The Gut Microbiome and the Big Eight


Abstract
:1. Introduction
2. We Are What We Eat: Diet and the Microbiome
3. Just One Bite, or Maybe Two: The Mechanisms of Food Allergy
4. “Good Source of Protein?”: Interactions of Food Matrices and Gut Permeability
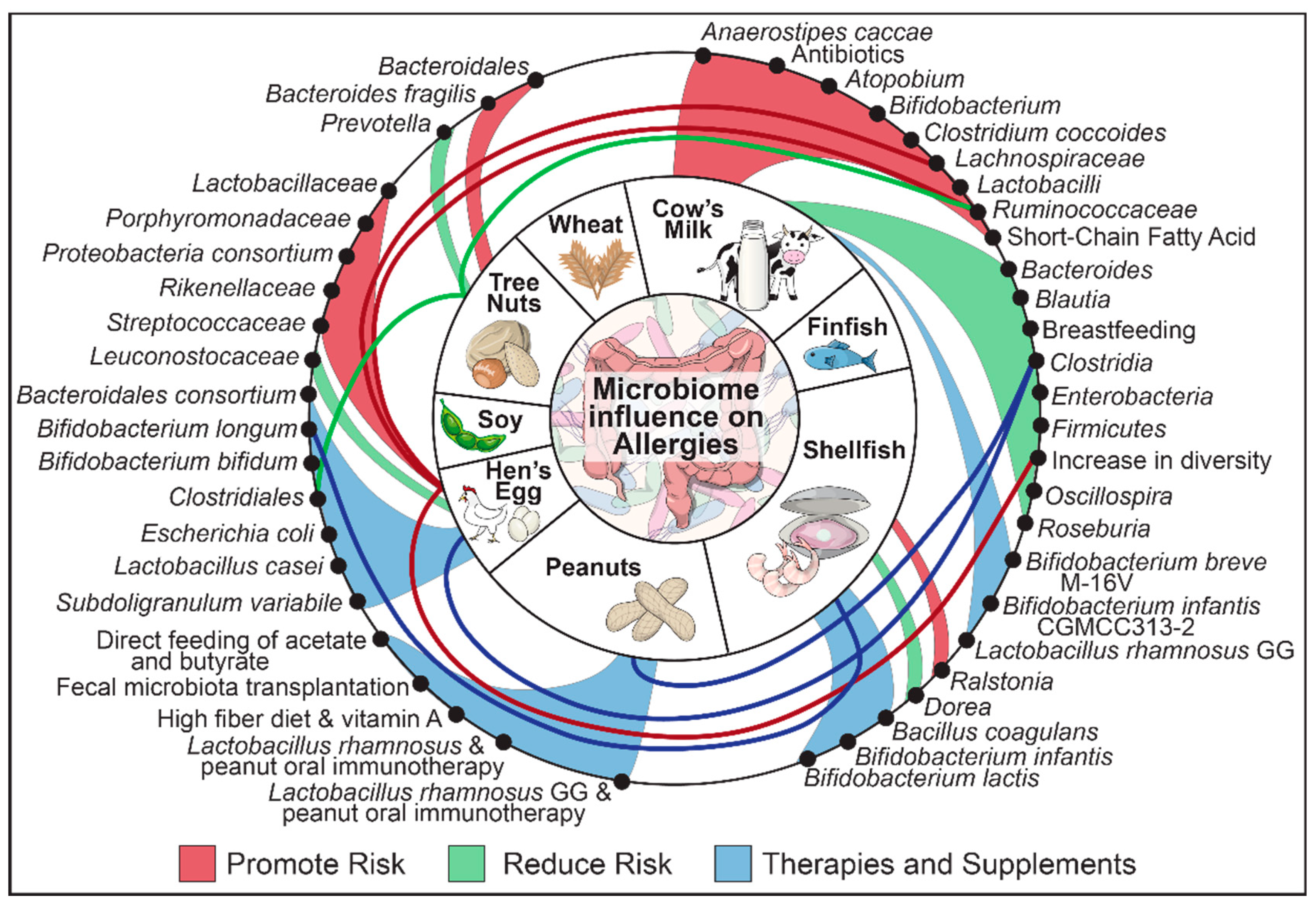
5. The Gut, The Big Eight and the Correlation between the Two
5.1. Cow Milk
5.2. Hen’s Egg
5.3. Peanut
5.4. Tree Nuts
5.5. Crustacean and Molluscan Shellfish
5.6. Wheat
5.7. Soy
5.8. Finfish
5.9. Common or Unique—Summary of Work
6. There Is More Than Meets the Gut: Gaps and Future Perspectives
6.1. Viral and Fungal Interactions
6.2. Clostridiales Cocktail
6.3. Exparimental Design and Analysis
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenta, R.; Hochwallner, H.; Linhart, B.; Pahr, S. Food allergies: The basics. Gastroenterology 2015, 148, 1120–1131.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contijoch, E.J.; Britton, G.J.; Yang, C.; Mogno, I.; Li, Z.; Ng, R.; Llewellyn, S.R.; Hira, S.; Johnson, C.; Rabinowitz, K.M.; et al. Gut microbiota density influences host physiology and is shaped by host and microbial factors. eLife 2019, 8, e40553. [Google Scholar] [CrossRef] [PubMed]
- Voreades, N.; Kozil, A.; Weir, T.L. Diet and the development of the human intestinal microbiome. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Venegas, D.P.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Noval Rivas, M.; Burton, O.T.; Wise, P.; Zhang, Y.Q.; Hobson, S.A.; Garcia Lloret, M.; Chehoud, C.; Kuczynski, J.; Desantis, T.; Warrington, J.; et al. A microbiota signature associated with experimental food allergy promotes allergic sensitization and anaphylaxis. J. Allergy Clin. Immunol. 2013, 131, 201–212. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Mattila, P.; Joenväärä, S.; Renkonen, J.; Toppila-Salmi, S.; Renkonen, R. Allergy as an epithelial barrier disease. Clin. Transl. Allergy 2011, 1, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hirata, S.; Kunisawa, J. Gut microbiome, metabolome, and allergic diseases. Allergol. Int. 2017, 66, 523–528. [Google Scholar] [CrossRef]
- Smolinska, S.; Jutel, M.; Crameri, R.; O’Mahony, L. Histamine and gut mucosal immune regulation. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary Fiber and Bacterial SCFA Enhance Oral Tolerance and Protect against Food Allergy through Diverse Cellular Pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, H.A.; O’mahony, L.; Burks, A.W.; Plaut, M.; Lack, G.; Akdis, C.A. Mechanisms of food allergy. J. Allergy Clin. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, A.; Ramachandran, S.; Smith, G.P.; Stevenson, M.L.; Pomeranz, M.K.; Cohen, D.E. Oral allergy syndrome (Pollen-food allergy syndrome). Dermatitis 2015, 26, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [Green Version]
- Verhoeckx, K.C.M.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheney, C.; Holzhauser, T.; Shimojo, R.; van der Bolt, N.; et al. Food processing and allergenicity. Food Chem. Toxicol. 2015, 80, 223–240. [Google Scholar] [CrossRef]
- Nowak-Wegrzyn, A.; Fiocchi, A. Rare, medium, or well done? The effect of heating and food matrix on food protein allergenicity. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 234–237. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Jansen, K.; Głobińska, A.; van de Veen, W.; Akdis, M. Regulatory Immune Mechanisms in Tolerance to Food Allergy. Front. Immunol. 2018, 9, 2939. [Google Scholar] [CrossRef]
- Allergy, M.P. Milk Protein Allergy. Refer. Mod. Food Sci. 2015, 1–5. [Google Scholar] [CrossRef]
- Caubet, J.C.; Wang, J. Current Understanding of Egg Allergy. Pediatr. Clin. North Am. 2011, 58, 427–443. [Google Scholar] [CrossRef] [Green Version]
- Mueller, G.A.; Maleki, S.J.; Pedersen, L.C. The molecular basis of peanut allergy topical collection on allergens. Curr. Allergy Asth. Rep. 2014, 14, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiselhart, S.; Hoffmann-Sommergruber, K.; Bublin, M. Tree nut allergens. Mol. Immunol. 2018, 100, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Lopata, A.L.; O’Hehir, R.E.; Lehrer, S.B. Shellfish allergy. Clin. Exp. Allergy 2010, 40, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Swoboda, I.; Arumugam, K.; Hilger, C.; Hentges, F. Fish allergens at a glance: Variable allergenicity of parvalbumins, the major fish allergens. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Mittag, D.; Vieths, S.; Vogel, L.; Becker, W.M.; Rihs, H.P.; Helbling, A.; Wüthrich, B.; Ballmer-Weber, B.K. Soybean allergy in patients allergic to birch pollen: Clinical investigation and molecular characterization of allergens. J. Allergy Clin. Immunol. 2004, 113, 148–154. [Google Scholar] [CrossRef]
- Cianferoni, A. Wheat allergy: Diagnosis and management. J. Asthma Allergy 2016, 9, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Caffarelli, C.; Baldi, F.; Bendandi, B.; Calzone, L.; Marani, M.; Pasquinelli, P. Cow’s milk protein allergy in children: A practical guide. Ital. J. Pediatr. 2010, 36. [Google Scholar] [CrossRef] [Green Version]
- Thompson-Chagoyan, O.C.; Vieites, J.M.; Maldonado, J.; Edwards, C.; Gil, A. Changes in faecal microbiota of infants with cow’s milk protein allergy—A Spanish prospective case-control 6-month follow-up study. Pediatr. Allergy Immunol. 2010, 21, e394–e400. [Google Scholar] [CrossRef]
- Hol, J.; van Leer, E.H.G.; Elink Schuurman, B.E.E.; de Ruiter, L.F.; Samsom, J.N.; Hop, W.; Neijens, H.J.; de Jongste, J.C.; Nieuwenhuis, E.E.S. The acquisition of tolerance toward cow’s milk through probiotic supplementation: A randomized, controlled trial. J. Allergy Clin. Immunol. 2008, 121, 1448–1454. [Google Scholar] [CrossRef]
- Martin, C.R.; Ling, P.R.; Blackburn, G.L. Review of infant feeding: Key features of breast milk and infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Ricci, G. Cow’s Milk Allergy: Management and Prevention. Int. J. Food Nutr. Sci. 2015, 2, 1–6. [Google Scholar] [CrossRef]
- Liao, S.-L.; Lai, S.-H.; Yeh, K.-W.; Huang, Y.-L.; Yao, T.-C.; Tsai, M.-H.; Hua, M.-C.; Huang, J.-L. Exclusive breastfeeding is associated with reduced cow’s milk sensitization in early childhood. Pediatr. Allergy Immunol. 2014, 25, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Caubet, J.C.; Lin, J.; Ahrens, B.; Gimenez, G.; Bardina, L.; Niggemann, B.; Sampson, H.A.; Beyer, K. Natural tolerance development in cow’s milk allergic children: IgE and IgG4 epitope binding. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Coates, M.E.; O’Donoghue, P.N. Milk Allergy in Infant Germ-free Rabbits. Nature 1967, 213, 307–308. [Google Scholar] [CrossRef]
- Rodriguez, B.; Prioult, G.; Bibiloni, R.; Nicolis, I.; Mercenier, A.; Butel, M.-J.; Waligora-Dupriet, A.-J. Germ-free status and altered caecal subdominant microbiota are associated with a high susceptibility to cow’s milk allergy in mice. FEMS Microbiol. Ecol. 2011, 76, 133–144. [Google Scholar] [CrossRef]
- Rodriguez, B.; Prioult, G.; Hacini-Rachinel, F.; Moine, D.; Bruttin, A.; Ngom-Bru, C.; Labellie, C.; Nicolis, I.; Berger, B.; Mercenier, A.; et al. Infant gut microbiota is protective against cow’s milk allergy in mice despite immature ileal T-cell response. FEMS Microbiol. Ecol. 2012, 79, 192–202. [Google Scholar] [CrossRef]
- Morin, S.; Bernard, H.; Przybylski-Nicaise, L.; Corthier, G.; Rabot, S.; Wal, J.-M.; Hazebrouck, S. Allergenic and immunogenic potential of cow’s milk β-lactoglobulin and caseins evidenced without adjuvant in germ-free mice. Mol. Nutr. Food Res. 2011, 55, 1700–1707. [Google Scholar] [CrossRef]
- Metsälä, J.; Lundqvist, A.; Virta, L.J.; Kaila, M.; Gissler, M.; Virtanen, S.M. Mother’s and offspring’s use of antibiotics and infant allergy to cow’s milk. Epidemiology 2013, 24, 303–309. [Google Scholar] [CrossRef]
- Bunyavanich, S.; Shen, N.; Grishin, A.; Wood, R.; Burks, W.; Dawson, P.; Jones, S.M.; Leung, D.Y.M.; Sampson, H.; Sicherer, S.; et al. Early-life gut microbiome composition and milk allergy resolution. J. Allergy Clin. Immunol. 2016, 138, 1122–1130. [Google Scholar] [CrossRef] [Green Version]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; Van Der Veeken, J.; Deroos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic T reg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canani, R.B.; Sangwan, N.; Stefka, A.T.; Nocerino, R.; Paparo, L.; Aitoro, R.; Calignano, A.; Khan, A.A.; Gilbert, J.A.; Nagler, C.R. Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants. ISME J. 2016, 10, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Choi Hong, S.M.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Peng, L.; Itzkowitz, S.; Holzman, I.R.; Babyatsky, M.W. Short-chain fatty acid induces intestinal mucosal injury in newborn rats and down-regulates intestinal trefoil factor gene expression in vivo and in vitro. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Chagoyan, O.C.; Fallani, M.; Maldonado, J.; Vieites, J.M.; Khanna, S.; Edwards, C.; Doré, J.; Gil, A. Faecal microbiota and short-chain fatty acid levels in faeces from infants with cow’s milk protein allergy. Int. Arch. Allergy Immunol. 2011, 156, 325–332. [Google Scholar] [CrossRef]
- Wopereis, H.; Van Ampting, M.T.J.; Cetinyurek-Yavuz, A.; Slump, R.; Candy, D.C.A.; Butt, A.M.; Peroni, D.G.; Vandenplas, Y.; Fox, A.T.; Shah, N.; et al. A specific synbiotic-containing amino acid-based formula restores gut microbiota in non-IgE mediated cow’s milk allergic infants: A randomized controlled trial. Clin. Transl. Allergy 2019, 9. [Google Scholar] [CrossRef]
- Berni Canani, R.; Nocerino, R.; Terrin, G.; Coruzzo, A.; Cosenza, L.; Leone, L.; Troncone, R. Effect of Lactobacillus GG on tolerance acquisition in infants with cow’s milk allergy: A randomized trial. J. Allergy Clin. Immunol. 2012, 129. [Google Scholar] [CrossRef]
- Villena, J.; Chiba, E.; Tomosada, Y.; Salva, S.; Marranzino, G.; Kitazawa, H.; Alvarez, S. Orally administered Lactobacillus rhamnosus modulates the respiratory immune response triggered by the viral pathogen-associated molecular pattern poly(I:C). BMC Immunol. 2012, 13, 53. [Google Scholar] [CrossRef] [Green Version]
- Salva, S.; Nuñez, M.; Villena, J.; Ramón, A.; Font, G.; Alvarez, S. Development of a fermented goats’ milk containing Lactobacillus rhamnosus: In vivo study of health benefits. J. Sci. Food Agric. 2011, 91, 2355–2362. [Google Scholar] [CrossRef]
- Cheng, R.Y.; Yao, J.R.; Wan, Q.; Guo, J.W.; Pu, F.F.; Shi, L.; Hu, W.; Yang, Y.H.; Li, L.; Li, M.; et al. Oral administration of Bifidobacterium bifidum TMC3115 to neonatal mice may alleviate IgE-mediated allergic risk in adulthood. Benef. Microb. 2018, 9, 815–828. [Google Scholar] [CrossRef]
- Takahashi, N.; Kitazawa, H.; Iwabuchi, N.; Xiao, J.Z.; Miyaji, K.; Iwatsuki, K.; Saito, T. Immunostimulatory oligodeoxynucleotide from Bifidobacterium longum suppresses Th2 immune responses in a murine model. Clin. Exp. Immunol. 2006, 145, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Schouten, B.; van Esch, B.C.A.M.; Hofman, G.A.; van Doorn, S.A.C.M.; Knol, J.; Nauta, A.J.; Garssen, J.; Willemsen, L.E.M.; Knippels, L.M.J. Cow Milk Allergy Symptoms Are Reduced in Mice Fed Dietary Synbiotics during Oral Sensitization with Whey. J. Nutr. 2009, 139, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Yang, Z.Y.; Dai, W.K.; Huang, J.Q.; Li, Y.H.; Juan, Z.; Qiu, C.Z.; Wei, C.; Zhou, Q.; Sun, X.; et al. Protective effect of Bifidobacterium infantis CGMCC313-2 on ovalbumin-induced airway asthma and β-lactoglobulininduced intestinal food allergy mouse models. World J. Gastroenterol. 2017, 23, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Sozańska, B. Raw Cow’s Milk and Its Protective Effect on Allergies and Asthma. Nutrients 2019, 11, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quigley, L.; O’sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. Molecular approaches to analysing the microbial composition of raw milk and raw milk cheese. Int. J. Food Microb. 2011. [Google Scholar] [CrossRef] [PubMed]
- Gran, H.M.; Wetlesen, A.; Mutukumira, A.N.; Rukure, G.; Narvhus, J.A. Occurrence of pathogenic bacteria in raw milk, cultured pasteurised milk and naturally soured milk produced at small-scale dairies in Zimbabwe. Food Control 2003, 14, 539–544. [Google Scholar] [CrossRef]
- Lemon-Mulé, H.; Sampson, H.A.; Sicherer, S.H.; Shreffler, W.G.; Noone, S.; Nowak-Wegrzyn, A. Immunologic changes in children with egg allergy ingesting extensively heated egg. J. Allergy Clin. Immunol. 2008, 122, 977–983.e1. [Google Scholar] [CrossRef]
- Leonard, S.A.; Sampson, H.A.; Sicherer, S.H.; Noone, S.; Moshier, E.L.; Godbold, J.; Nowak-Wegrzyn, A. Dietary baked egg accelerates resolution of egg allergy in children. J. Allergy Clin. Immunol. 2012, 130, 473–480.e1. [Google Scholar] [CrossRef] [Green Version]
- Fazlollahi, M.; Chun, Y.; Grishin, A.; Wood, R.A.; Burks, A.W.; Dawson, P.; Jones, S.M.; Leung, D.Y.M.; Sampson, H.A.; Sicherer, S.H.; et al. Early-life gut microbiome and egg allergy. Allergy 2018, 73, 1515–1524. [Google Scholar] [CrossRef]
- Yu, M.; Cui, F.X.; Jia, H.M.; Zhou, C.; Yang, Y.; Zhang, H.W.; Ding, G.; Zou, Z.M. Aberrant purine metabolism in allergic asthma revealed by plasma metabolomics. J. Pharm. Biomed. Anal. 2016, 120, 181–189. [Google Scholar] [CrossRef]
- Kim, H.; Kwack, K.; Kim, D.-Y.; Ji, G.E. Oral probiotic bacterial administration suppressed allergic responses in an ovalbumin-induced allergy mouse model. FEMS Immunol. Med. Microbiol. 2005, 45, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jeun, E.J.; Hong, C.P.; Kim, S.H.; Jang, M.S.; Lee, E.J.; Moon, S.J.; Yun, C.H.; Im, S.H.; Jeong, S.G.; et al. Extracellular vesicle-derived protein from Bifidobacterium longum alleviates food allergy through mast cell suppression. J. Allergy Clin. Immunol. 2016, 137, 507–516.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Gadir, A.; Stephen-Victor, E.; Gerber, G.K.; Noval Rivas, M.; Wang, S.; Harb, H.; Wang, L.; Li, N.; Crestani, E.; Spielman, S.; et al. Microbiota therapy acts via a regulatory T cell MyD88/RORγt pathway to suppress food allergy. Nat. Med. 2019, 25, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.; Bunyavanich, S. Microbial Adjuncts for Food Allergen Immunotherapy. Curr. Allergy Asthma Rep. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Iweala, O.I.; Choudhary, S.K.; Commins, S.P. Food Allergy. Curr. Gastroenterol. Rep. 2018, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, L. Quality of life in the setting of anaphylaxis and food allergy. Allergo J. Int. 2014, 23, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherkaoui, S.; Ben-Shoshan, M.; Alizadehfar, R.; Asai, Y.; Chan, E.; Cheuk, S.; Shand, G.; St-Pierre, Y.; Harada, L.; Allen, M.; et al. Accidental exposures to peanut in a large cohort of Canadian children with peanut allergy. Clin. Transl. Allergy 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Warren, C.; Blumenstock, J.; Kotowska, J.; Mittal, K.; Smith, B. The prevalence of childhood food allergy in the United States: An update. Ann. Allergy Asthma Immunol. 2017, 119, S11. [Google Scholar] [CrossRef]
- Chu, D.K.; Wood, R.A.; French, S.; Fiocchi, A.; Jordana, M.; Waserman, S.; Brożek, J.L.; Schünemann, H.J. Oral immunotherapy for peanut allergy (PACE): A systematic review and meta-analysis of efficacy and safety. Lancet 2019, 393, 2222–2232. [Google Scholar] [CrossRef]
- PALFORZIA | FDA. Available online: https://www.fda.gov/vaccines-blood-biologics/allergenics/palforzia (accessed on 11 June 2020).
- Tang, M.L.K.; Ponsonby, A.L.; Orsini, F.; Tey, D.; Robinson, M.; Su, E.L.; Licciardi, P.; Burks, W.; Donath, S. Administration of a probiotic with peanut oral immunotherapy: A randomized trial. J. Allergy Clin. Immunol. 2015, 135, 737–744.e8. [Google Scholar] [CrossRef]
- Hsiao, K.C.; Ponsonby, A.L.; Axelrad, C.; Pitkin, S.; Tang, M.L.K.; Burks, W.; Donath, S.; Orsini, F.; Tey, D.; Robinson, M.; et al. Long-term clinical and immunological effects of probiotic and peanut oral immunotherapy after treatment cessation: 4-year follow-up of a randomised, double-blind, placebo-controlled trial. Lancet Child Adolesc. Heal. 2017, 1, 97–105. [Google Scholar] [CrossRef]
- Evaluating the Safety and Efficacy of Oral Encapsulated Fecal Microbiota Transplant in Peanut Allergic Patients-Full Text View-ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02960074 (accessed on 7 December 2019).
- NCT03936998 VE416 for Treatment of Food Allergy. Available online: https://clinicaltrials.gov/ct2/show/NCT03936998 (accessed on 16 January 2020).
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K.; Mazmanian, S.K.; Tjota, M.Y.; Seo, G.Y.; Cao, S.; Theriault, B.R.; et al. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berin, M.C. Mucosal antibodies in the regulation of tolerance and allergy to foods. Semin. Immunopathol. 2012, 34, 633–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, J.; Chalcraft, K.; Mandur, T.S.; Jimenez-Saiz, R.; Walker, T.D.; Goncharova, S.; Gordon, M.E.; Naji, L.; Flader, K.; Larché, M.; et al. Comprehensive metabolomics identifies the alarmin uric acid as a critical signal for the induction of peanut allergy. Allergy 2015, 70, 495–505. [Google Scholar] [CrossRef]
- Bamberger, C.; Rossmeier, A.; Lechner, K.; Wu, L.; Waldmann, E.; Fischer, S.; Stark, R.; Altenhofer, J.; Henze, K.; Parhofer, K. A Walnut-Enriched Diet Affects Gut Microbiome in Healthy Caucasian Subjects: A Randomized, Controlled Trial. Nutrients 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, C.E.; Nicklas, T.A.; Fulgoni, V.L. Tree nut consumption is associated with better nutrient adequacy and diet quality in adults: National health and nutrition examination survey 2005–2010. Nutrients 2015, 7, 595–607. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.; Goedert, J.J.; Pu, A.; Yu, G.; Shi, J. Allergy associations with the adult fecal microbiota: Analysis of the American Gut Project. EBioMedicine 2016, 3, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, T.; Sicherer, S. Current perspectives on tree nut allergy: A review. J. Asthma Allergy 2018, 11, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Elizur, A.; Appel, M.Y.; Nachshon, L.; Levy, M.B.; Epstein-Rigbi, N.; Pontoppidan, B.; Lidholm, J.; Goldberg, M.R. Walnut oral immunotherapy for desensitisation of walnut and additional tree nut allergies (Nut CRACKER): A single-centre, prospective cohort study. Lancet Child Adolesc. Heal. 2019, 3, 312–321. [Google Scholar] [CrossRef]
- Enrique, E.; Pineda, F.; Malek, T.; Bartra, J.; Basagaña, M.; Tella, R.; Castelló, J.V.; Alonso, R.; De Mateo, J.A.; Cerdá-Trias, T.; et al. Sublingual immunotherapy for hazelnut food allergy: A randomized, double-blind, placebo-controlled study with a standardized hazelnut extract. J. Allergy Clin. Immunol. 2005, 116, 1073–1079. [Google Scholar] [CrossRef]
- Wong, L.; Huang, C.H.; Lee, B.W. Shellfish and house dust mite allergies: Is the link tropomyosin? Allergy Asthma Immunol. Res. 2016, 8, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.S.; Warren, C.M.; Smith, B.M.; Jiang, J.; Blumenstock, J.A.; Davis, M.M.; Schleimer, R.P.; Nadeau, K.C. Prevalence and Severity of Food Allergies Among US Adults. JAMA Netw. Open 2019, 2, e185630. [Google Scholar] [CrossRef] [PubMed]
- Wild, L.G.; Lehrer, S.B. Fish and shellfish allergy. Curr. Allergy Asthma Rep. 2005, 5, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Song, J.; Wang, C.; Fu, S.; Wang, Y. Bifidobacterium infantis potentially alleviates shrimp tropomyosin-induced allergy by tolerogenic dendritic cell-dependent induction of regulatory T cells and alterations in gut microbiota. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Liu, Q.; Jing, W.; Wang, W. Bifidobacterium lactis Ameliorates the Risk of Food Allergy in Chinese Children by Affecting Relative Percentage of Treg and Th17 Cells. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Fu, S.; Wang, C.; Xie, M.; Wang, Y. Yogurt-sourced probiotic bacteria alleviate shrimp tropomyosin-induced allergic mucosal disorders, potentially through microbiota and metabolism modifications. Allergol. Int. 2019, 68, 506–514. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Muller, A.; Young, V.B.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2017, 167, 1339–1353. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Bastida, J.A.; Piskuła, M.; Zieliński, H. Recent advances in development of gluten-free buckwheat products. Trends Food Sci. Technol. 2015, 44, 58–65. [Google Scholar] [CrossRef]
- Savage, J.H.; Kaeding, A.J.; Matsui, E.C.; Wood, R.A. The natural history of soy allergy. J. Allergy Clin. Immunol. 2010, 125, 683–686. [Google Scholar] [CrossRef]
- Anandan, C.; Nurmatov, U.; Sheikh, A. Omega 3 and 6 oils for primary prevention of allergic disease: Systematic review and meta-analysis. Allergy 2009, 64, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Untersmayr, E.; Schöll, I.; Swoboda, I.; Beil, W.J.; Förster-Waldl, E.; Walter, F.; Riemer, A.; Kraml, G.; Kinaciyan, T.; Spitzauer, S.; et al. Antacid medication inhibits digestion of dietary proteins and causes food allergy: A fish allergy model in Balb/c mice. J. Allergy Clin. Immunol. 2003, 112, 616–623. [Google Scholar] [CrossRef]
- Hougee, S.; Vriesema, A.J.M.; Wijering, S.C.; Knippels, L.M.J.; Folkerts, G.; Nijkamp, F.P.; Knol, J.; Garssen, J. Oral Treatment with Probiotics Reduces Allergic Symptoms in Ovalbumin-Sensitized Mice: A Bacterial Strain Comparative Study. Int. Arch. Allergy Immunol. 2010, 151, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Matricardi, P.M.; Rosmini, F.; Panetta, V.; Ferrigno, L.; Bonini, S. Hay fever and asthma in relation to markers of infection in the United States. J. Allergy Clin. Immunol. 2002, 110, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Leach, D.; Hunter, D.A.; Sanfelippo, D.; Buell, E.J.; Zemple, S.J.; Grayson, M.H. Characterization of intestinal dendritic cells in murine norovirus infection. Open Immunol. J. 2011, 4, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Sullender, M.E.; Baldridge, M.T. Norovirus interactions with the commensal microbiota. PLoS Pathogens 2018. [Google Scholar] [CrossRef]
- Pearson, J.A.; Tai, N.; Ekanayake-Alper, D.K.; Peng, J.; Hu, Y.; Hager, K.; Compton, S.; Wong, F.S.; Smith, P.C.; Wen, L. Norovirus Changes Susceptibility to Type 1 Diabetes by Altering Intestinal Microbiota and Immune Cell Functions. Front. Immunol. 2019, 10, 2654. [Google Scholar] [CrossRef]
- Holm, S.; Andersson, Y.; Gothefors, L.; Lindberg, T. Increased protein absorption after acute gastroenteritis in children. Acta Pædiatr. 1992, 81, 585–588. [Google Scholar] [CrossRef]
- Minot, S.; Sinha, R.; Chen, J.; Li, H.; Keilbaugh, S.A.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. The human gut virome: Inter-individual variation and dynamic response to diet. Genom. Res. 2011, 21, 1616–1625. [Google Scholar] [CrossRef] [Green Version]
- Langella, P.; Guarner, F.; Martín, R. Editorial: Next-Generation Probiotics: From Commensal Bacteria to Novel Drugs and Food Supplements. Front. Microbiol. 2019, 10, 1973. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Cow milk | Casein, whey, bovine serum albumin, immunoglobulin G heavy chain and a-lactalbumin [19] |
| Hen’s egg | Whites: Ovomucoid, ovalbumin, ovotransferring, and lysosome [20] Yolk: Livetin, vitellenin and apoprotein B [20] |
| Peanut | Cupin(Ara h 1, 3), prolamin(Ara h 2, 6, 7, 9), profilin(Ara h 5), bet v-1-related proteins(Ara h 8), oleosin(Ara h 10,11) and defensin(Ara h 12, 13) [21] |
| Tree nut | Vicilin, 2S albumin, nsLTP, legumin, bet v 1-like, oleosin, 60 s acidic ribosomal prot. P2, manganese superoxide dismutase and profiling [22] |
| Shellfish | Tropomyosin, arginine kinase, myosin light chain, and sarcoplasmic calcium-binding protein [23] |
| Finfish | Parvalbumins, enolase, aldolase, tropomyosin, vitellogenin [24] |
| Soy | Hull proteins, kunitz trypsin inhibitor, glycinin, α-subunit of β-con-glycinin, and 50-kd protein with homology to chlorophyll A-B binding protein and starvation-associated message [25] |
| Wheat | α-Amylase/trypsin inhibitor, album, globulins, serine proteinase inhibitor, lipid protein transfer, thioredoxin, peroxidase, gliadin, thiol reductase, and thaumatin-like protein [26] |
| Promote Risk | Reduce Risk | Therapies and Supplements | |
|---|---|---|---|
| Cow milk | d General antibiotic use before and during pregnancy h [38] | a Increase in diversity h [39] | a,bBifidobacterium infantis CGMCC313-2, 5 × 1010 CFU/mL for 6 days m [53] |
| d Short-chain fatty acid m [3] | a Breastfeeding h [32] | cLactobacillus rhamnosus GG, 4.5 × 107–8.5 × 107 CFU/g of formula for 6 months h [42] | |
| eClostridium coccoidesh [45] | a,e Firmicutes h,m [39,42,43] | cLactobacillus rhamnosus GG, 1.4 × 107 CFU/100 mL of formula for 6–12 months [47] | |
| eAtopobiumh [45] | a,e Clostridia h,m [39,42,43] | ||
| eLachnospiraceaeh [42] | eOscillospirah [42] | aBifidobacterium breve M-16V, 2 × 109 colony forming unit (CFU)/g for 7 weeks m [52] | |
| eRuminococcaceaeh [42] | eRoseburiah [42] | ||
| eLactobacillih [4] | eBlautiah [42] | ||
| aAnaerostipes caccaeh,m [43] | a,eBacteroidesh,m [36] | ||
| a,eBifidobacteriumh,m [28,36] | eEnterobacteriah [4] | ||
| Hen’s egg | e Increase in diversity h [59] | eLeuconostocaceaeh [59] | aBifidobacterium longum, 5 × 109 CFU daily for 36 days m [62] |
| eLachnospiraceaeh [6,59] | a Clostridiales consortium, 5 × 107 CFU twice weekly for five weeks m [63] | ||
| eStreptococcaceaeh [59] | aSubdoligranulum variabile, 2.4 × 106 CFU twice weekly for five weeks m [63] | ||
| eLactobacillaceaem [6] | a Bacteroidales consortium 5 × 107 CFU twice weekly for five weeks m [63] | ||
| eRikenellaceaem [6] | aBifidobacterium bifidum, 0.2% lyophilized for 8 weeks m [61] | ||
| ePorphyromonadaceaem [6] | aLactobacillus casei, lyophilized for 8 weeks m [61] | ||
| dProteobacteria consortiumm [63] | aEscherichia coli lyophilized for 8 weeks m [61] | ||
| eRuminococcaceaeh [59] | a Clostridia via monocolonization m [64] | ||
| Peanut | cLactobacillus rhamnosus and peanut oral immunotherapy, 2 × 1010 CFU once daily with peanut protein for 18 months h [71] | ||
| cLactobacillus rhamnosus GG and peanut oral immunotherapy, 2 × 1010 CFU of L rhamnosus CGMCC 1.3724 and 2 g of peanut protein once daily for 18 months h [72] | |||
| a High-fiber diet and vitamin A for 2 weeks m [12] | |||
| a Direct feeding of acetate and butyrate for 3 weeks m [12] | |||
| a Clostridia via 2 oral gavages, once a week m [75] | |||
| Tree nuts | e Bacteroidales h [80] | e Clostridiales h [80] | |
| eBacteroides fragilish [80] | ePrevotellah [80] | ||
| e Bacteroidales h [80] | eRuminococcaceaeh [80] | ||
| Shellfish | eRalstoniam [87] | eDoream [87] | a,bBifidobacterium infantis, 107 CFU/mL for 20 days m [87] |
| bBifidobacterium lactis, 10 mL (1 × 106/mL) daily for 3 months h [88] | |||
| aBifidobacterium longum, 2 × 1010 CFU for 22 days m [89] | |||
| aBacillus coagulans 2 × 1010 CFU for 22 days m [89] | |||
| Finfish | |||
| Wheat | No data present | ||
| Soy | |||
| Proposed Mechanisms | Allergens Involved |
|---|---|
| Increase in diversity strengthens and stablishes immune system. h [39] | Cow milk |
| Exclusive breastfeeding prevents the introduction of cow milk required for sensitization. h [32] | Cow milk |
| Firmicutes, those in the Clostridia order and those which increased butyrate production, which regulate colonic regulatory T cells. mh [12,39,42,43,63,64] | Cow milk, Hen’s egg |
| Those in the Clostridia order regulate innate lymphoid cell function to alter epithelial permeability and reduce allergen uptake into the systemic circulation. m [75] | Cow milk |
| Pyruvate metabolism from those in the Clostridia order and Leuconostocaceae family depletes uric acid, which inhibits the activity of xanthine oxidase. mh [43,59,75] | Cow milk, Hen’s egg |
| Bifidobacterium and Bacteroides invoke foxp3 gene activation, which is responsible for the development and function of regulatory T cells. mh [28,36,52,53] | Cow milk |
| B. infantis increased the abundance of butyrate-producing bacteria, which in turn suppress the inflammatory responses triggered by Th2 cytokines. m [53,96] | Shellfish |
| Lactobacillus rhamnosus GG promotes growth of short-chain fatty acid-producing bacteria. h [42] | Cow milk, Peanut |
| Bifidobacterium longum induces apoptosis to bonded mast cells. m [62] | Hen’s egg |
| Antibiotics decrease level of beneficial bacteria. h [38] | Cow milk |
| Firmicutes and those in the Clostridia order increased butyrate production, which in turn increases the permeability of the intestinal mucosa. mh [28,44] | Cow milk |
| Bifidobacterium longum and Bacillus coagulans regulate gut arginine metabolism pathways and the metabolites of aspartate and arginine may be critical for prevention of food allergy. m [89] | Shellfish |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suther, C.; Moore, M.D.; Beigelman, A.; Zhou, Y. The Gut Microbiome and the Big Eight. Nutrients 2020, 12, 3728. https://doi.org/10.3390/nu12123728
Suther C, Moore MD, Beigelman A, Zhou Y. The Gut Microbiome and the Big Eight. Nutrients. 2020; 12(12):3728. https://doi.org/10.3390/nu12123728
Chicago/Turabian StyleSuther, Cassandra, Matthew D. Moore, Avraham Beigelman, and Yanjiao Zhou. 2020. "The Gut Microbiome and the Big Eight" Nutrients 12, no. 12: 3728. https://doi.org/10.3390/nu12123728