.3.8. RANKL and OPG Assessment via Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.3.9. Statistical Analysis
2.4. Metabolites Characterization of L. sativum Seed Extract
2.4.1. Extraction and Preparation of Samples for UHPLC/PDA/ESI-MS
2.4.2. Analysis of UHPLC-Orbitrap-HRMS
2.4.3. Volatiles Analysis of L. sativum Seeds via Headspace GC/MS
3. Results and Discussion
3.1. Biological Study
3.1.1. Oral LD50 of L. sativum Extract
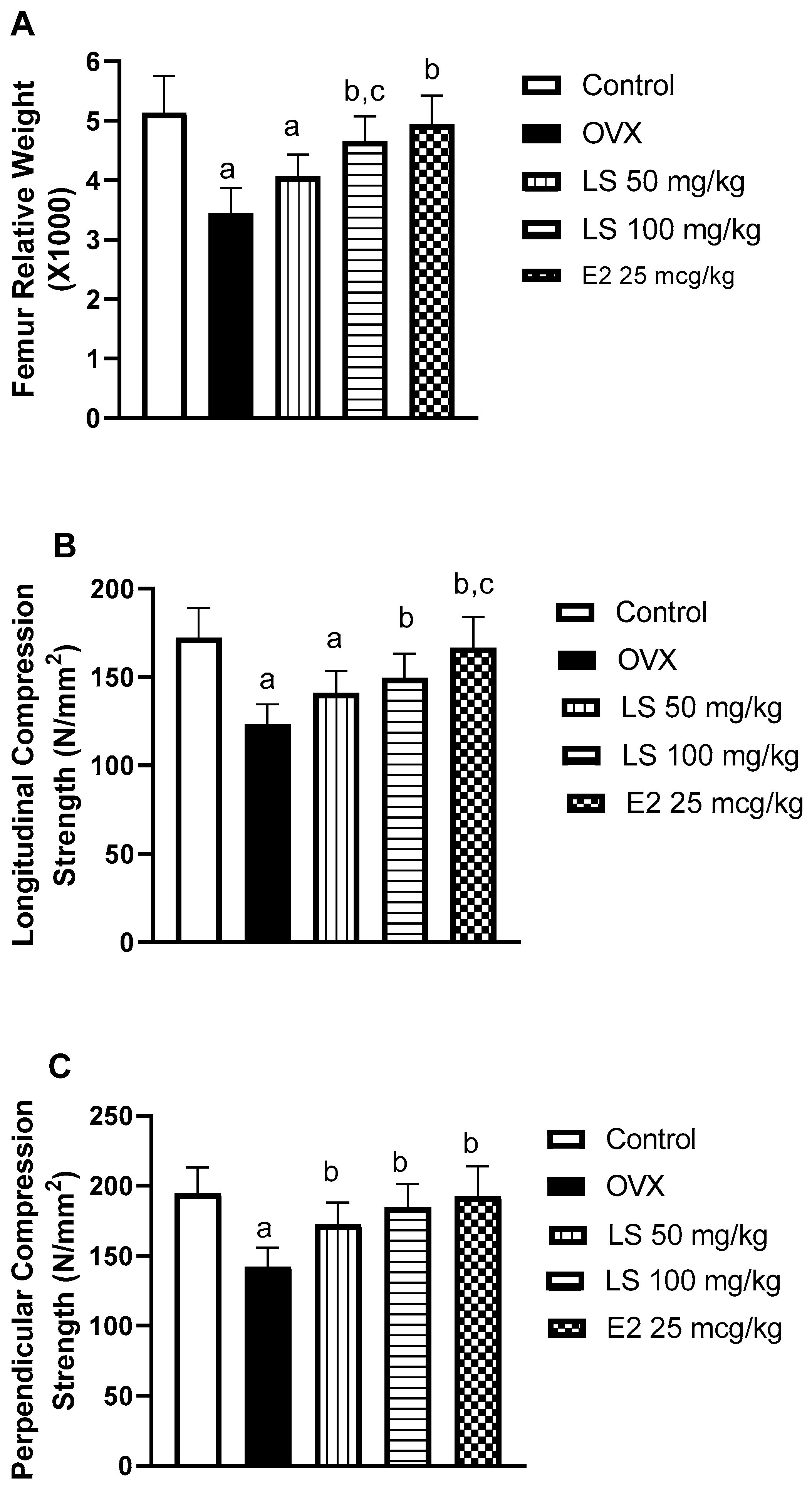
3.1.2. Effect of L. sativum on Femur Relative Weight and Compression Strength
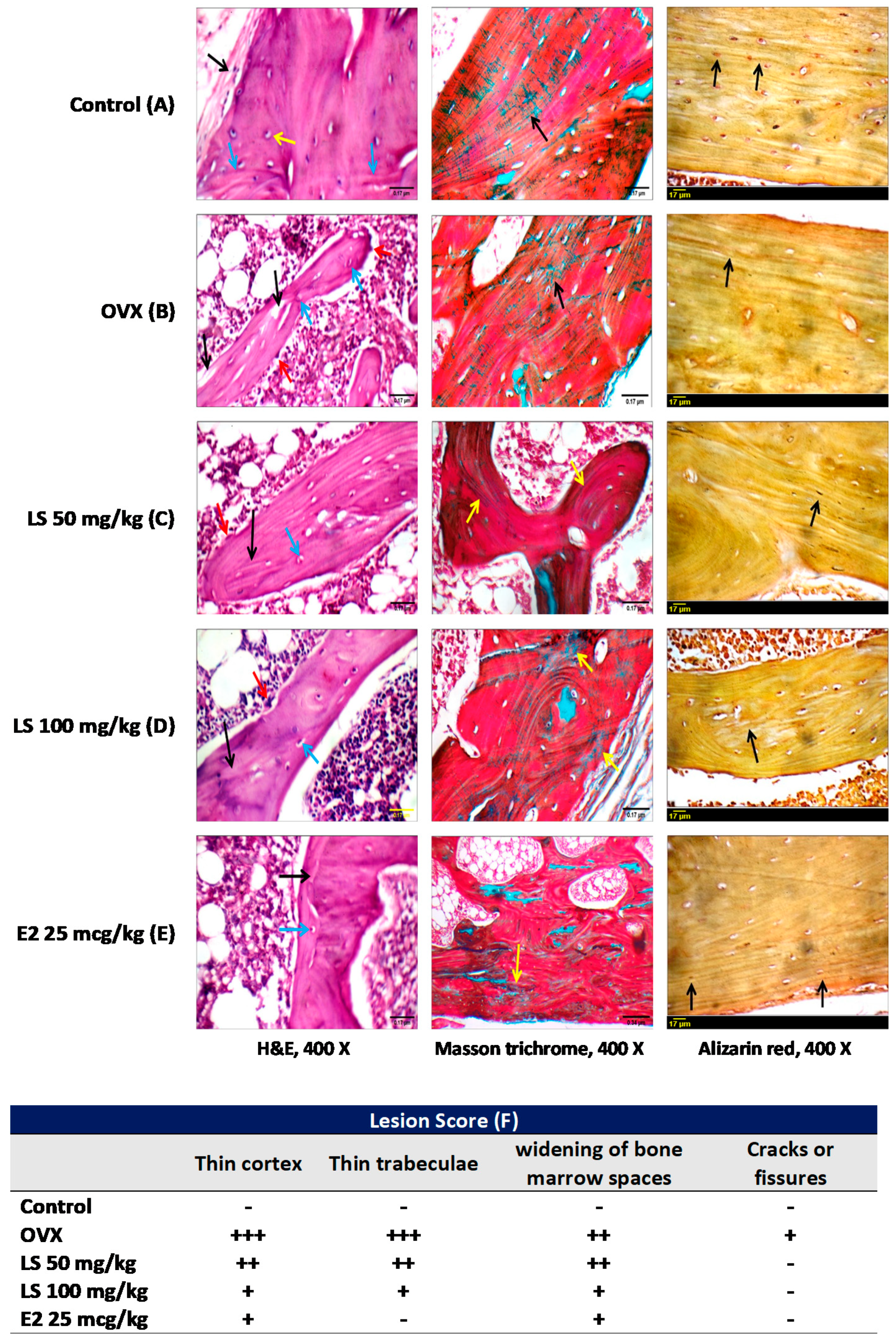
3.1.3. Histological Examination of Femurs
3.1.4. Effect of L. sativum on Serum Biomarkers of Bone Formation and Resorption
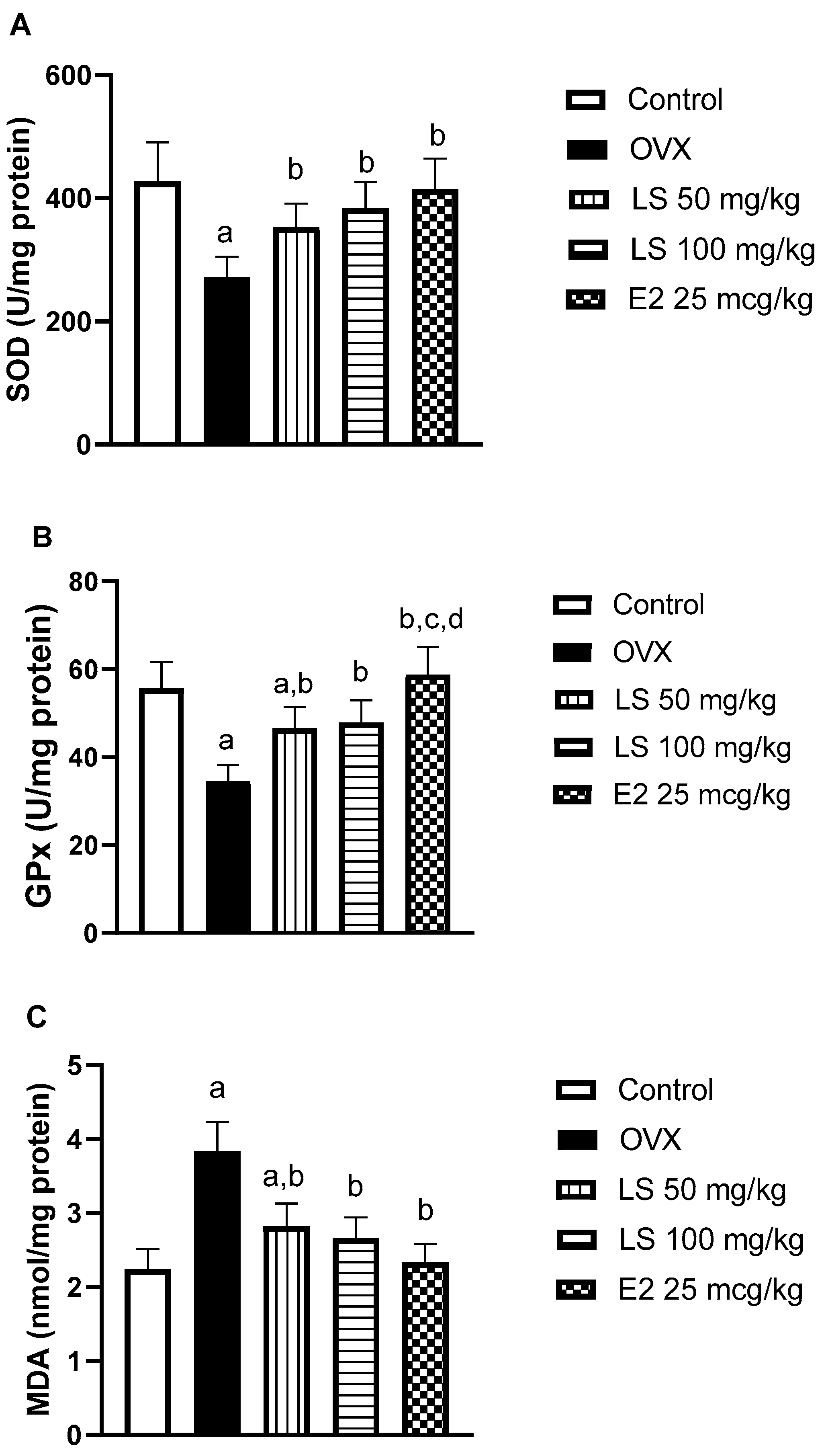
3.1.5. Effect of L. sativum on Ovariectomy-Induced Oxidative Stress in Bone Tissue
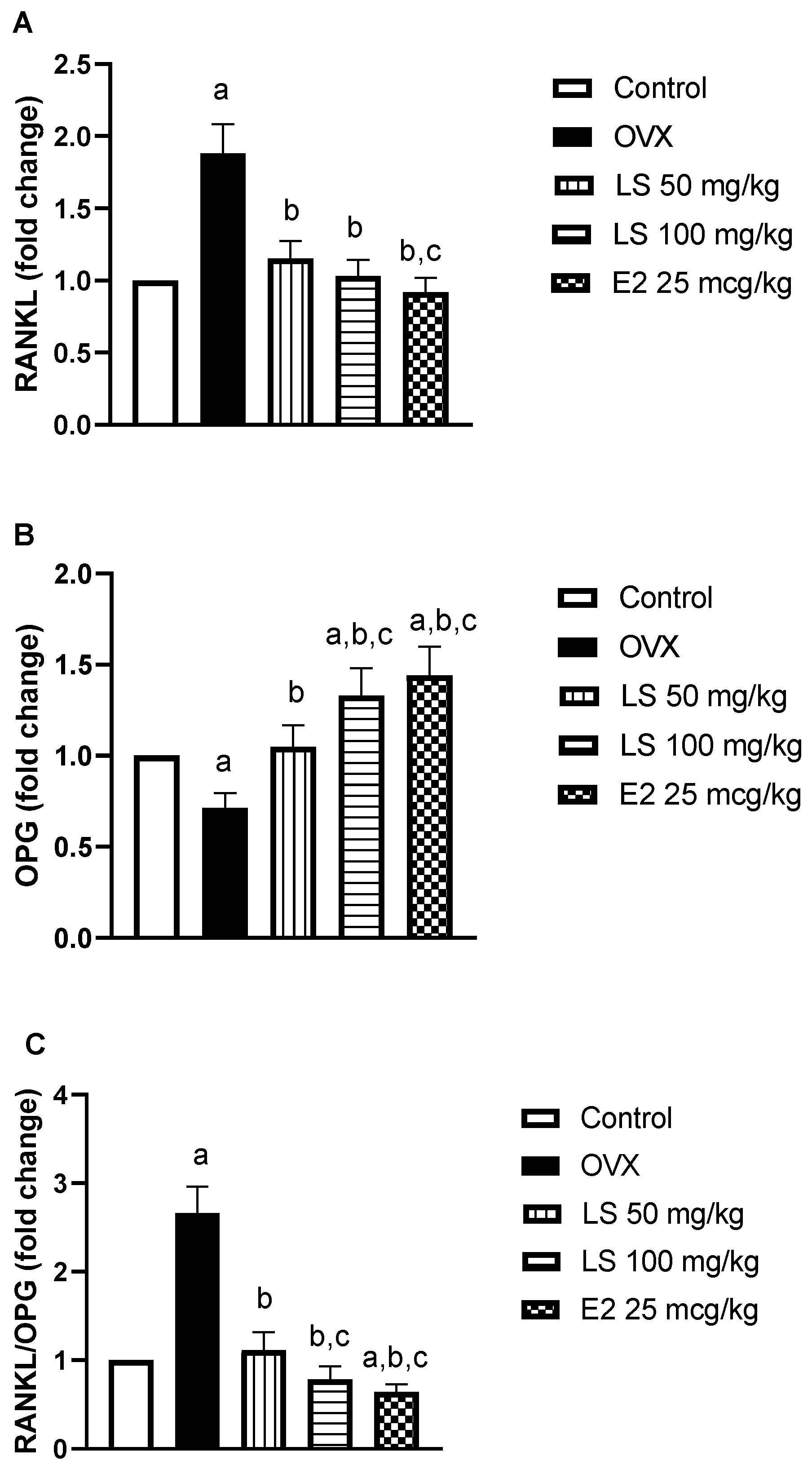
3.1.6. Effect of L. sativum on RANKL and OPG mRNA Expression
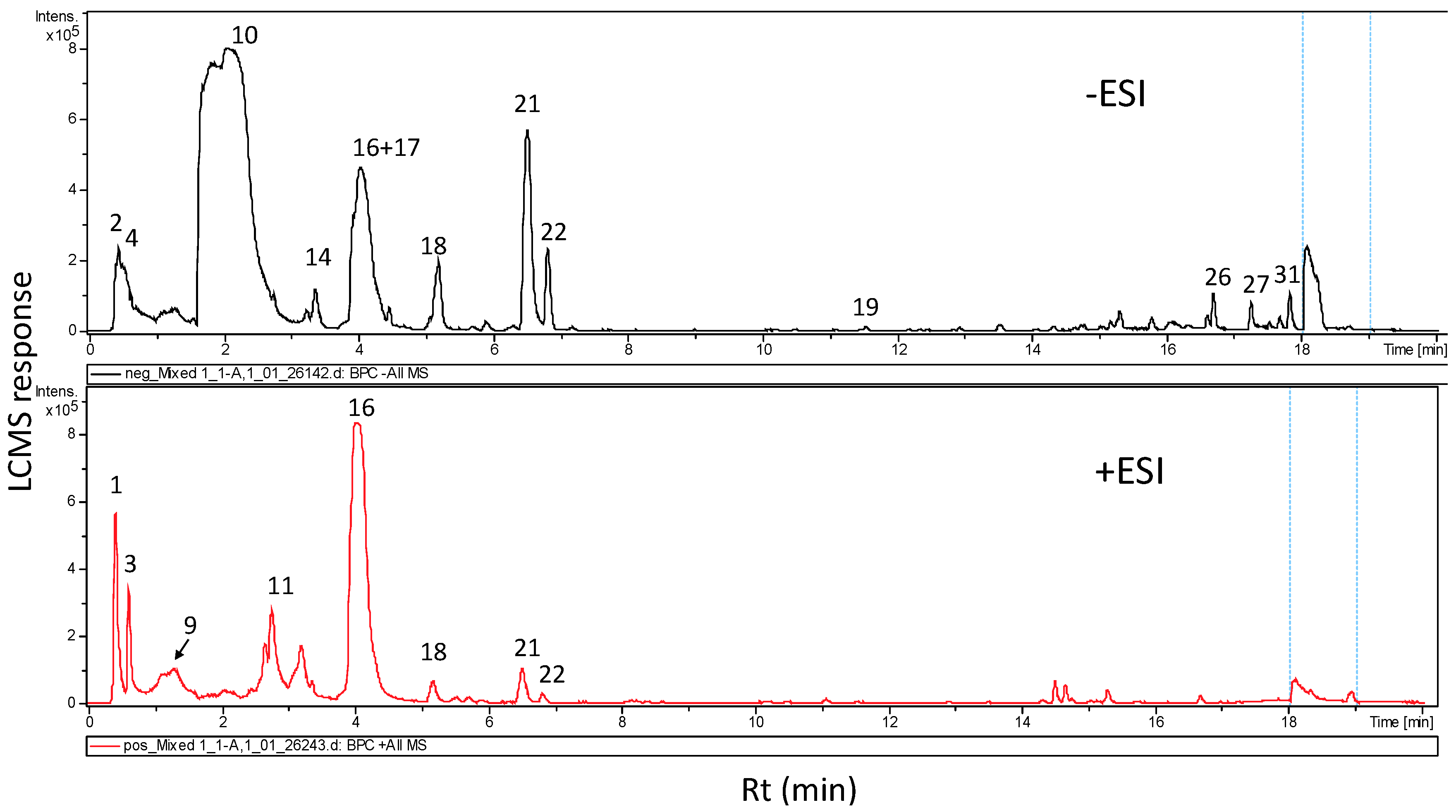
3.2. Metabolites Characterization of L. sativum Seed Extract via UPLC and GC/MS Analyses
3.2.1. Alkaloids
3.2.2. Glucosinolates
3.2.3. Phenolic Acids & Flavonoids/Lignans
3.2.4. Coumarins
3.2.5. Fatty Acids
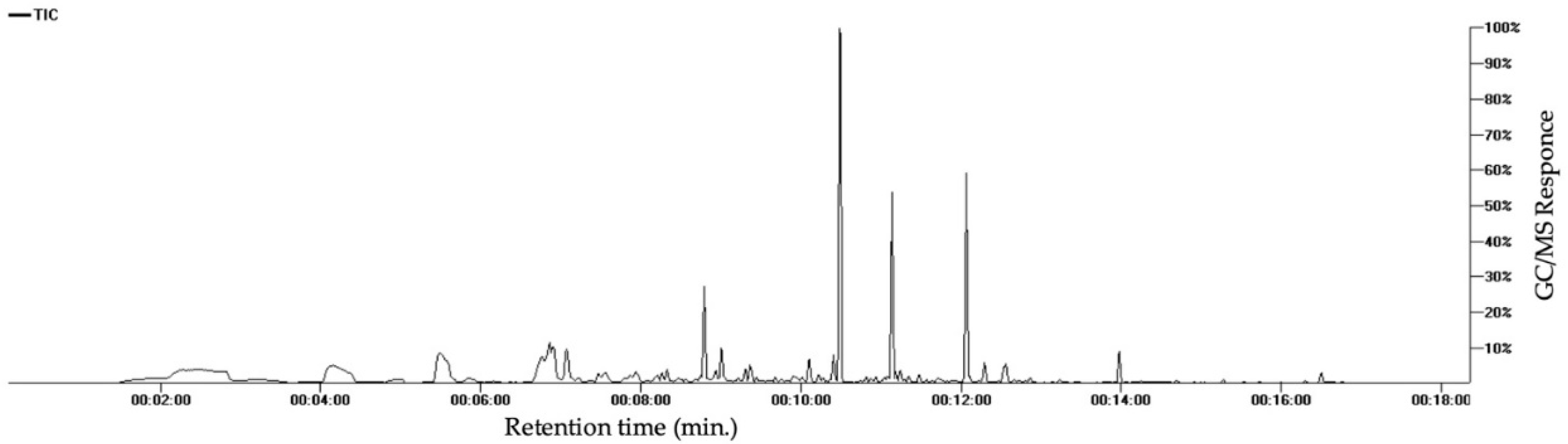
3.2.6. Volatile Compounds
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aghebati-Maleki, L.; Dolati, S.; Zandi, R.; Fotouhi, A.; Ahmadi, M.; Aghebati, A.; Nouri, M.; Kazem Shakouri, S.; Yousefi, M. Prospect of mesenchymal stem cells in therapy of osteoporosis: A review. J. Cell. Physiol. 2019, 234, 8570–8578. [Google Scholar] [CrossRef] [PubMed]
- Elshal, M.F.; Almalki, A.L.; Hussein, H.K.; Khan, J.A. Synergistic antiosteoporotic effect of Lepidium sativum and alendronate in glucocorticoid-induced osteoporosis in Wistar rats. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 267–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Yang, H.; Zhang, Q.; Liu, C.; Zhao, J.; Zhang, L.; Chen, B. Natural products for treatment of osteoporosis: The effects and mechanisms on promoting osteoblast-mediated bone formation. Life Sci. 2016, 147, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Caracchini, G.; Cavalli, L. Severe osteoporosis: Diagnosis of femoral fractures. Clin. Cases Miner. Bone Metab. 2010, 7, 97. [Google Scholar] [PubMed]
- Wu, C.-H.; Kao, I.-J.; Hung, W.-C.; Lin, S.-C.; Liu, H.-C.; Hsieh, M.-H.; Bagga, S.; Achra, M.; Cheng, T.-T.; Yang, R.-S. Economic impact and cost-effectiveness of fracture liaison services: A systematic review of the literature. Osteoporos. Int. 2018, 29, 1227–1242. [Google Scholar] [CrossRef]
- Peng, Y.-L.; Hu, H.-Y.; Luo, J.-C.; Hou, M.-C.; Lin, H.-C.; Lee, F.-Y. Alendronate, a bisphosphonate, increased upper and lower gastrointestinal bleeding: Risk factor analysis from a nationwide population-based study. Osteoporos. Int. 2014, 25, 1617–1623. [Google Scholar] [CrossRef] [PubMed]
- Russo, J.; Russo, I.H. The role of estrogen in the initiation of breast cancer. J. Steroid Biochem. Mol. Biol. 2006, 102, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horcajada, M.-N.; Offord, E. Naturally plant-derived compounds: Role in bone anabolism. Curr. Mol. Pharmacol. 2012, 5, 205–218. [Google Scholar] [CrossRef]
- Abdel-Naim, A.B.; Alghamdi, A.A.; Algandaby, M.M.; Al-Abbasi, F.A.; Al-Abd, A.M.; Eid, B.G.; Abdallah, H.M.; El-Halawany, A.M. Rutin isolated from Chrozophora tinctoria enhances bone cell proliferation and ossification markers. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Mossa, J.S.; Al-Said, M.S.; Al-Yahya, M.A. Medicinal plant diversity in the flora of Saudi Arabia 1: A report on seven plant families. Fitoterapia 2004, 75, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Vinti Dixit, J.R., III; Kumar, I.; Palandurkar, K.; Giri, R.; Giri, K. Lepidium sativum: Bone healer in traditional medicine, an experimental validation study in rats. J. Family Med. Prim. Care 2020, 9, 812. [Google Scholar]
- Maier, U.H.; Gundlach, H.; Zenk, M.H. Seven imidazole alkaloids from Lepidium sativum. Phytochemistry 1998, 49, 1791–1795. [Google Scholar] [CrossRef]
- Fan, Q.-L.; Zhu, Y.-D.; Huang, W.-H.; Qi, Y.; Guo, B.-L. Two new acylated flavonol glycosides from the seeds of Lepidium sativum. Molecules 2014, 19, 11341–11349. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmari, A.K.; Athar, M.T.; Al-Shahrani, H.M.; Al-Dakheel, S.I.; Al-Ghamdi, M.A. Efficacy of Lepidium sativum against carbon tetra chloride induced hepatotoxicity and determination of its bioactive compounds by GC/MS. Toxicol. Rep. 2015, 2, 1319–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Haroun, H.; Soliman, M.; El-Gawad, A. Comparative Study on the Possible Protective Effect of Lepidium Sativum versus Teriparatide in Induced Osteoporosis in Adult Male Guinea Pigs. Egypt. J. Histol. 2020. [Google Scholar] [CrossRef]
- bin Abdullah Juma, A.b.H. The effects of Lepidium sativum seeds on fracture-induced healing in rabbits. Medscape Gen. Med. 2007, 9, 23. [Google Scholar]
- OECD, O. Guide Lines for the Testing of Chemicals/Section 4: Health Effects Test No 423: Acute oral Toxicity–Acute Toxic Class Method; OECD: Paris, France, 2002. [Google Scholar]
- Balgoon, M.J. Assessment of the Protective Effect of Lepidium sativum against Aluminum-Induced Liver and Kidney Effects in Albino Rat. Biomed. Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Attia, E.S.; Amer, A.H.; Hasanein, M.A. The hypoglycemic and antioxidant activities of garden cress (Lepidium sativum L.) seed on alloxan-induced diabetic male rats. Nat. Prod. Res. 2019, 33, 901–905. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, Z.-X.; Zhang, Y.; Yan, Y.-B.; He, Q.; Cao, P.-C.; Lei, W. Anti-osteoporosis activity of Cibotium barometz extract on ovariectomy-induced bone loss in rats. J. Ethnopharmacol. 2011, 137, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Fayed, H.A.; Barakat, B.M.; Elshaer, S.S.; Abdel-Naim, A.B.; Menze, E.T. Antiosteoporotic activities of isoquercitrin in ovariectomized rats: Role of inhibiting hypoxia inducible factor-1 alpha. Eur. J. Pharmacol. 2019, 865, 172785. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Al-Abd, A.M.; Asaad, G.F.; Abdel-Naim, A.B.; El-Halawany, A.M. Isolation of antiosteoporotic compounds from seeds of Sophora japonica. PLoS ONE 2014, 9, e98559. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Ali, S.E.; Hodaya, R.H.; El-Seedi, H.R.; Sultani, H.N.; Laub, A.; Eissa, T.F.; Abou-Zaid, F.O.F.; Wessjohann, L.A. Phytochemical Profiles and Antimicrobial Activities of Allium cepa Red cv. and A. sativum Subjected to Different Drying Methods: A Comparative MS-Based Metabolomics. Molecules 2017, 22, 761. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; El-Kersh, D.M.; Ehrlich, A.; Choucry, M.A.; El-Seedi, H.; Frolov, A.; Wessjohann, L.A. Variation in Ceratonia siliqua pod metabolome in context of its different geographical origin, ripening stage and roasting process. Food Chem. 2019, 283, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Sakna, S.T.; Mocan, A.; Sultani, H.N.; El-Fiky, N.M.; Wessjohann, L.A.; Farag, M.A. Metabolites profiling of Ziziphus leaf taxa via UHPLC/PDA/ESI-MS in relation to their biological activities. Food Chem. 2019, 293, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; El-Kersh, D.M.; Rasheed, D.M.; Heiss, A.G. Volatiles distribution in Nigella species (black cumin seeds) and in response to roasting as analyzed via solid-phase microextraction (SPME) coupled to chemometrics. Ind. Crops Prod. 2017, 108, 564–571. [Google Scholar] [CrossRef]
- Watts, N.B. Postmenopausal osteoporosis: A clinical review. J. Womens Health 2018, 27, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, L.; Yao, M.; Cui, X.-J.; Xue, C.-C.; Wang, Y.-J.; Shu, B. Biomechanical characteristics of osteoporotic fracture healing in ovariectomized rats: A systematic review. PLoS ONE 2016, 11, e0153120. [Google Scholar] [CrossRef] [Green Version]
- El, N.K.; Hazzaa, H.; Hemoud, H.; Imam, O. Clinical and Radiographic Assessment of Single or Combined Treatment with Lepidium sativum and Alendronate of Non-Surgically Treated Chronic Periodontitis in Postmenopausal Osteoporotic Women. J. Int. Acad. Periodontol. 2019, 21, 20–28. [Google Scholar]
- Minkin, C. Bone acid phosphatase: Tartrate-resistant acid phosphatase as a marker of osteoclast function. Calcif. Tissue Int. 1982, 34, 285–290. [Google Scholar] [CrossRef]
- Selek, S.; Koyuncu, I.; Caglar, H.G.; Bektas, I.; Yilmaz, M.A.; Gonel, A.; Akyuz, E. The evaluation of antioxidant and anticancer effects of Lepidium Sativum Subsp Spinescens L. methanol extract on cancer cells. Cell. Mol. Biol. 2018, 64, 72–80. [Google Scholar] [CrossRef]
- Smolinska, B.; Szczodrowska, A. Antioxidative response of Lepidium sativum L. during assisted phytoremediation of Hg contaminated soil. New Biotech. 2017, 38, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Al-Sheddi, E.S.; Farshori, N.N.; Al-Oqail, M.M.; Musarrat, J.; Al-Khedhairy, A.A.; Siddiqui, M.A. Protective effect of Lepidium sativum seed extract against hydrogen peroxide-induced cytotoxicity and oxidative stress in human liver cells (HepG2). Pharm. Biol. 2016, 54, 314–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Zhu, L.; Zhang, D.; Li, N.; Li, Q.; Dai, P.; Mao, Y.; Li, X.; Ma, J.; Huang, S. Oxidative stress-related biomarkers in postmenopausal osteoporosis: A systematic review and meta-analyses. Dis. Markers 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorsi, G.; Piva, I.; Greco, P.; Cervellati, C. Oxidative stress as a possible pathogenic cofactor of post-menopausal osteoporosis: Existing evidence in support of the axis oestrogen deficiency-redox imbalance-bone loss. Indian J. Med. Res. 2018, 147, 341. [Google Scholar]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [Green Version]
- Altindag, O.; Erel, O.; Soran, N.; Celik, H.; Selek, S. Total oxidative/anti-oxidative status and relation to bone mineral density in osteoporosis. Rheumatol. Int. 2008, 28, 317–321. [Google Scholar] [CrossRef]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.-C.; Lu, D.; Liu, A.-L.; Zhang, Z.-M.; Li, X.-M.; Zou, Z.-P.; Zeng, W.-S.; Cheng, B.-L.; Luo, S.-Q. Reactive oxygen species stimulates receptor activator of NF-κB ligand expression in osteoblast. J. Biol. Chem. 2005, 280, 17497–17506. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-K.; Yang, L.; Meng, G.-L.; Fan, J.; Chen, J.-Z.; He, Q.-Z.; Chen, S.; Fan, J.-Z.; Luo, Z.-J.; Liu, J. Protective effect of tetrahydroxystilbene glucoside against hydrogen peroxide-induced dysfunction and oxidative stress in osteoblastic MC3T3-E1 cells. Eur. J. Pharmacol. 2012, 689, 31–37. [Google Scholar] [CrossRef]
- Martin, T.J. Historically significant events in the discovery of RANK/RANKL/OPG. World J. Orthop. 2013, 4, 186. [Google Scholar] [CrossRef]
- Azizieh, F.Y.; Shehab, D.; Jarallah, K.A.; Gupta, R.; Raghupathy, R. Circulatory Levels of RANKL, OPG, and Oxidative Stress Markers in Postmenopausal Women With Normal or Low Bone Mineral Density. Biomark. Insights 2019, 14, 1177271919843825. [Google Scholar] [CrossRef] [PubMed]
- Imade, O.V.; Erinfolami, W.A.; Ajadi, R.A.; Abioja, M.O.; Rahman, S.A.; Smith, O.F.; Gazal, O.S. Effects of Lepidium sativum supplementation on growth and gonadotropins secretion in ovariectomized, estrogen-implanted rabbits. Asian Pac. J. Reprod. 2018, 7, 155. [Google Scholar] [CrossRef]
- Ghante, M.H.; Badole, S.L.; Bodhankar, S.L. Health benefits of garden cress (Lepidium sativum Linn.) seed extracts. InNuts and seeds in health and disease prevention. Academic Press. 2011, 521–525. [Google Scholar] [CrossRef]
- Oszmianski, J.; Kolniak-Ostek, J.; Wojdylo, A. Application of ultra performance liquid chromatography-photodiode detector-quadrupole/time of flight-mass spectrometry (UPLC-PDA-Q/TOF-MS) method for the characterization of phenolic compounds of Lepidium sativum L. sprouts. Eur. Food Res. Technol. 2013, 236, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Piacente, S.; Carbone, V.; Plaza, A.; Zampelli, A.; Pizza, C. Investigation of the tuber constituents of maca (Lepidium meyenii Walp.). J. Agric. Food Chem. 2002, 50, 5621–5625. [Google Scholar] [CrossRef]
- Sansom, C.E.; Jones, V.S.; Joyce, N.I.; Smallfield, B.M.; Perry, N.B.; van Klink, J.W. Flavor, glucosinolates, and isothiocyanates of Nau (Cook’s scurvy grass, Lepidium oleraceum) and other rare New Zealand Lepidium species. J. Agric. Food Chem. 2015, 63, 1833–1838. [Google Scholar] [CrossRef]
- Wang, J.; Yu, H.; Zhao, Z.; Sheng, X.; Shen, Y.; Gu, H. Natural Variation of Glucosinolates and Their Breakdown Products in Broccoli (Brassica oleracea var. italica) Seeds. J. Agric. Food Chem. 2019, 67, 12528–12537. [Google Scholar] [CrossRef]
- Wang, S.; Shi, P.; Qu, L.; Ruan, J.; Yang, S.; Yu, H.; Zhang, Y.; Wang, T. Bioactive constituents obtained from the seeds of Lepidium apetalum Willd. Molecules 2017, 22, 540. [Google Scholar] [CrossRef] [Green Version]
- Hijazin, T.; Radwan, A.; Abouzeid, S.; Dräger, G.; Selmar, D. Uptake and modification of umbelliferone by various seedlings. Phytochemistry 2019, 157, 194–199. [Google Scholar] [CrossRef]
- Alqahtani, F.Y.; Aleanizy, F.S.; Mahmoud, A.Z.; Farshori, N.N.; Alfaraj, R.; Al-Sheddi, E.S.; Alsarra, I.A. Chemical composition and antimicrobial, antioxidant, and anti-inflammatory activities of Lepidium sativum seed oil. Saudi J. Biol. Sci. 2019, 26, 1089–1092. [Google Scholar] [CrossRef]
- Kerwin, J.L.; Torvik, J.J. Identification of monohydroxy fatty acids by electrospray mass spectrometry and tandem mass spectrometry. J. Anal. Biochem. 1996, 237, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Martin-Arjol, I.; Bassas-Galia, M.; Bermudo, E.; Garcia, F.; Manresa, A. Identification of oxylipins with antifungal activity by LC-MS/MS from the supernatant of Pseudomonas 42A2. Chem. Phys. Lipids 2010, 163, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Park, H.; Hyun, H.; Kim, J.; Kim, H.; Oh, H.I.; Hwang, H.S.; Kim, D.K.; Kim, H.H. Effects of Glucosinolates from Turnip (Brassica rapa L.) Root on Bone Formation by Human Osteoblast-Like MG-63 Cells and in Normal Young Rats. Phytother. Res. 2015, 29, 902–909. [Google Scholar] [CrossRef]
- Folwarczna, J.; Zych, M.; Burczyk, J.; Trzeciak, H.; Trzeciak, H.I. Effects of natural phenolic acids on the skeletal system of ovariectomized rats. Planta Med. 2009, 75, 1567–1572. [Google Scholar] [CrossRef]
- Zhao, X.-L.; Chen, L.-F.; Wang, Z. Aesculin modulates bone metabolism by suppressing receptor activator of NF-κB ligand (RANKL)-induced osteoclastogenesis and transduction signals. Biochem. Biophys. Res. Commun. 2017, 488, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Vanek, C.; Connor, W.E. Do n−3 Fatty Acids Prevent Osteoporosis? Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Van Papendorp, D.; Coetzer, H.; Kruger, M. Biochemical profile of osteoporotic patients on essential fatty acid supplementation. Nutr. Res. 1995, 15, 325–334. [Google Scholar] [CrossRef]
- Pankova, S.; Tsvetkova, D. Role of phytoestrogens in prevention of osteoporosis. Int. J. Curr. Pharm. Res. 2015, 7, 1–6. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALP (U/dL) | Osteocalcin (pg/mL) | TRAP (U/dL) | CTXI (ng/mL) | |
|---|---|---|---|---|
| Control | 28.65 ± 2.71 | 4.25 ± 0.45 | 2.25 ± 0.32 | 3.25 ± 0.47 |
| OVX | 21.33 ± 2.83 a | 2.68 ± 0.28 a | 4.10 ± 0.45 a | 5.47 ± 0.58 a |
| LS 50 mg/kg | 25.74 ± 2.88 | 3.15 ± 0.32 a | 3.27 ± 0.34 a | 4.10 ± 0.47 a,b |
| LS 100 mg/kg | 27.61 ± 2.80 b | 3.66 ± 0.38 b | 2.94 ± 0.25 a,b | 3.85 ± 0.40 b |
| E2 25 mcg/kg | 28.47 ± 2.90 b | 4.10 ± 0.43 b,c | 2.44 ± 0.32 b,c | 3.55 ± 0.42 b |
| Peak No | RT (min) | UV (nm) | Mol. Ion | Mol. Formula | Name | Class | Error (ppm) | MSMS Fragments |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.3 | Nd | 705.1815 | C36H35O15+ | Hexahydroxy-biflavanone-O-hexoside | Flavonoid | −0.1 | 543 |
| 2 | 0.35 | Nd | 325.1135 | C12H21O10+ | Unknown disaccharide | Sugar | 2.3 | 223, 127 |
| 3 | 0.5 | 280 | 337.1384 | C16H21N2O6+ | Semilepidinoside A | Glycoalkaloid | 0.9 | 347, 174 |
| 4 | 0.52 | Nd | 719.2016 | C37H37O15+ | Dimethoxycinnamoyl-O-feruloyl-O-caffeoylquinic acid | Phenolic acid | −5.4 | 539, 377, 341 |
| 5 | 0.5 | Nd | 723.1882 | C36H37O16+ | Tetrahydroxyflavan-tetrahydroxyflavanone hexoside | Flavonoid | 4.9 | |
| 6 | 0.57 | Nd | 539. 1378 | C24H27O14− | Unknown | Sugar | 1.4 | 377 |
| 7 | 0.6 | 280 | 337.1384 | C16H21N2O6+ | Semilepidinoside A (isomer) | Glycoalkaloid | 2.9 | 174 |
| 8 | 0.7 | Nd | 337.1409 | C12H17O8+ | Aconitic acid-O-hexoside | Organic acid | 7.7 | 175 |
| 9 | 1.1 | 280 | 347.1476 | C20H19N4O2+ | Lepidine (B/D/E/F) | Alkaloid | 7.4 | 174 |
| 10 | 1.9 | 280 | 408.0463 | C14H19NO9S2− | Benzyl-glucosinolate (Glucotropaeolin) | Glucosinolate | - | 246, 212, 96 |
| 11 | 2.3 | 280 | 359.1525 | C20H23O6− | Lariciresinol | Lignan | 3.1 | 345, 298 |
| 12 | 2.6 | 280 | 361.1648 | C21H21N4O2+ | Lepidine C | Alkaloid | 3 | 174 |
| 13 | 2.9 | Nd | 347.167 | C20H19N4O2+ | Lepidine (B/D/E/F) | Alkaloid | ||
| 14 | 3.2 | 280 | 361.167 | C21H21N4O2+ | Lepidine C | Alkaloid | 3.0 | 214 |
| 15 | 3.4 | 280.5 | 425.0897 | C22H17O9− | Unknown | Flavonoid | −4.5 | 359 |
| 16 | 3.9 | 328 | 310.1657 | C16H25NO5+ | Sinapine | Alkaloid | 0.6 | 251, 207 |
| 17 | 4.1 | 328 | 294.1348 | - | Unknown | Glucosinolate | 96 | |
| 18 | 5.0 | Nd | 369.1191 | C17H21O9+ | O-Caffeoylquinic acid methyl ester | Phenolic acid | −2.5 | |
| 19 | 5.8 | Nd | 225.0745 | C11H13O5+ | Trihydroxycinnamic acid-O-di-methyl ester | Phenolic acid | 5.3 | 175 |
| 20 | 6.1 | 328 | 207.0642 | C11H9O4− | Dimethylesculetin | Coumarin | 0.4 | 175 |
| 21 | 6.5 | 329 | 339.0722 | C15H15O9 | Esculin | Coumarin | −4.6 | 223, 179 |
| 22 | 6.9 | 327 | 613.1870 | C26H33N2O15+ | Unknown | |||
| 23 | 11.6 | 326 | 591.1684 | C28H31O14− | Acacetin-O-rutinoside | Flavonoid | 5.9 | 283 |
| 24 | 12.2 | Nd | 329.2319 | C18H33O5− | Trihydroxy octadecenoic acid | Hydroxy fatty acid | 4.4 | 283 |
| 25 | 16.5 | Nd | 271.2299 | C16H31O3− | Hydroxy hexadecanoic acid | Hydroxy fatty acid | 3.5 | 225, 116 |
| 26 | 16.6 | Nd | 277.2173 | C18H29O2− | Linolenic acid | Unsaturated fatty acid | 4.5 | 251, 211 |
| 27 | 17.2 | Nd | 279.2330 | C18H31O2- | Linoleic acid | Unsaturated fatty acid | 4 | 211 |
| 28 | 17.3 | Nd | 425.287+ | C24H41O6+ | Unknown steroid | Steroid | 3.3 | |
| 29 | 17.5 | Nd | 327.2888 | C20H39O3− | Hydroxy eicosanoic acid | Hydroxy fatty acid | 5.1 | - |
| 30 | 17.7 | Nd | 353.3044 | C22H41O3− | Oxo docosanoic acid | Oxygenated fatty acid | 4.9 | - |
| 31 | 17.8 | Nd | 281.2488 | C18H33O2− | Oleic acid | Unsaturated fatty acid | −0.7 | |
| 32 | 18.8 | Nd | 311.2941 | C20H39O2+ | Eicosenoic acid | Unsaturated fatty acid | 1.2 |
| No. | CAS no. | Name | Class | RT | KI | Expected KI | % |
|---|---|---|---|---|---|---|---|
| 1 | 64-19-7 | Acetic acid | Acid | 2.811 | 591 | 610 | 1.95 |
| 2 | 111-27-3 | 1-Hexanol | Alcohol | 5.473 | 843 | 868 | 12.21 |
| 3 | 142-62-1 | Caproic acid | Acid | 6.8486 | 973 | 990 | 1.70 |
| 4 | 72237-36-6 | 4-Hexenyl acetate | Ester | 7.2139 | 1008 | 1020 | 0.69 |
| 5 | 535-77-3 | m-Cymene | Monoterpene hydrocarbon | 7.4558 | 1030 | 1023 | 1.29 |
| 6 | 100-51-6 | Benzyl alcohol | Aromatic | 7.5425 | 1038 | 1036 | 1.21 |
| 7 | 111-87-5 | 1-Octanol | Alcohol | 7.9197 | 1073 | 1071 | 1.41 |
| 8 | 30086-02-3 | 3,5-Octadien-2-one | Ketone | 7.9493 | 1076 | 1073 | 0.84 |
| 9 | 99-86-5 | α-Terpinene | Monoterpene hydrocarbon | 8.1916 | 1099 | 1017 | 0.59 |
| 10 | 1120-21-4 | Undecane | Aliphatic hydrocarbon | 8.2512 | 1104 | 1100 | 0.99 |
| 11 | 124-19-6 | Nonanal | Aldehyde | 8.3168 | 1110 | 1104 | 1.21 |
| 14 | 140-29-4 | Benzyl cyanide | Nitrogenous | 8.7354 | 1150 | 1144 | 0.97 |
| 16 | 124-07-2 | Caprylic acid | Acid | 8.924 | 1167 | 1180 | 0.91 |
| 17 | 143-08-8 | 1-Nonanol | Alcohol | 8.9942 | 1174 | 1173 | 3.78 |
| 19 | 91-20-3 | Naphthalene | Aromatic | 9.2767 | 1200 | 1182 | 0.21 |
| 20 | 112-40-3 | Dodecane | Aliphatic hydrocarbon | 9.2915 | 1202 | 1200 | 1.21 |
| 21 | 140-67-0 | Estragole | Ether | 9.3507 | 1208 | 1196 | 1.92 |
| 22 | 112-31-2 | Decanal | Aldehyde | 9.3689 | 1210 | 1206 | 0.85 |
| 23 | 17301-23-4 | 2,6-Dimethylundecane | Aliphatic hydrocarbon | 9.4331 | 1217 | 1210 | 0.41 |
| 28 | 112-05-0 | Nonanoic acid | Acid | 9.8915 | 1265 | 1273 | 0.59 |
| 30 | 14371-10-9 | Cinnamaldehyde | Aldehyde | 10.0882 | 1286 | 1270 | 2.65 |
| 31 | 1197-15-5 | Alpha-Terpinen-7-al | Aldehyde | 10.2019 | 1298 | 1283 | 0.45 |
| 32 | 629-50-5 | Tridecane | Aliphatic hydrocarbon | 10.2611 | 1304 | 1300 | 0.29 |
| 33 | 104-54-1 | Cinnamyl alcohol | Alcohol | 10.3897 | 1318 | 1312 | 3.23 |
| 36 | 80-26-2 | α-Terpinyl acetate | Ester | 10.8007 | 1361 | 1350 | 0.49 |
| 37 | 104-61-0 | γ-Nonalactone | Lactone | 10.9187 | 1373 | 1363 | 0.42 |
| 38 | 622-78-6 | Benzyl Isothiocyanate | Nitrogenous | 10.9927 | 1381 | 1364 | 0.15 |
| 39 | 77-68-9 | 1,3-Pentanediol, 2, 2,4-trimethyl-, 1-isobutyrate | Ester | 11.0303 | 1385 | 1380 | 0.48 |
| 40 | 22469-52-9 | Cyclosativene | Sesquiterpene hydrocarbon | 11.065 | 1389 | 1368 | 0.46 |
| 41 | 3856-25-5 | Alpha-Copaene | Sesquiterpene hydrocarbon | 11.1217 | 1395 | 1376 | 22.70 |
| 42 | 629-59-4 | Tetradecane | Aliphatic hydrocarbon | 11.1699 | 1400 | 1400 | 1.00 |
| 43 | 3650-28-0 | Sativene | Sesquiterpene hydrocarbon | 11.3348 | 1416 | 1396 | 0.53 |
| 44 | 475-20-7 | Longifolene | Sesquiterpene hydrocarbon | 11.4616 | 1428 | 1405 | 0.85 |
| 45 | 87-44-5 | Caryophyllene | Sesquiterpene hydrocarbon | 11.5517 | 1437 | 1419 | 0.13 |
| 46 | 91-64-5 | Coumarin | Lactone | 11.6997 | 1451 | 1441 | 0.41 |
| 47 | - | Unknown 6 | Aromatic | 12.0477 | 1485 | - | 16.60 |
| 48 | 629-62-9 | Pentadecane | Aliphatic hydrocarbon | 12.0883 | 1489 | 1500 | 0.56 |
| 49 | 10208-80-7 | α-Muurolene | Sesquiterpene hydrocarbon | 12.2757 | 1507 | 1440 | 2.82 |
| 50 | 39029-41-9 | g-Cadinene | Sesquiterpene hydrocarbon | 12.4499 | 1524 | 1513 | 0.08 |
| 51 | 483-76-1 | δ-Cadinene | Sesquiterpene hydrocarbon | 12.518 | 1531 | 1524 | 2.12 |
| 52 | 483-77-2 | Calamenene | Sesquiterpene hydrocarbon | 12.5412 | 1533 | 1523 | 2.23 |
| 53 | 29837-12-5 | Cubenene | Sesquiterpene hydrocarbon | 12.6469 | 1543 | 1532 | 0.25 |
| 54 | 21391-99-1 | α-Calacorene | Sesquiterpene hydrocarbon | 12.7865 | 1557 | 1542 | 0.18 |
| 55 | 629-73-2 | Cetene | Sesquiterpene hydrocarbon | 13.1248 | 1590 | 1592 | 0.09 |
| 56 | 544-76-3 | Hexadecane | Aliphatic hydrocarbon | 13.2144 | 1598 | 1600 | 0.35 |
| 57 | 119-61-9 | Benzophenone | Ketone | 13.7904 | 1662 | 1635 | 0.09 |
| 59 | - | Unknown 8 | Unknown | 13.9587 | 1680 | - | 3.49 |
| 60 | 544-63-8 | Myristic acid | Acid | 14.6776 | 1759 | 1768 | 0.19 |
| 61 | 120-51-4 | Benzyl Benzoate | Ester | 14.8996 | 1784 | 1762 | 0.05 |
| 63 | 1002-84-2 | Pentadecanoic acid | Acid | 15.5098 | 1848 | 1867 | 0.10 |
| 65 | 373-49-9 | Palmitoleic acid | Acid | 16.2828 | 1929 | 1951 | 0.12 |
| 66 | 57-10-3 | Palmitic acid | Acid | 16.4798 | 1949 | 1968 | 1.51 |
| 100.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdallah, H.M.; Farag, M.A.; Algandaby, M.M.; Nasrullah, M.Z.; Abdel-Naim, A.B.; Eid, B.G.; Safo, M.K.; Koshak, A.E.; Malebari, A.M. Osteoprotective Activity and Metabolite Fingerprint via UPLC/MS and GC/MS of Lepidium sativum in Ovariectomized Rats. Nutrients 2020, 12, 2075. https://doi.org/10.3390/nu12072075
Abdallah HM, Farag MA, Algandaby MM, Nasrullah MZ, Abdel-Naim AB, Eid BG, Safo MK, Koshak AE, Malebari AM. Osteoprotective Activity and Metabolite Fingerprint via UPLC/MS and GC/MS of Lepidium sativum in Ovariectomized Rats. Nutrients. 2020; 12(7):2075. https://doi.org/10.3390/nu12072075
Chicago/Turabian StyleAbdallah, Hossam M., Mohamed A. Farag, Mardi M. Algandaby, Mohammed Z. Nasrullah, Ashraf B. Abdel-Naim, Basma G. Eid, Martin K. Safo, Abdulrahman E. Koshak, and Azizah M. Malebari. 2020. "Osteoprotective Activity and Metabolite Fingerprint via UPLC/MS and GC/MS of Lepidium sativum in Ovariectomized Rats" Nutrients 12, no. 7: 2075. https://doi.org/10.3390/nu12072075