Z-ajoene from Crushed Garlic Alleviates Cancer-Induced Skeletal Muscle Atrophy

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Ajoene Extract and Purification of Z-Ajoene from Garlic
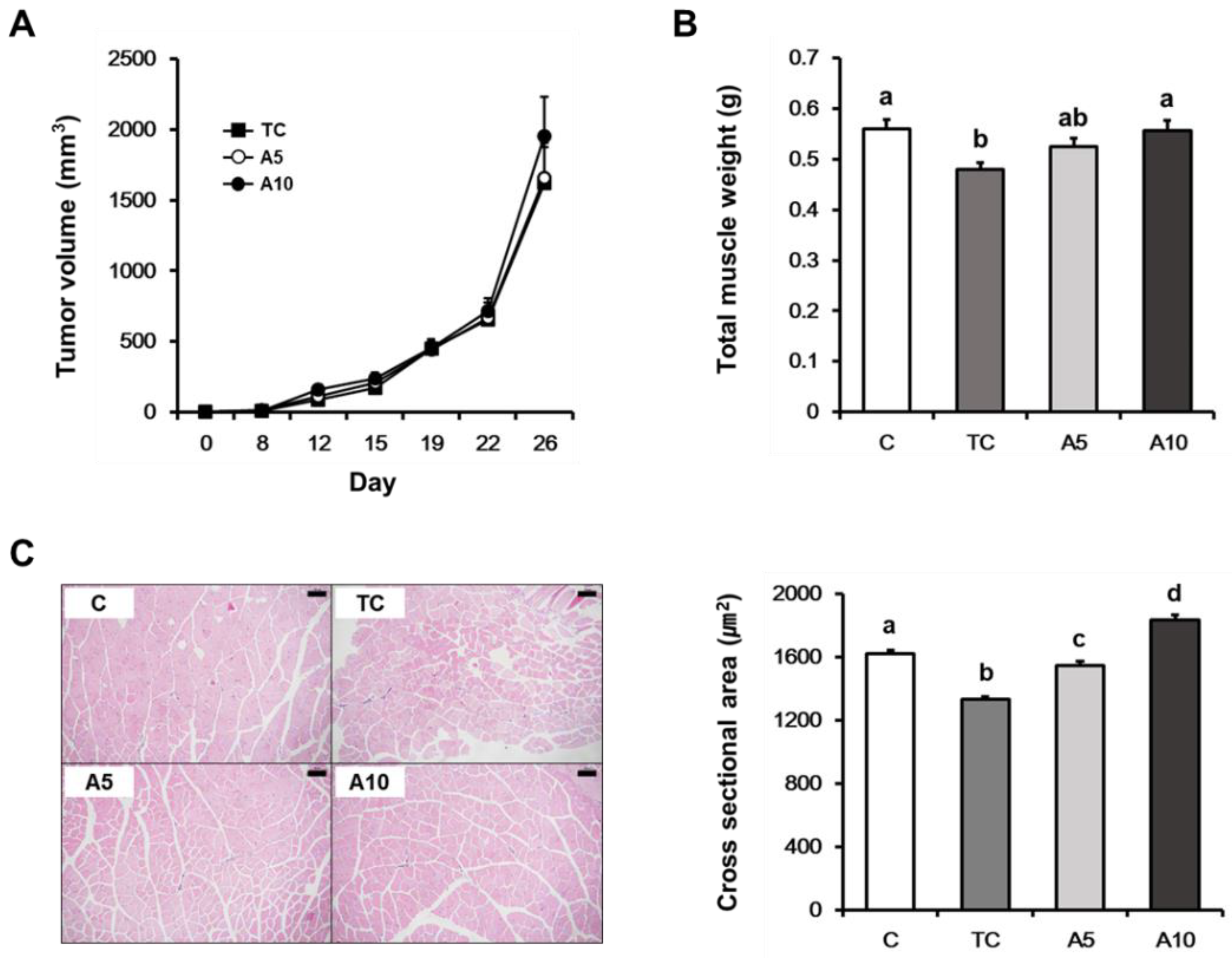
2.2. Mouse Model of Cancer Cachexia
2.3. Assessment of Muscle Cross-Sectional Area
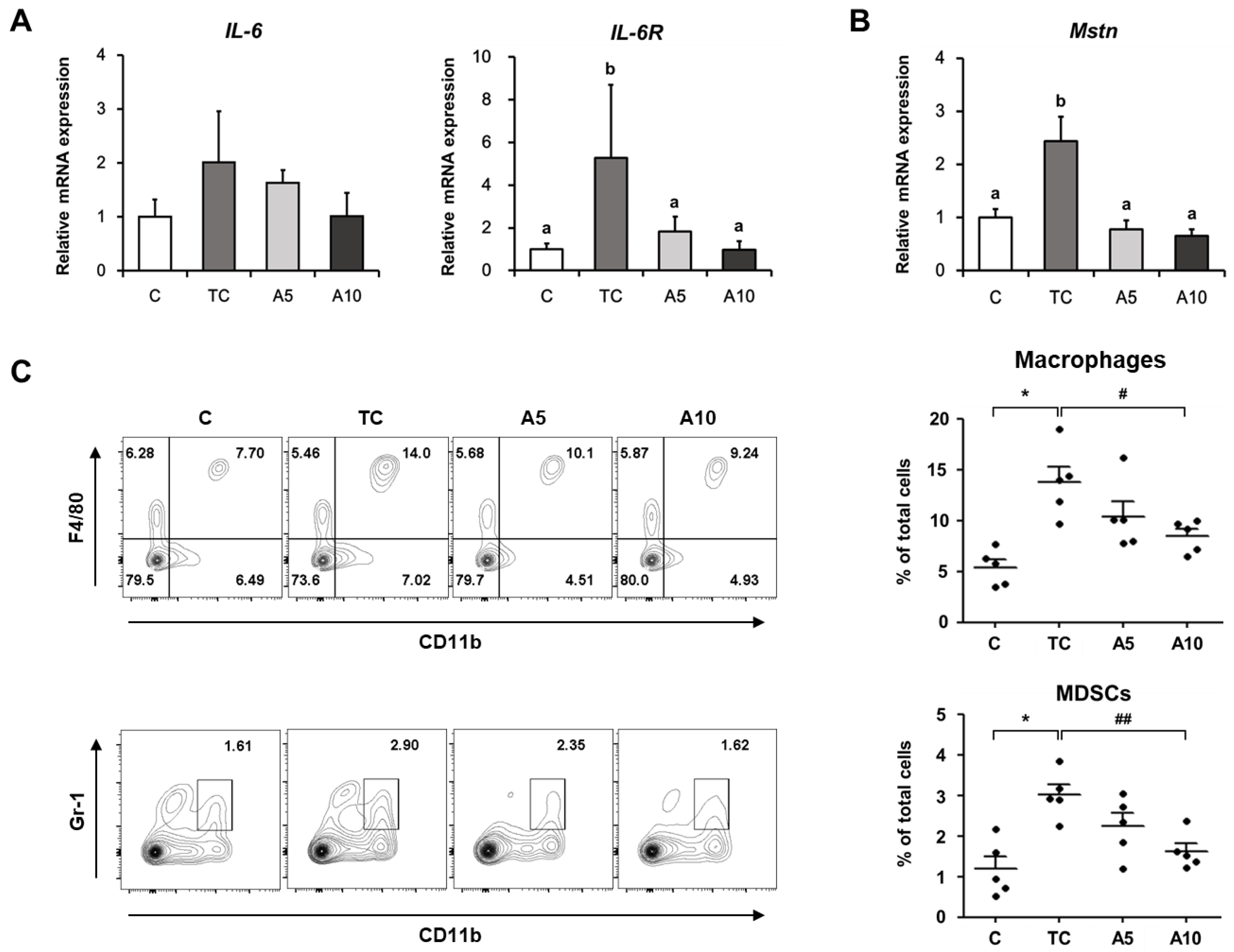
2.4. Flow Cytometry
2.5. Cell Culture, Myoblast Differentiation and Collection of Conditioned Medium of CT26 Cancer Cells
2.6. Immunostaining of MHC
2.7. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction Analysis (qRT-PCR)
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
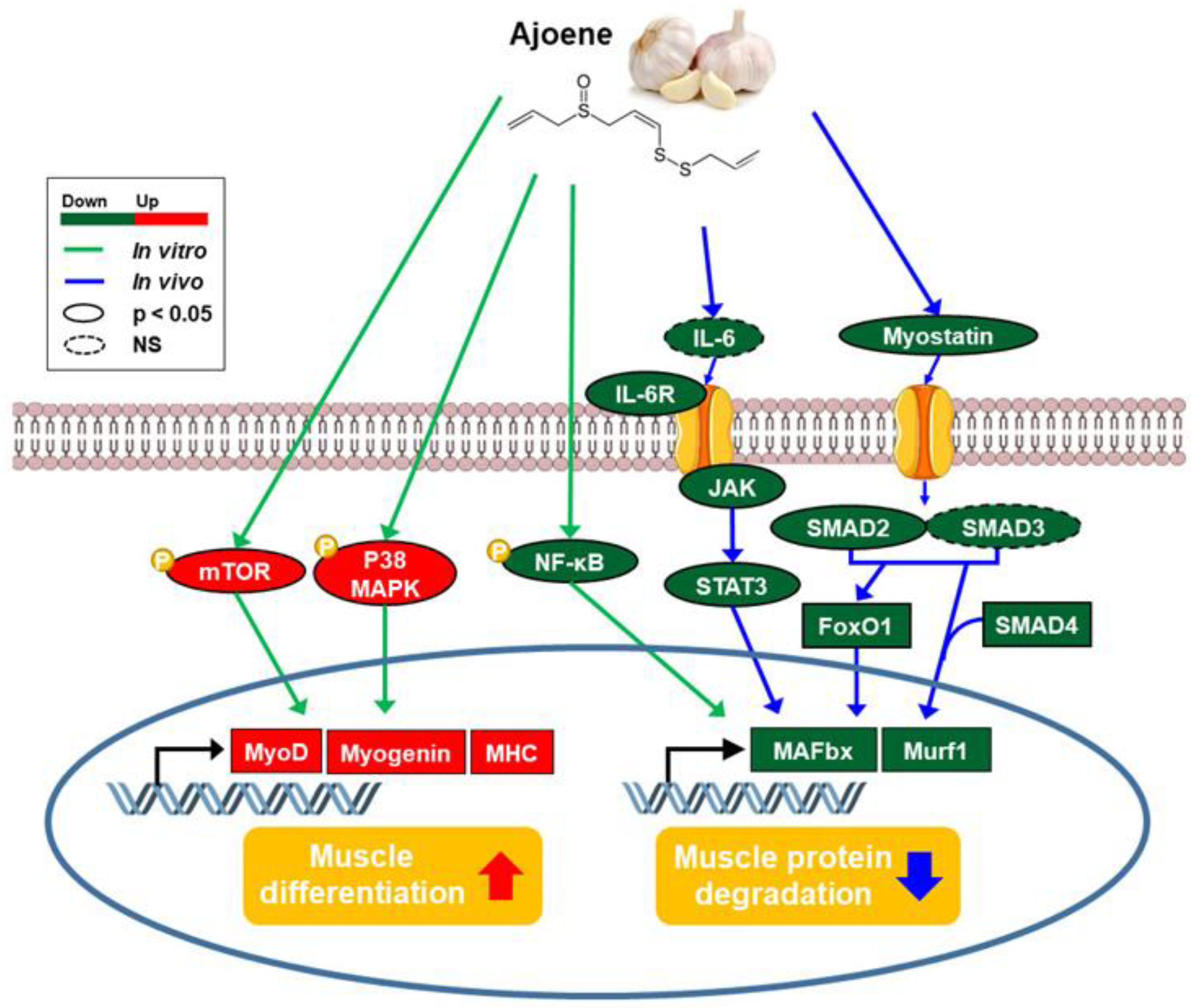
3.1. Ajoene Extract of Garlic Attenuates Cancer-Induced Muscle Atrophy in CT26 Tumor-Bearing Mice
3.2. Ajoene Extract Suppresses Muscle Wasting by Reducing Myokines Secretion in CT26 Tumor-Bearing Mice
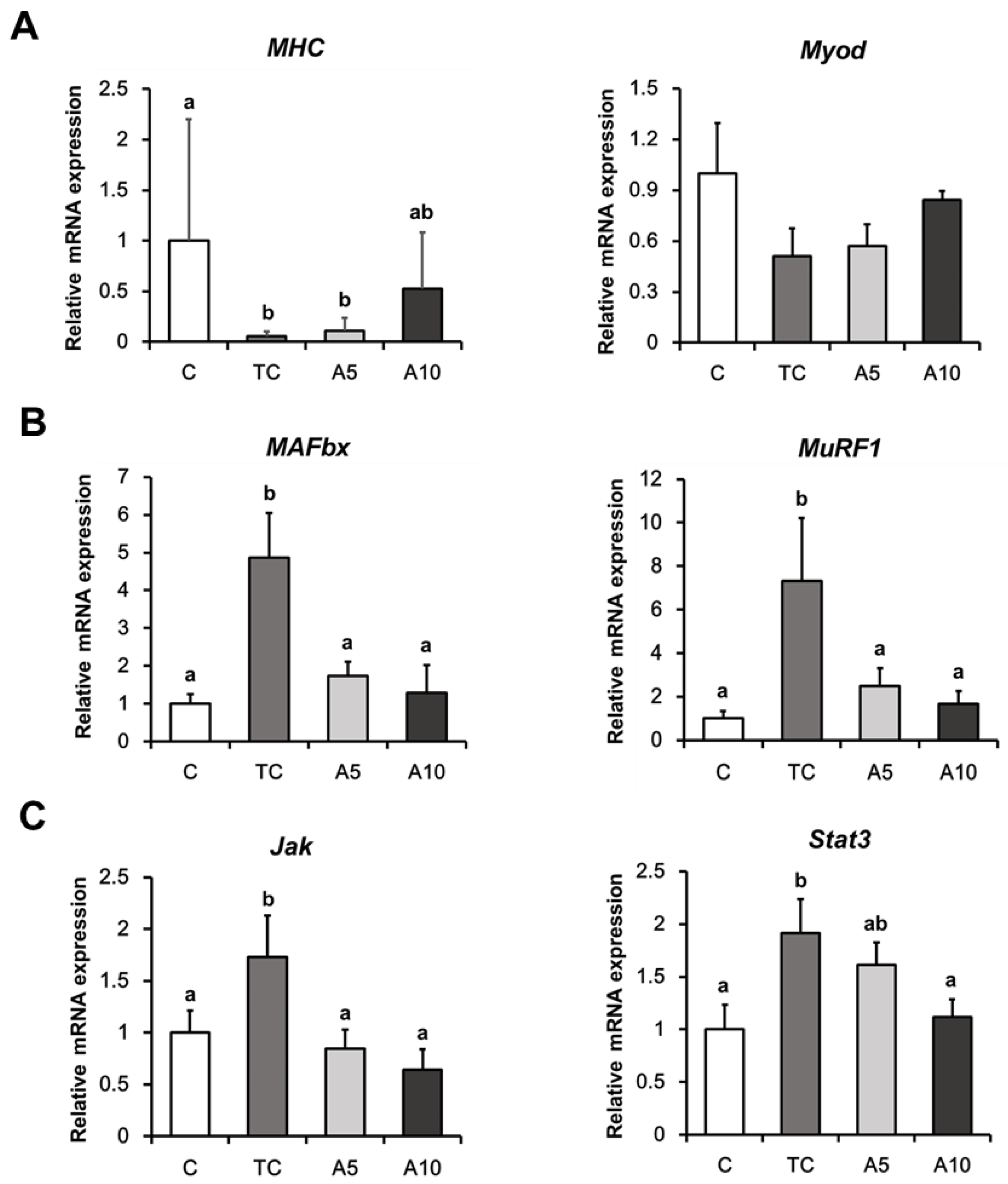
3.3. Ajoene Extract Inhibits Muscle Degradation by Down-Regulating JAK/STAT3 and SMADs/FoxO Signaling Pathways In CT26 Tumor-Bearing Mice
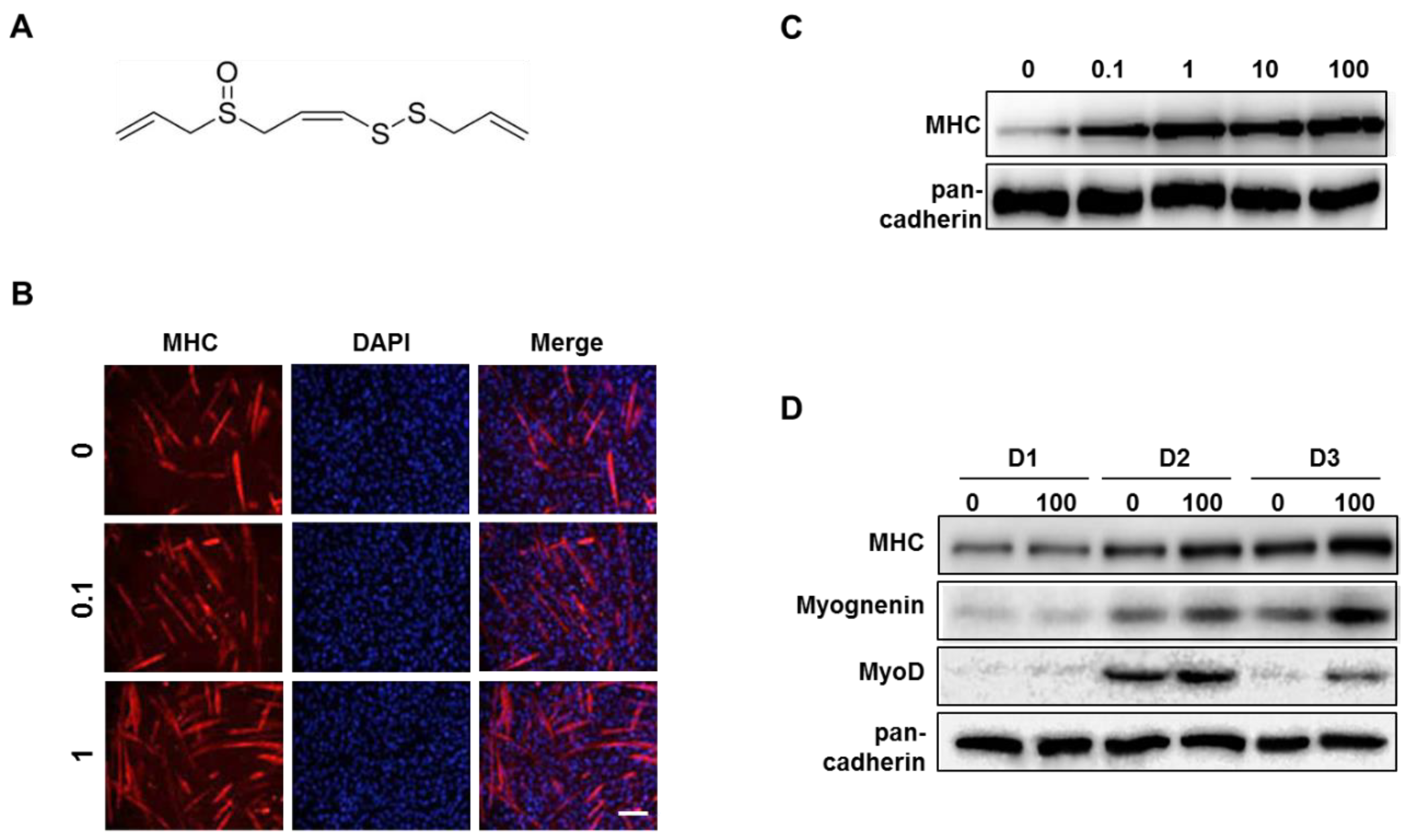
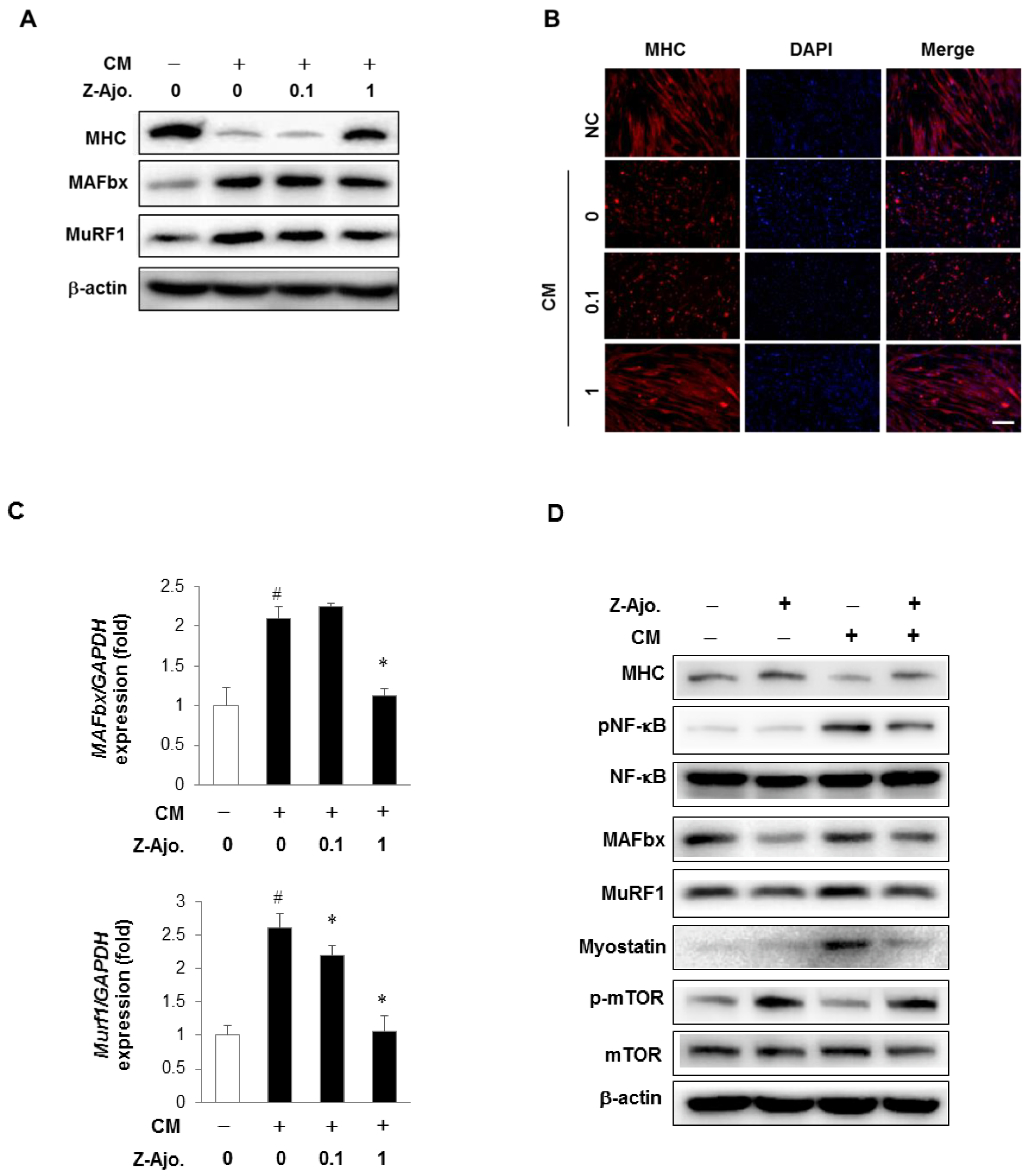
3.4. Z-ajoene Stimulates Myogenesis
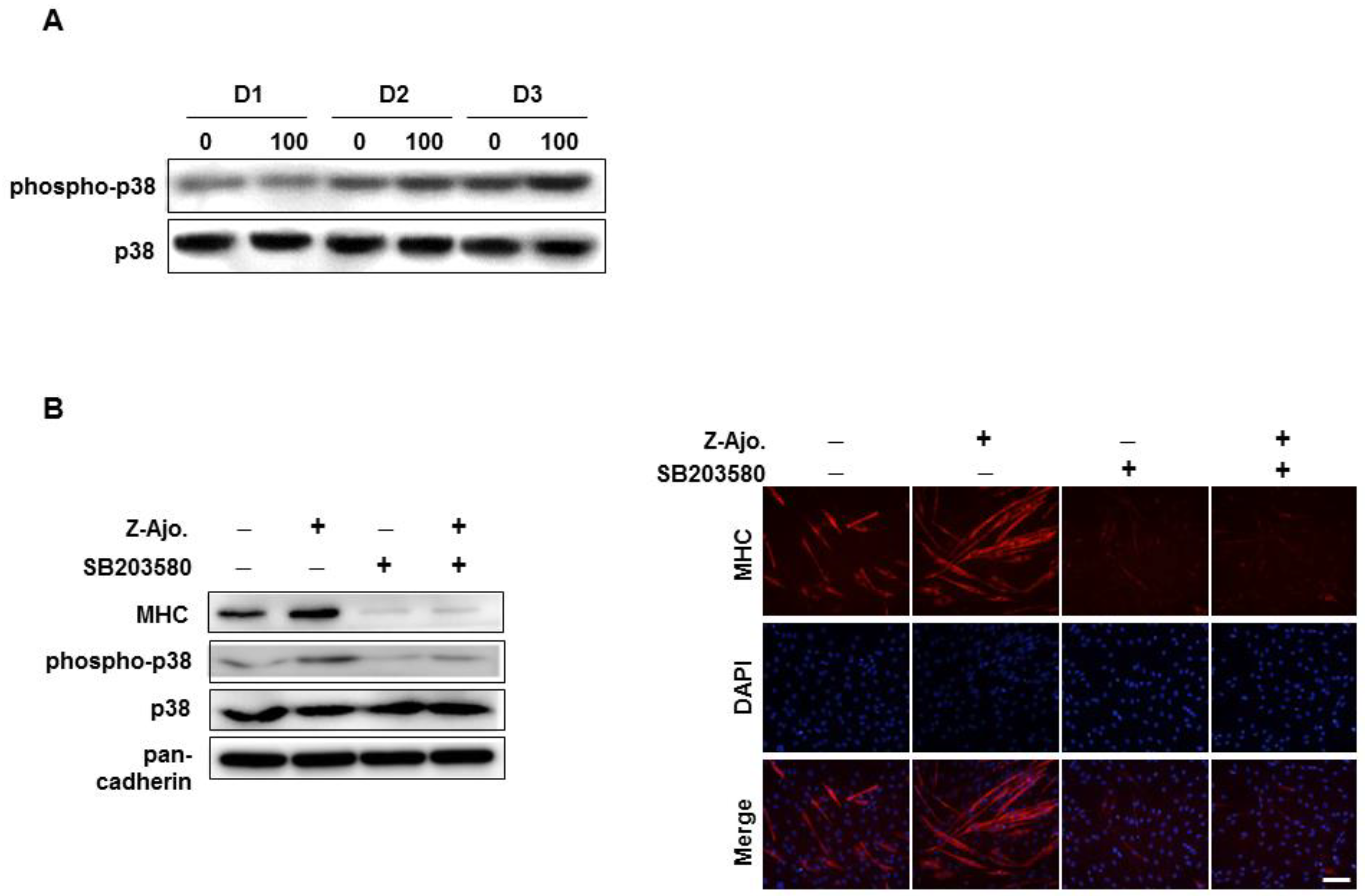
3.5. Z-ajoene Activates p38 MAPK During Myogenesis
3.6. Z-ajoene Prevents Myotube Protein Loss in Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porporato, P.E. Understanding cachexia as a cancer metabolism syndrome. Oncogenesis 2016, 5, e200. [Google Scholar] [CrossRef] [PubMed]
- Argiles, J.M.; Busquets, S.; Stemmler, B.; Lopez-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.F.; Rohm, M.; Herzig, S.; Berriel Diaz, M. Cancer cachexia: More than skeletal muscle wasting. Trends Cancer 2018, 4, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, M.J. Cachexia in cancer patients. Nat. Rev. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Fearon, K.C.; Glass, D.J.; Guttridge, D.C. Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Sandri, M. Protein breakdown in muscle wasting: Role of autophagy-lysosome and ubiquitin-proteasome. Int. J. Biochem. Cell Biol. 2013, 45, 2121–2129. [Google Scholar] [CrossRef] [Green Version]
- Carson, J.A.; Baltgalvis, K.A. Interleukin 6 as a key regulator of muscle mass during cachexia. Exerc. Sport Sci. Rev. 2010, 38, 168–176. [Google Scholar] [CrossRef]
- Lokireddy, S.; Mouly, V.; Butler-Browne, G.; Gluckman, P.D.; Sharma, M.; Kambadur, R.; McFarlane, C. Myostatin promotes the wasting of human myoblast cultures through promoting ubiquitin-proteasome pathway-mediated loss of sarcomeric proteins. Am. J. Physiol. Cell Physiol. 2011, 301, C1316–C1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Wang, J.L.; Lu, J.; Song, Y.; Kwak, K.S.; Jiao, Q.; Rosenfeld, R.; Chen, Q.; Boone, T.; Simonet, W.S.; et al. Reversal of cancer cachexia and muscle wasting by ActRIIB antagonism leads to prolonged survival. Cell 2010, 142, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Takahashi, F.; Kato, M.; Kaneko, N.; Doi, T.; Ohe, Y.; Koizumi, F.; Nishio, K.; Takahashi, K. Tocilizumab, a proposed therapy for the cachexia of Interleukin6-expressing lung cancer. PLoS ONE 2014, 9, e102436. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, T.J.; Smith, J.T.; Schuster, M.; Dragnev, K.H.; Rigas, J.R. A humanized anti-IL-6 antibody (ALD518) in non-small cell lung cancer. Expert Opin. Biol. Ther. 2011, 11, 1663–1668. [Google Scholar] [CrossRef]
- Argiles, J.M.; Lopez-Soriano, F.J.; Stemmler, B.; Busquets, S. Therapeutic strategies against cancer cachexia. Eur. J. Transl. Myol. 2019, 29, 7960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazibuko-Mbeje, S.E.; Dludla, P.V.; Nkambule, B.B.; Obonye, N.; Louw, J. Skeletal muscle as a therapeutic target for natural products to reverse metabolic syndrome. Muscle Cell Tissue Curr. Status Res. Field 2018, 175. [Google Scholar] [CrossRef]
- Pandrangi, A. Cancer chemoprevention by garlic: A review. Hered. Genet. 2015, 4, 1–7. [Google Scholar]
- Hwang, K.A.; Hwang, Y.J.; Hwang, I.G.; Heo, W.; Kim, Y.J. Effects of Low Temperature-Aged Garlic on Exercise Performance and Fatigue in Mice. J. Med. Food 2019, 22, 944–951. [Google Scholar] [CrossRef]
- Bhandari, P.R. Garlic (Allium sativum L.): A review of potential therapeutic applications. Int. J. Green Pharm. 2012, 6, 118–129. [Google Scholar] [CrossRef]
- Bayan, L.; Koulivand, P.H.; Gorji, A. Garlic: A review of potential therapeutic effects. Avicenna J. Phytomed. 2014, 4, 1–14. [Google Scholar]
- Dušica, I. Allicin and related compounds: Biosynthesis, synthesis and pharmacological activity. Facta Univ. 2012, 9, 9–20. [Google Scholar]
- Kay, H.Y.; Yang, J.W.; Kim, T.H.; Lee, D.Y.; Kang, B.; Ryu, J.H.; Jeon, R.; Kim, S.G. Ajoene, a stable garlic by-product, has an antioxidant effect through Nrf2-mediated glutamate-cysteine ligase induction in HepG2 cells and primary hepatocytes. J. Nutr. 2010, 140, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Ryu, J.H.; Surh, Y.J. Ajoene, a major organosulfide found in crushed garlic, induces NAD(P)H:quinone oxidoreductase expression through nuclear factor E2-related factor-2 activation in human breast epithelial cells. J. Cancer Prev. 2019, 24, 112–122. [Google Scholar] [CrossRef]
- Lee, D.Y.; Li, H.; Lim, H.J.; Lee, H.J.; Jeon, R.; Ryu, J.H. Anti-inflammatory activity of sulfur-containing compounds from garlic. J. Med. Food 2012, 15, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Al-Sawaf, O.; Fragoulis, A.; Rosen, C.; Keimes, N.; Liehn, E.A.; Hölzle, F.; Kan, Y.W.; Pufe, T.; Sönmez, T.T.; Wruck, C.J. Nrf2 augments skeletal muscle regeneration after ischaemia-reperfusion injury. J. Pathol. 2014, 234, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.E.; Cole, P.D.; Cho, R.C.; Ly, A.; Ishiguro, L.; Sohn, K.J.; Croxford, R.; Kamen, B.A.; Kim, Y.I. gamma-Glutamyl hydrolase modulation and folate influence chemosensitivity of cancer cells to 5-fluorouracil and methotrexate. Br. J. Cancer 2013, 109, 2175–2188. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academy of Sciences: Washington, DC, USA, 1996. [Google Scholar]
- Lee, J.H.; Jun, H.S. Role of myokines in regulating skeletal muscle mass and function. Front. Physiol. 2019, 10, 42. [Google Scholar] [CrossRef]
- Mueller, T.C.; Bachmann, J.; Prokopchuk, O.; Friess, H.; Martignoni, M.E. Molecular pathways leading to loss of skeletal muscle mass in cancer cachexia—Can findings from animal models be translated to humans? BMC Cancer 2016, 16, 75. [Google Scholar] [CrossRef]
- Rose-John, S. IL-6 trans-signaling via the soluble IL-6 receptor: Importance for the pro-inflammatory activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef]
- Lesina, M.; Kurkowski, M.U.; Ludes, K.; Rose-John, S.; Treiber, M.; Kloppel, G.; Yoshimura, A.; Reindl, W.; Sipos, B.; Akira, S.; et al. Stat3/Socs3 activation by IL-6 transsignaling promotes progression of pancreatic intraepithelial neoplasia and development of pancreatic cancer. Cancer Cell 2011, 19, 456–469. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Nishikata, R.; Senju, S.; Nishimura, Y. Myeloid-derived suppressor cells attenuate TH1 development through IL-6 production to promote tumor progression. Cancer Immunol. Res. 2013, 1, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Winfield, R.D.; Delano, M.J.; Pande, K.; Scumpia, P.O.; Laface, D.; Moldawer, L.L. Myeloid-derived suppressor cells in cancer cachexia syndrome: A new explanation for an old problem. J Parenter. Enteral. Nutr. 2008, 32, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkina, Y.; von Haehling, S.; Anker, S.D.; Springer, J. The role of myostatin in muscle wasting: An overview. J. Cachexia Sarcopenia Muscle 2011, 2, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Kollias, H.D.; McDermott, J.C. Transforming growth factor-beta and myostatin signaling in skeletal muscle. J. Appl. Physiol. (1985) 2008, 104, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, S.J.; Bae, G.U.; Baek, N.I.; Ryu, J.H. Canadine from Corydalis turtschaninovii stimulates myoblast differentiation and protects against myotube atrophy. Int. J. Mol. Sci. 2017, 18, 2748. [Google Scholar] [CrossRef]
- Oeztuerk-Winder, F.; Ventura, J.J. The many faces of p38 mitogen-activated protein kinase in progenitor/stem cell differentiation. Biochem. J. 2012, 445, 1–10. [Google Scholar] [CrossRef]
- Kim, A.; Im, M.; Gu, M.J.; Ma, J.Y. Citrus unshiu peel extract alleviates cancer-induced weight loss in mice bearing CT-26 adenocarcinoma. Sci. Rep. 2016, 6, 24214. [Google Scholar] [CrossRef]
- Dutt, V.; Gupta, S.; Dabur, R.; Injeti, E.; Mittal, A. Skeletal muscle atrophy: Potential therapeutic agents and their mechanisms of action. Pharmacol. Res. 2015, 99, 86–100. [Google Scholar] [CrossRef]
- Jeong, S.M.; Seo, B.K.; Park, Y.C.; Baek, Y.H. A Review of Complementary and Alternative Medicine Therapies on Muscular Atrophy: A Literature Review of In Vivo/In Vitro Studies. Evid. Based Complement. Alternat. Med. 2018, 2018, 8654719. [Google Scholar] [CrossRef]
- Hamilton, M.T. The role of skeletal muscle contractile duration throughout the whole day: Reducing sedentary time and promoting universal physical activity in all people. J. Physiol. 2018, 596, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Advani, S.M.; Advani, P.G.; VonVille, H.M.; Jafri, S.H. Pharmacological management of cachexia in adult cancer patients: A systematic review of clinical trials. BMC Cancer 2018, 18, 1174. [Google Scholar] [CrossRef] [PubMed]
- Penna, F.; Busquets, S.; Argiles, J.M. Experimental cancer cachexia: Evolving strategies for getting closer to the human scenario. Semin. Cell Dev. Biol. 2016, 54, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Aulino, P.; Berardi, E.; Cardillo, V.M.; Rizzuto, E.; Perniconi, B.; Ramina, C.; Padula, F.; Spugnini, E.P.; Baldi, A.; Faiola, F.; et al. Molecular, cellular and physiological characterization of the cancer cachexia-inducing C26 colon carcinoma in mouse. BMC Cancer 2010, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Tocmo, R.; Wu, Y.; Liang, D.; Fogliano, V.; Huang, D. Boiling enriches the linear polysulfides and the hydrogen sulfide-releasing activity of garlic. Food Chem. 2017, 221, 1867–1873. [Google Scholar] [CrossRef]
- Kim, J.W.; Ku, S.K.; Han, M.H.; Kim, K.Y.; Kim, S.G.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, C.M.; Choi, Y.H. The administration of Fructus Schisandrae attenuates dexamethasone-induced muscle atrophy in mice. Int. J. Mol. Med. 2015, 36, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Eddins, M.J.; Marblestone, J.G.; Suresh Kumar, K.G.; Leach, C.A.; Sterner, D.E.; Mattern, M.R.; Nicholson, B. Targeting the ubiquitin E3 ligase MuRF1 to inhibit muscle atrophy. Cell Biochem. Biophys. 2011, 60, 113–118. [Google Scholar] [CrossRef]
- Bowen, T.S.; Adams, V.; Werner, S.; Fischer, T.; Vinke, P.; Brogger, M.N.; Mangner, N.; Linke, A.; Sehr, P.; Lewis, J.; et al. Small-molecule inhibition of MuRF1 attenuates skeletal muscle atrophy and dysfunction in cardiac cachexia. J. Cachexia Sarcopenia Muscle 2017, 8, 939–953. [Google Scholar] [CrossRef]
- McFarlane, C.; Plummer, E.; Thomas, M.; Hennebry, A.; Ashby, M.; Ling, N.; Smith, H.; Sharma, M.; Kambadur, R. Myostatin induces cachexia by activating the ubiquitin proteolytic system through an NF-kappaB-independent, FoxO1-dependent mechanism. J. Cell. Physiol. 2006, 209, 501–514. [Google Scholar] [CrossRef]
- Sriram, S.; Subramanian, S.; Juvvuna, P.K.; Ge, X.; Lokireddy, S.; McFarlane, C.D.; Wahli, W.; Kambadur, R.; Sharma, M. Myostatin augments muscle-specific ring finger protein-1 expression through an NF-kB independent mechanism in SMAD3 null muscle. Mol. Endocrinol. 2014, 28, 317–330. [Google Scholar] [CrossRef]
- Samant, S.A.; Kanwal, A.; Pillai, V.B.; Bao, R.; Gupta, M.P. The histone deacetylase SIRT6 blocks myostatin expression and development of muscle atrophy. Sci. Rep. 2017, 7, 11877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Mallidis, C.; Artaza, J.; Taylor, W.; Gonzalez-Cadavid, N.; Bhasin, S. Characterization of 5’-regulatory region of human myostatin gene: Regulation by dexamethasone in vitro. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E1128–E1136. [Google Scholar] [CrossRef] [PubMed]
- Dedieu, S.; Mazeres, G.; Cottin, P.; Brustis, J.J. Involvement of myogenic regulator factors during fusion in the cell line C2C12. Int. J. Dev. Biol. 2002, 46, 235–241. [Google Scholar] [PubMed]
- Ferri, P.; Barbieri, E.; Burattini, S.; Guescini, M.; D’Emilio, A.; Biagiotti, L.; Del Grande, P.; De Luca, A.; Stocchi, V.; Falcieri, E. Expression and subcellular localization of myogenic regulatory factors during the differentiation of skeletal muscle C2C12 myoblasts. J. Cell. Biochem. 2009, 108, 1302–1317. [Google Scholar] [CrossRef]
- Lassar, A.B. The p38 MAPK family, a pushmi-pullyu of skeletal muscle differentiation. J. Cell. Biol. 2009, 187, 941–943. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | TC | A5 | A10 | p-Value | |
|---|---|---|---|---|---|
| Gastrocnemius (g) | 0.1297 ± 0.0048 | 0.1244 ± 0.0029 | 0.1278 ± 0.0038 | 0.1324 ± 0.0031 | 0.543 |
| Tibialis (g) | 0.0256 ± 0.0038 | 0.0192 ± 0.0030 | 0.0237 ± 0.0026 | 0.0236 ± 0.0018 | 0.509 |
| Extensor digitorum longus (g) | 0.0161 ± 0.0008 | 0.0143 ± 0.0005 | 0.0167 ± 0.0008 | 0.0144 ± 0.0007 | 0.058 |
| Soleus (g) | 0.0082 ± 0.0008 | 0.0088 ± 0.0004 | 0.0080 ± 0.0008 | 0.0092 ± 0.0009 | 0.662 |
| Quadriceps (g) | 0.1365 ± 0.0068 | 0.1172 ± 0.0050 | 0.1349 ± 0.0062 | 0.1402 ± 0.0080 | 0.109 |
| Extra muscle (g) | 0.2269 ± 0.0105 | 0.1902 ± 0.0112 | 0.2175 ± 0.0142 | 0.2350 ± 0.0142 | 0.101 |
| Total muscle (g) | 0.5592 ± 0.0183 a | 0.4804 ± 0.0131 b | 0.5254 ± 0.0158 ab | 0.5562 ± 0.0196 a | 0.011 |
| Heart (g) | 0.13 ± 0.00 | 0.13 ± 0.01 | 0.12 ± 0.00 | 0.12 ± 0.00 | 0.394 |
| Spleen (g) | 0.12 ± 0.01 a | 0.28 ± 0.02 b | 0.27 ± 0.02 b | 0.27 ± 0.02 b | <0.0001 |
| Liver (g) | 1.10 ± 0.06 a | 1.35 ± 0.05 b | 1.41 ± 0.05 b | 1.48 ± 0.04 b | <0.0001 |
| Epididymal fat (g) | 0.34 ± 0.02 | 0.25 ± 0.03 | 0.27 ± 0.03 | 0.30 ± 0.03 | 0.127 |
| Mesenteric fat (g) | 0.21 ± 0.02 | 0.19 ± 0.02 | 0.18 ± 0.02 | 0.17 ± 0.02 | 0.452 |
| Perirenal fat (g) | 0.09 ± 0.01 a | 0.05 ± 0.00 b | 0.07 ± 0.01 ab | 0.06 ± 0.01 ab | 0.020 |
| Total fat (g) | 0.65 ± 0.04 | 0.46 ± 0.05 | 0.52 ± 0.06 | 0.55 ± 0.06 | 0.193 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Heo, J.-W.; Kim, A.-R.; Kweon, M.; Nam, S.; Lim, J.-S.; Sung, M.-K.; Kim, S.-E.; Ryu, J.-H. Z-ajoene from Crushed Garlic Alleviates Cancer-Induced Skeletal Muscle Atrophy. Nutrients 2019, 11, 2724. https://doi.org/10.3390/nu11112724
Lee H, Heo J-W, Kim A-R, Kweon M, Nam S, Lim J-S, Sung M-K, Kim S-E, Ryu J-H. Z-ajoene from Crushed Garlic Alleviates Cancer-Induced Skeletal Muscle Atrophy. Nutrients. 2019; 11(11):2724. https://doi.org/10.3390/nu11112724
Chicago/Turabian StyleLee, Hyejin, Ji-Won Heo, A-Reum Kim, Minson Kweon, Sorim Nam, Jong-Seok Lim, Mi-Kyung Sung, Sung-Eun Kim, and Jae-Ha Ryu. 2019. "Z-ajoene from Crushed Garlic Alleviates Cancer-Induced Skeletal Muscle Atrophy" Nutrients 11, no. 11: 2724. https://doi.org/10.3390/nu11112724