Relationship Between the Gastrointestinal Side Effects of an Anti-Hypertensive Medication and Changes in the Serum Lipid Metabolome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Rats
2.2. Serum Collection and Serum Lipid Metabolite Analysis
2.3. Processing and Analysis of Mass Spectrometry Data
2.4. The Criteria for Metabolite Selection
3. Results and Discussion
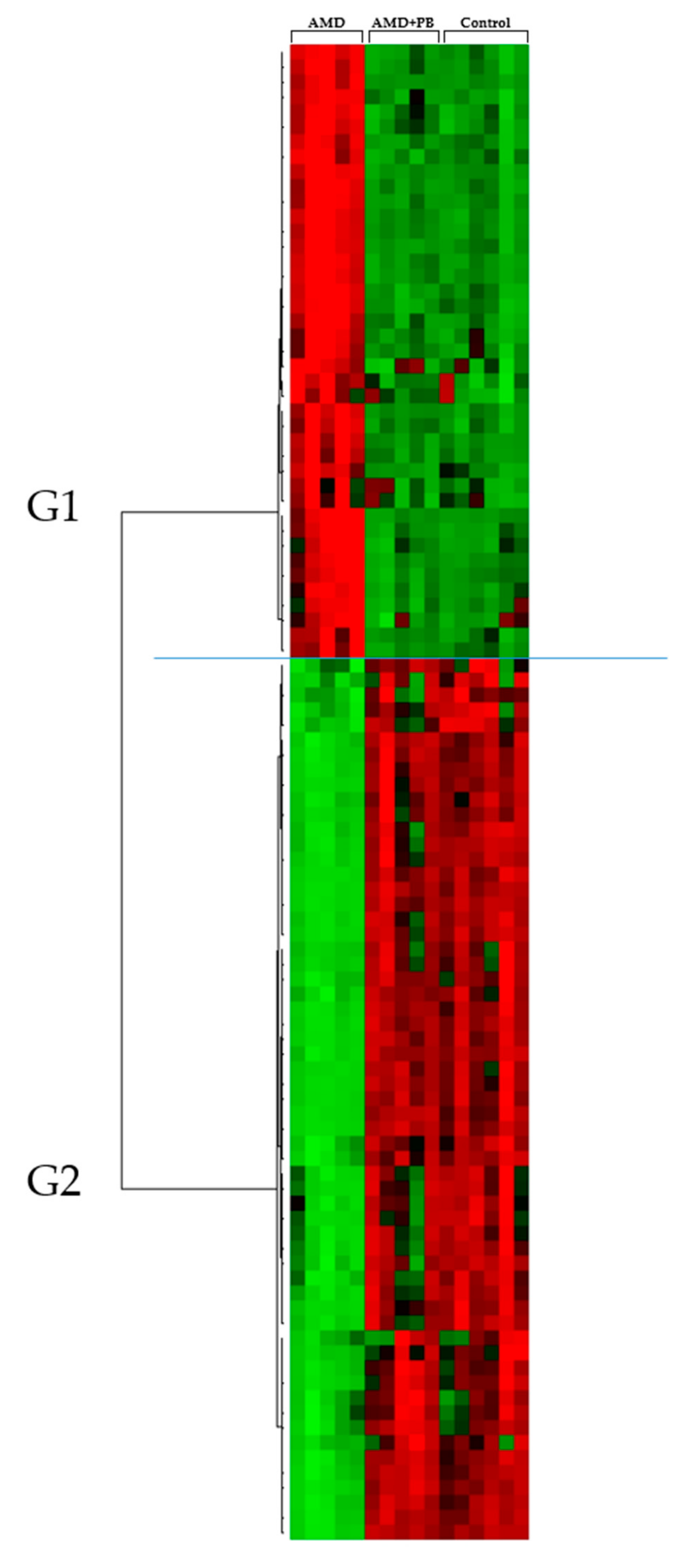
3.1. Statistical Analysis of the Serum Metabolome
3.2. Screening and Identification of Candidate Markers for Lipid Metabolites
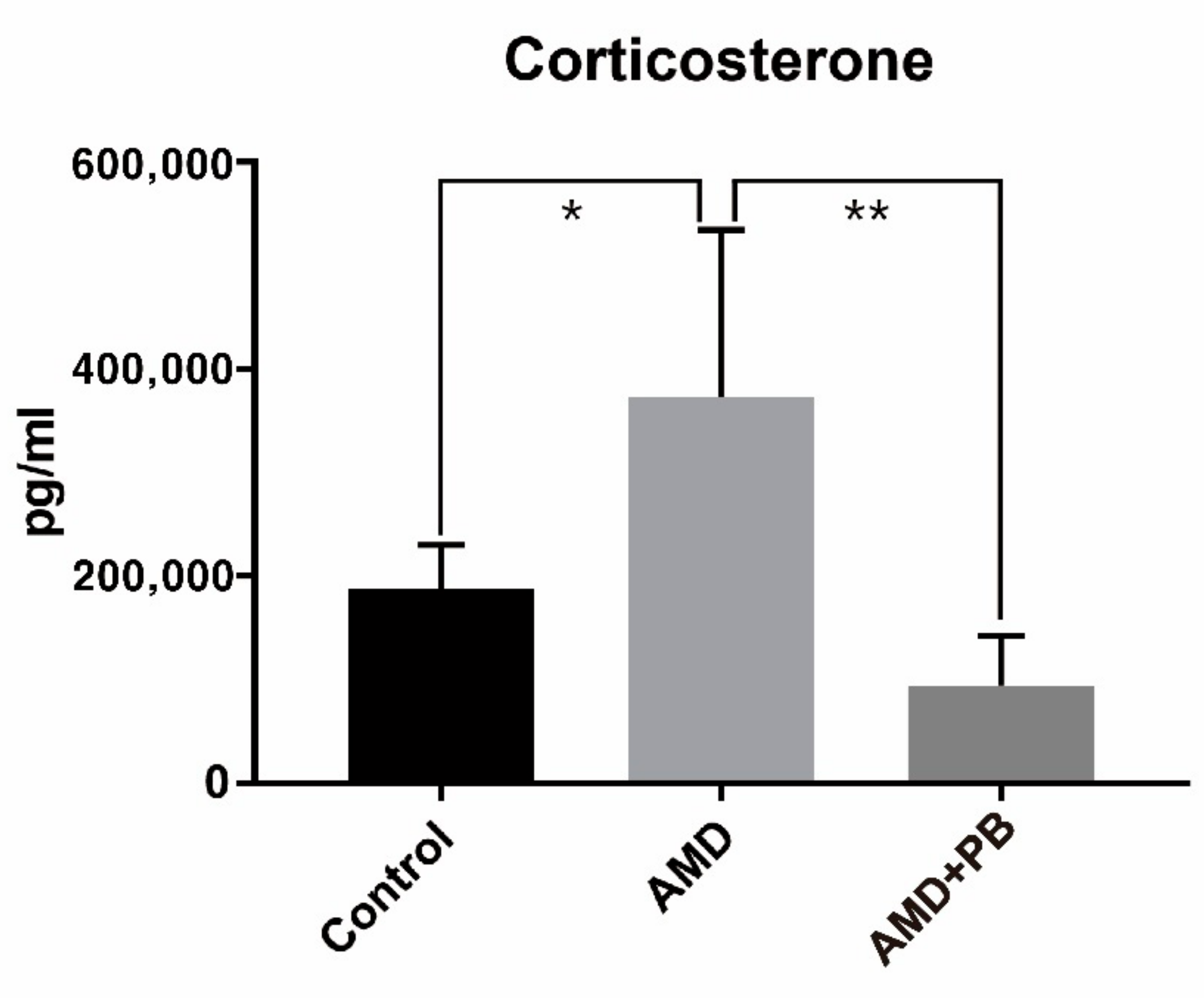
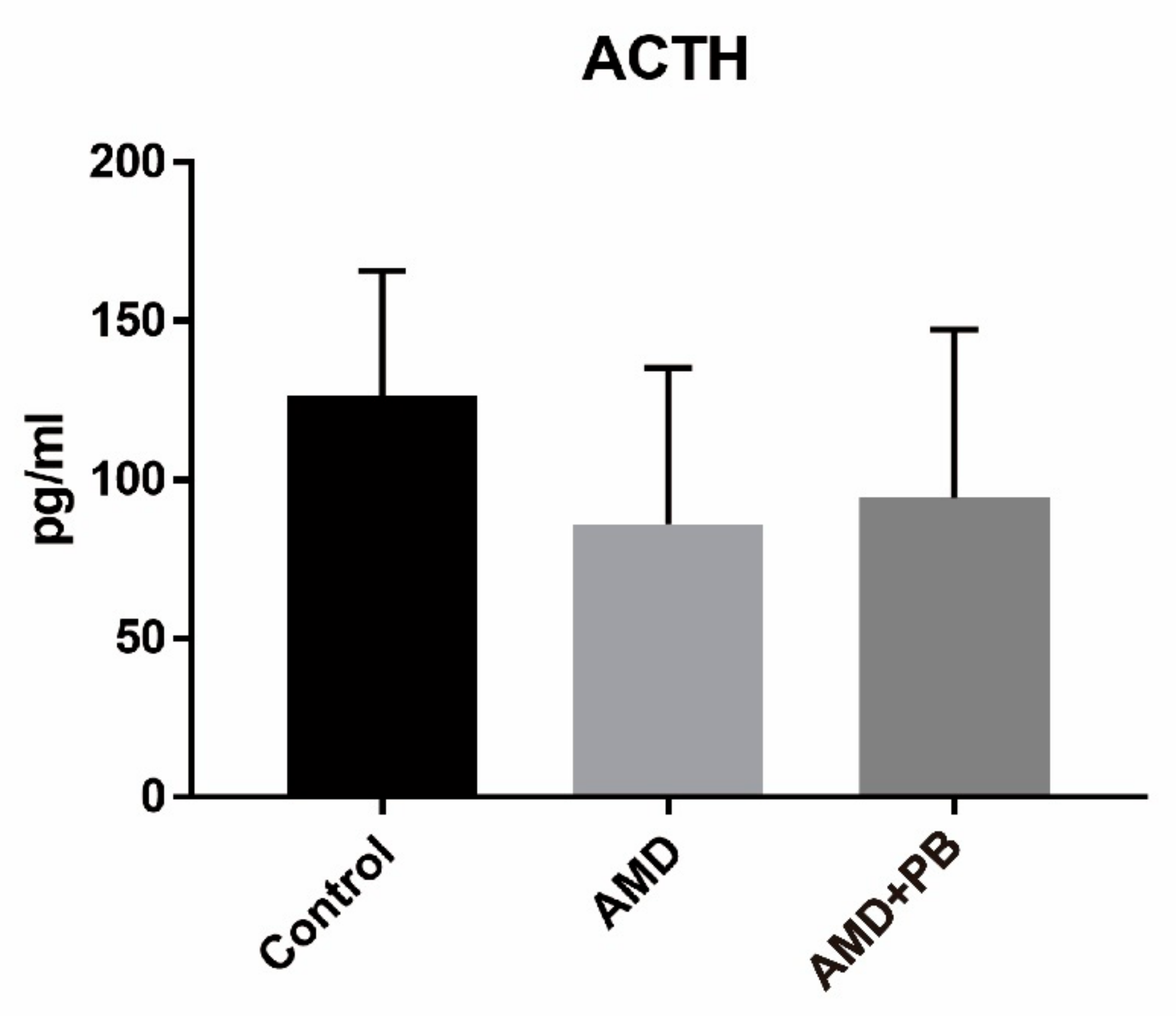
3.3. Identification and Comparison of Corticosterone and Adrenocorticotropic Hormone (ACTH)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dorans, K.S.; Mills, K.T.; Liu, Y.; He, J. Trends in Prevalence and Control of Hypertension According to the 2017 American College of Cardiology/American Heart Association (ACC/AHA) Guideline. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the Gut Microbiota in Disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Chassard, C.; Dapoigny, M.; Scott, K.P.; Crouzet, L.; Del’homme, C.; Marquet, P.; Martin, J.C.; Pickering, G.; Ardid, D.; Eschalier, A.; et al. Functional Dysbiosis within the Gut Microbiota of Patients with Constipated-Irritable Bowel Syndrome. Aliment. Pharm. 2012, 35, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Duboc, H.; Rainteau, D.; Rajca, S.; Humbert, L.; Farabos, D.; Maubert, M.; Grondin, V.; Jouet, P.; Bouhassira, D.; Seksik, P.; et al. Increase in Fecal Primary Bile Acids and Dysbiosis in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2012, 24, 513–e247. [Google Scholar] [CrossRef] [PubMed]
- Belizário, J.E.; Faintuch, J. Microbiome and Gut Dysbiosis. Exp. Suppl. 2018, 109, 459–476. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H. Problems with the Concept of Gut Microbiota Dysbiosis. Microb. Biotechnol. 2019, 1751–7915, 13479. [Google Scholar] [CrossRef] [Green Version]
- Le Bastard, Q.; Al-Ghalith, G.A.; Grégoire, M.; Chapelet, G.; Javaudin, F.; Dailly, E.; Batard, E.; Knights, D.; Montassier, E. Systematic Review: Human Gut Dysbiosis Induced by Non-Antibiotic Prescription Medications. Aliment. Pharm. 2018, 47, 332–345. [Google Scholar] [CrossRef] [Green Version]
- Althani, A.A.; Marei, H.E.; Hamdi, W.S.; Nasrallah, G.K.; El Zowalaty, M.E.; Al Khodor, S.; Al-Asmakh, M.; Abdel-Aziz, H.; Cenciarelli, C. Human Microbiome and Its Association with Health and Diseases. J. Cell. Physiol. 2016, 231, 1688–1694. [Google Scholar] [CrossRef]
- Guinane, C.M.; Cotter, P.D. Role of the Gut Microbiota in Health and Chronic Gastrointestinal Disease: Understanding a Hidden Metabolic Organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. Gut Microbiome Dysbiosis and Immunometabolism: New Frontiers for Treatment of Metabolic Diseases. Mediat. Inflamm. 2018, 2018, 2037838. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional Interactions between the Gut Microbiota and Host Metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Liang, S.; Wu, X.; Jin, F. Gut-Brain Psychology: Rethinking Psychology from the Microbiota–Gut–Brain Axis. Front. Integr. Neurosci. 2018, 12, 33. [Google Scholar] [CrossRef]
- Erny, D.; Hrabě de Angelis, A.L.; Prinz, M. Communicating Systems in the Body: How Microbiota and Microglia Cooperate. Immunology 2017, 150, 7–15. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Song, S.-C.; An, Y.-M.; Shin, J.-H.; Chung, M.-J.; Seo, J.-G.; Kim, E. Beneficial Effects of a Probiotic Blend on Gastrointestinal Side Effects Induced by Leflunomide and Amlodipine in a Rat Model. Benef. Microbes 2017, 8, 801–808. [Google Scholar] [CrossRef]
- Zicha, J.; Kunes, J.; Devynck, M.A. Abnormalities of Membrane Function and Lipid Metabolism in Hypertension: A Review. Am. J. Hypertens. 1999, 12, 315–331. [Google Scholar] [CrossRef]
- Sevilla, M.A.; Voces, F.; Carrón, R.; Guerrero, E.I.; Ardanaz, N.; San Román, L.; Arévalo, M.A.; Montero, M.J. Amlodipine Decreases Fibrosis and Cardiac Hypertrophy in Spontaneously Hypertensive Rats: Persistent Effects after Withdrawal. Life Sci. 2004, 75, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Eros, G.; Varga, G.; Vairadi, R.; Czobel, M.; Kaszaki, J.; Ghyczy, M.; Boros, M. Anti-Inflammatory Action of a Phosphatidylcholine, Phosphatidylethanolamine and N-Acylphosphatidylethanolamine-Enriched Diet in Carrageenan-Induced Pleurisy. Eur. Surg. Res. 2009, 42, 40–48. [Google Scholar] [CrossRef]
- Treede, I.; Braun, A.; Sparla, R.; Kühnel, M.; Giese, T.; Turner, J.R.; Anes, E.; Kulaksiz, H.; Füllekrug, J.; Stremmel, W.; et al. Anti-Inflammatory Effects of Phosphatidylcholine. J. Biol. Chem. 2007, 282, 27155–27164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oneta, C.M.; Mak, K.M.; Lieber, C.S. Dilinoleoylphosphatidylcholine Selectively Modulates Lipopolysaccharide-Induced Kupffer Cell Activation. J. Lab. Clin. Med. 1999, 134, 466–470. [Google Scholar] [CrossRef]
- Taniuchi, M.; Otani, H.; Kodama, N.; Tone, Y.; Sakagashira, M.; Yamada, Y.; Mune, M.; Yukawa, S. Lysophosphatidylcholine Up-Regulates IL-1 Beta-Induced INOS Expression in Rat Mesangial Cells. Kidney Int. Suppl. 1999, 71, S156–S158. [Google Scholar] [CrossRef] [Green Version]
- Han, M.S.; Park, S.Y.; Shinzawa, K.; Kim, S.; Chung, K.W.; Lee, J.-H.; Kwon, C.H.; Lee, K.-W.; Lee, J.-H.; Park, C.K.; et al. Lysophosphatidylcholine as a Death Effector in the Lipoapoptosis of Hepatocytes. J. Lipid Res. 2008, 49, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Xu, M.-J.; Gao, B. Hepatocytes: A Key Cell Type for Innate Immunity. Cellular and Molecular Immunology 2015, 13, 301. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Zhang, W.; Gu, X.; Chen, X.; Hong, L.; Laird, J.M.; Salomon, R.G. Lysophosphatidylcholine Is Generated by Spontaneous Deacylation of Oxidized Phospholipids. Chem. Res. Toxicol. 2011, 24, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Wi, S.J.; Seo, S.Y.; Cho, K.; Nam, M.H.; Park, K.Y. Lysophosphatidylcholine Enhances Susceptibility in Signaling Pathway against Pathogen Infection through Biphasic Production of Reactive Oxygen Species and Ethylene in Tobacco Plants. Phytochemistry 2014, 104, 48–59. [Google Scholar] [CrossRef]
- Riederer, M.; Lechleitner, M.; Hrzenjak, A.; Koefeler, H.; Desoye, G.; Heinemann, A.; Frank, S. Endothelial Lipase (EL) and EL-Generated Lysophosphatidylcholines Promote IL-8 Expression in Endothelial Cells. Atherosclerosis 2011, 214, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Feng, G.; Zhao, B.; Zhao, J.; Pi, Z.; Liu, S.; Song, F.; Liu, Z. A Non-Target Urinary and Serum Metabolomics Strategy Reveals Therapeutical Mechanism of Radix Astragali on Adjuvant-Induced Arthritis Rats. J. Chromatogr. B 2017, 1048, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-H.; Nam, M.H.; Lee, H.; Lee, J.-S.; Kim, H.; Chung, M.-J.; Seo, J.-G. Amelioration of Obesity-Related Characteristics by a Probiotic Formulation in a High-Fat Diet-Induced Obese Rat Model. Eur. J. Nutr. 2018, 57, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Yaligar, J.; Gopalan, V.; Kiat, O.W.; Sugii, S.; Shui, G.; Lam, B.D.; Henry, C.J.; Wenk, M.R.; Tai, E.S.; Velan, S.S. Evaluation of Dietary Effects on Hepatic Lipids in High Fat and Placebo Diet Fed Rats by In Vivo MRS and LC-MS Techniques. Plos ONE 2014, 9, e91436. [Google Scholar] [CrossRef] [PubMed]
- Abbott, M.J.; Tang, T.; Sul, H.S. The Role of Phospholipase A2-Derived Mediators in Obesity. Drug Discov. Today Dis. Mech. 2010, 7, e213–e218. [Google Scholar] [CrossRef] [Green Version]
- Fox, L.M.; Cox, D.G.; Lockridge, J.L.; Wang, X.; Chen, X.; Scharf, L.; Trott, D.L.; Ndonye, R.M.; Veerapen, N.; Besra, G.S.; et al. Recognition of Lyso-Phospholipids by Human Natural Killer T Lymphocytes. PLoS Biol. 2009, 7, e1000228. [Google Scholar] [CrossRef] [Green Version]
- Frasch, S.C.; Zemski-Berry, K.; Murphy, R.C.; Borregaard, N.; Henson, P.M.; Bratton, D.L. Lysophospholipids of Different Classes Mobilize Neutrophil Secretory Vesicles and Induce Redundant Signaling through G2A. J. Immunol. 2007, 178, 6540–6548. [Google Scholar] [CrossRef] [Green Version]
- AbouRjaili, G.; Shtaynberg, N.; Wetz, R.; Costantino, T.; Abela, G.S. Current Concepts in Triglyceride Metabolism, Pathophysiology, and Treatment. Metabolism 2010, 59, 1210–1220. [Google Scholar] [CrossRef]
- Jonkers, I.J.; Mohrschladt, M.F.; Westendorp, R.G.; van der Laarse, A.; Smelt, A.H. Severe Hypertriglyceridemia with Insulin Resistance Is Associated with Systemic Inflammation: Reversal with Bezafibrate Therapy in a Randomized Controlled Trial. Am. J. Med. 2002, 112, 275–280. [Google Scholar] [CrossRef]
- Wollam, J.; Antebi, A. Sterol Regulation of Metabolism, Homeostasis, and Development. Annu. Rev. Biochem. 2011, 80, 885–916. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Li, J.; Zhang, W. Hormonal Regulation of Cholesterol Homeostasis. In Cholesterol—Good, Bad and the Heart; Nagpal, M.L., Ed.; InTech: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, Inflammation and Innate Immunity. Nat. Rev. Immunol. 2015, 15, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Milot, M.R.; James, J.S.; Merali, Z.; Plamondon, H. A Refined Blood Collection Method for Quantifying Corticosterone. Lab. Anim. 2012, 41, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Vahl, T.P.; Ulrich-Lai, Y.M.; Ostrander, M.M.; Dolgas, C.M.; Elfers, E.E.; Seeley, R.J.; D’Alessio, D.A.; Herman, J.P. Comparative Analysis of ACTH and Corticosterone Sampling Methods in Rats. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E823–E828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djordjević, J.; Cvijić, G.; Davidović, V. Different Activation of ACTH and Corticosterone Release in Response to Various Stressors in Rats. Physiol. Res. 2003, 52, 67–72. [Google Scholar] [PubMed]
- Yoon, J.S.; Sohn, W.; Lee, O.Y.; Lee, S.P.; Lee, K.N.; Jun, D.W.; Lee, H.L.; Yoon, B.C.; Choi, H.S.; Chung, W.-S.; et al. Effect of Multispecies Probiotics on Irritable Bowel Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial: Probiotics in Irritable Bowel Syndrome. J. Gastroenterol. Hepatol. 2014, 29, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.J.; Lim, T.J.; Lim, Y.J.; Seo, J.G.; Chung, M.J. In Vivo Effects of s-Pantoprazole, Polaprenzinc, and Probiotic Blend on Chronic Small Intestinal Injury Induced by Indomethacin. Benef. Microbes 2016, 7, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.H.; Jacobson, L.; Pacák, K.; Widmaier, E.P.; Goldstein, D.S.; Majzoub, J.A. Impaired Basal and Restraint-Induced Epinephrine Secretion in Corticotropin-Releasing Hormone-Deficient Mice. Endocrinology 2000, 141, 1142–1150. [Google Scholar] [CrossRef]
- Chio, S.L.; Sin, Y.M. Changes in Corticosterone Levels under Different Degrees of Acute Inflammation in Mice. Agents Actions 1992, 36, 93–98. [Google Scholar] [CrossRef]
- Kant, G.J.; Leu, J.R.; Anderson, S.M.; Mougey, E.H. Effects of Chronic Stress on Plasma Corticosterone, ACTH and Prolactin. Physiol. Behav. 1987, 40, 775–779. [Google Scholar] [CrossRef]
- Hur, K.Y.; Lee, M.-S. Gut Microbiota and Metabolic Disorders. Diabetes Metab. J. 2015, 39, 198. [Google Scholar] [CrossRef] [Green Version]
- Falcinelli, S.; Picchietti, S.; Rodiles, A.; Cossignani, L.; Merrifield, D.L.; Taddei, A.R.; Maradonna, F.; Olivotto, I.; Gioacchini, G.; Carnevali, O. Lactobacillus Rhamnosus Lowers Zebrafish Lipid Content by Changing Gut Microbiota and Host Transcription of Genes Involved in Lipid Metabolism. Sci. Rep. 2015, 5, 9336. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Shi, B. Gut Microbiota as a Potential Target of Metabolic Syndrome: The Role of Probiotics and Prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Identification | Mean | p Value | Fold Change | ||||
|---|---|---|---|---|---|---|---|---|
| C | A | A+P | A/C | A/A+P | A/C | A/A+P | ||
| G1 | CE (22:6) | 6 ± 3 | 57 ± 20 | 7 ± 1 | 0.004 | 0.005 | 8.97 | 8.38 |
| CE (20:4) | 12 ± 3 | 43 ± 7 | 14 ± 2 | 0.000 | 0.001 | 3.69 | 3.08 | |
| Sterol derivatives | 42 ± 13 | 135 ± 37 | 49 ± 8 | 0.003 | 0.005 | 3.22 | 2.77 | |
| SM (d18:1/24:1) | 120 ± 27 | 343 ± 66 | 122 ± 17 | 0.001 | 0.001 | 2.86 | 2.82 | |
| SM (d16:1/18:0) | 207 ± 44 | 563 ± 110 | 223 ± 44 | 0.001 | 0.001 | 2.72 | 2.52 | |
| LysoPC (18:0) | 894 ± 160 | 2136 ± 356 | 840 ± 50 | 0.001 | 0.001 | 2.39 | 2.54 | |
| PC (18:0/20:4) | 1561 ± 302 | 3524 ± 573 | 1830 ± 339 | 0.001 | 0.001 | 2.26 | 1.93 | |
| PC (16:0/18:0) | 66 ± 11 | 148 ± 27 | 73 ± 12 | 0.001 | 0.002 | 2.24 | 2.04 | |
| PC (16:0/16:0) | 54 ± 8 | 118 ± 14 | 62 ± 10 | 0.000 | 0.000 | 2.17 | 1.91 | |
| Cholesterol | 217 ± 53 | 445 ± 112 | 229 ± 17 | 0.007 | 0.012 | 2.05 | 1.94 | |
| LysoPC (16:0) | 1484 ± 205 | 2893 ± 284 | 1385 ± 131 | 0.000 | 0.000 | 1.95 | 2.09 | |
| TG (60:12) | 25 ± 9 | 201 ± 64 | 22 ± 13 | 0.003 | 0.003 | 8.20 | 9.32 | |
| TG (60:11) | 56 ± 15 | 221 ± 75 | 43 ± 20 | 0.007 | 0.005 | 3.92 | 5.20 | |
| TG (58:10) | 88 ± 26 | 287 ± 71 | 83 ± 25 | 0.002 | 0.002 | 3.28 | 3.46 | |
| TG (60:10) | 39 ± 6 | 108 ± 33 | 30 ± 10 | 0.009 | 0.005 | 2.74 | 3.64 | |
| PC (18:0/22:6) | 279 ± 75 | 687 ± 244 | 285 ± 23 | 0.018 | 0.021 | 2.46 | 2.41 | |
| G2 | TG (48:2) | 159 ± 18 | 16 ± 6 | 175 ± 51 | 0.000 | 0.002 | 0.10 | 0.09 |
| TG (48:1) | 112 ± 16 | 15 ± 8 | 124 ± 51 | 0.000 | 0.008 | 0.14 | 0.12 | |
| TG (51:2) | 148 ± 15 | 21 ± 7 | 155 ± 20 | 0.000 | 0.000 | 0.14 | 0.14 | |
| TG (51:3) | 144 ± 22 | 20 ± 7 | 156 ± 12 | 0.000 | 0.000 | 0.14 | 0.13 | |
| TG (53:3) | 133 ± 24 | 24 ± 8 | 134 ± 15 | 0.000 | 0.000 | 0.18 | 0.18 | |
| TG (54:2) | 443 ± 51 | 116 ± 34 | 410 ± 89 | 0.000 | 0.001 | 0.26 | 0.28 | |
| TG (50:2) | 1177 ± 140 | 356 ± 102 | 1290 ± 210 | 0.000 | 0.000 | 0.30 | 0.28 | |
| TG (54:3) | 1296 ± 143 | 430 ± 77 | 1288 ± 131 | 0.000 | 0.000 | 0.33 | 0.33 | |
| TG (52:1) | 231 ± 25 | 84 ± 23 | 203 ± 48 | 0.000 | 0.003 | 0.36 | 0.41 | |
| TG (50:1) | 562 ± 72 | 209 ± 63 | 579 ± 161 | 0.000 | 0.004 | 0.37 | 0.36 | |
| TG (56:3) | 163 ± 27 | 42 ± 22 | 148 ± 36 | 0.000 | 0.001 | 0.26 | 0.28 | |
| TG (50:3) | 830 ± 83 | 179 ± 67 | 945 ± 112 | 0.000 | 0.000 | 0.22 | 0.19 | |
| DG (34:1) | 184 ± 18 | 61 ± 18 | 191 ± 15 | 0.000 | 0.000 | 0.33 | 0.32 | |
| TG (52:5) | 489 ± 101 | 179 ± 50 | 549 ± 118 | 0.000 | 0.001 | 0.37 | 0.33 | |
| TG (52:2) | 3124 ± 323 | 1189 ± 347 | 3363 ± 272 | 0.000 | 0.000 | 0.38 | 0.35 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, Y.; Nam, M.H.; Kim, E. Relationship Between the Gastrointestinal Side Effects of an Anti-Hypertensive Medication and Changes in the Serum Lipid Metabolome. Nutrients 2020, 12, 205. https://doi.org/10.3390/nu12010205
Ahn Y, Nam MH, Kim E. Relationship Between the Gastrointestinal Side Effects of an Anti-Hypertensive Medication and Changes in the Serum Lipid Metabolome. Nutrients. 2020; 12(1):205. https://doi.org/10.3390/nu12010205
Chicago/Turabian StyleAhn, Yoomin, Myung Hee Nam, and Eungbin Kim. 2020. "Relationship Between the Gastrointestinal Side Effects of an Anti-Hypertensive Medication and Changes in the Serum Lipid Metabolome" Nutrients 12, no. 1: 205. https://doi.org/10.3390/nu12010205