Diet: Cause or Consequence of the Microbial Profile of Cholelithiasis Disease?


Abstract
:1. Introduction
2. Subjects and Methods
2.1. Participants
2.2. Nutritional Assessment
2.3. Food Groups
2.4. Anthropometric Measures
2.5. Biochemical Analysis
2.6. Bile Sample Collection and Microbiota Analysis Based on Sequencing of 16S rDNA Amplicons
2.7. Statistical Analyses
3. Results
3.1. General Characteristics of the Study Sample
3.2. Diet and Gallbladder Microbiota in Cholelithiasic Patients
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Díaz, I.; Fernández-Navarro, T.; Sánchez, B.; Margolles, A.; González, S. Mediterranean diet and faecal microbiota: A transversal study. Food Funct. 2016, 7, 2347–2356. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.C.; Lowe, E.C.; Chiang, H.; Pudlo, N.A.; Wu, M.; McNutly, N.P.; Abbott, D.W.; Henrissat, B.; Gilbert, H.J.; Bolam, D.N.; et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLoS Biol. 2011, 9, e1001221. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.C.; Koropatkin, N.M.; Smith, T.J.; Gordon, J.I. Complex glycan catabolism by the human gut microbiota: The bacteroidetes sus-like paradigm. J. Biol. Chem. 2009, 284, 24673–24677. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.P.; Rosendale, D.I.; Roberton, A.M. Prevotella enzymes involved in mucin oligosaccharide degradation and evidence for a small operon of genes expressed during growth on mucin. FEMS Microbiol Lett. 2000, 190, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broussard, J.L.; Devkota, S. The changing microbial landscape of Western society: Diet, dwellings and discordance. Mol. Metab. 2016, 5, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Moschetta, A.; Palasciano, G. Cholesterol gallstone disease. Lancet 2006, 368, 230–239. [Google Scholar] [CrossRef]
- Attili, A.F.; Scafato, E.; Marchioli, R.; Marfisis, R.M.; Festi, D. Diet and gallstones in Italy: The cross-sectional MICOL results. Hepatology 1998, 27, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10−/−mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Yokota, A.; Fukiya, S.; Islam, K.B.; Ooka, T.; Ogura, Y.; Hayashi, T.; Hagio, M.; Ishizuka, S. Is bile acid a determinant of the gut microbiota on a high-fat diet? Gut Microbes 2012, 3, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, D.P. Economic development-not all bonus. Nutr. Today 1976, 11, 6–13. [Google Scholar] [CrossRef]
- Russell, M.; Hollingsworth, D.F. Proceedings of the British Nutrition Foundation Research Conference on ‘Nutritional Problems in a Changing World-Nutrition in Britain Today and Tomorrow’. In Nutritional Problems in a Changing World; Applied Science Publishers: London, UK, 1973. [Google Scholar]
- Antonis, A.; Bersohn, I. The influence of diet on fecal lipids in South African white and Bantu prisoners. Am. J. Clin. Nutr. 1962, 11, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.; Hill, M.S.; Cummings, J.H. Effect of wheat fiber on blood lipids, fecal steroid excretion and serum iron. Am. J. Clin. Nutr. 1975, 28, 1408–1411. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yan, Q.; Luo, F.; Shang, D.; Wu, D.; Zhang, H.; Shang, X.; Kang, X.; Abdo, M.; Liu, B.; et al. Acute cholecystitis associated with infection of Enterobacteriaceae from gut microbiota. Clin. Microbiol. Infect. 2015, 21, e851–e859. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Shen, H.; Li, Z.; Meng, F.; Li, L.; Yang, J.; Chen, Y.; Bo, X.; Zhang, X.; Ni, M. Influence of the biliary system on biliary bacteria revealed by bacterial communities of the human biliary and upper digestive tracts. PLoS ONE 2016, 11, e0150519. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhang, Z.; Liu, B.; Hou, D.; Liang, Y.; Zhang, J.; Shi, P. Gut microbiota dysbiosis and bacterial community assembly associated with cholesterol gallstones in large-scale study. BMC Genom. 2013, 14, 669. [Google Scholar] [CrossRef] [PubMed]
- Saltykova, I.V.; Petrov, V.A.; Logacheva, M.D.; Ivanova, P.G.; Merzlikin, N.V.; Sazonov, A.E.; Ogorodova, L.M.; Brindley, P.J. Biliary microbiota, gallstone disease and infection with Opisthorchis felineus. PLoS Negl. Trop. Dis. 2016, 10, e0004809. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.; Valdes, L.; Salazar, N.; De los Reyes-Gavilan, C.G.; Ruas-Madiedo, P.; Gueimonde, M.; Gonzalez, S. Pilot study of diet and microbiota: Interactive associations of fibers and polyphenols with human intestinal bacteria. J. Agric. Food Chem. 2014, 62, 5330–5336. [Google Scholar] [CrossRef] [PubMed]
- Centro de Enseñanza Superior de Nutricion Humana y Dietetica (CESNID). Tablas de Composición de Alimentos por Medidas Caseras de Consumo Habitual en España; (Food composition tables in household measures commonly consumed in Spain); McGraw-Hill, Publicaciones y Ediciones de la Universidad de Barcelona: Barcelona, Spain, 2008. (In Spanish) [Google Scholar]
- United States Department of Agriculture (USDA). Agriculture Research Service, 2016 USDA National Nutrient Database for Standard References. Available online: http://www.ars.usda.gov/services/docs.htm?docid=8964 (accessed on 5 March 2018).
- Marlett, J.A.; Cheung, T.F. Database and quick methods of assessing typical dietary fiber intakes using data for 228 commonly consumed foods. J. Am. Diet. Assoc. 1997, 97, 1139–1148, 1151. [Google Scholar] [CrossRef]
- Theander, O.; Westerlund, E.A. Studies on dietary fiber. 3. Improved procedures for analysis of dietary fiber. J. Agric. Food Chem. 1986, 34, 330–336. [Google Scholar] [CrossRef]
- Seaman, J.F. Kinetics of wood saccharification: Hydrolysis of cellulose and decomposition of sugars in dilute acid at high temperature. Ind. Eng. Chem. 1945, 37, 43–52. [Google Scholar] [CrossRef]
- Neveu, V.; Perez-Jimenez, J.; Vos, F.; Crespy, V.; Du, C.L.; Mennen, L.; Knox, C.; Eisner, R.; Cruz, J.; Wishart, D.; et al. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database 2010, 2010, bap024. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Sánchez, B.; Farina, A.; Margolles, A.; Rodríguez, J.M. Characterization of the bile and gall bladder microbiota of healthy pigs. Microbiologyopen 2014, 3, 937–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, C.; Hevia, A.; Foroni, E.; Duranti, S.; Turroni, F.; Lugli, G.A.; Sanchez, B.; Martin, R.; Gueimonde, M.; Van Sinderen, D.; et al. Assessing the fecal microbiota: An optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS ONE 2013, 8, e68739. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Meehan, C.J.; Beiko, R.G. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Sarles, H.; Chabert, C.; Pommeau, Y.; Save, E.; Mouret, H.; Gérolami, A. Diet and cholesterol gallstones. A study of 101 patients with cholelithiasis compared to 101 matched controls. Am. J. Dig. Dis. 1969, 14, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Reid, J.M.; Fullmer, S.D.; Pettigrew, K.D.; Burch, T.A.; Bennett, P.H.; Miller, M.; Whedon, G.D. Nutrient intake of Pima Indian women: Relationships to diabetes mellitus and gallbladder disease. Am. J. Clin. Nutr. 1971, 24, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.; Hills, L.L.; Laby, B. Cholelithiasis: A clinical and dietary survey. Gut 1970, 11, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, J.; Francis, K.P.; DeBoer, M.; Chu, P.; Gibbs, K.; Contag, C.H. Extracellular replication of Listeria monocytogenes in the murine gall bladder. Science 2004, 303, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Margolis, J.J.; Contag, C.H. Induced biliary excretion of Listeria monocytogenes. Infect. Immun. 2006, 74, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Escobedo, G.; Marshall, J.M.; Gunn, J.S. Chronic and acute infection of the gall bladder by Salmonella Typhi: Understanding the carrier state. Nat. Rev. Microbiol. 2011, 9, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Ye, F.; Xie, L.; Yang, J.; Li, Z.; Xu, P.; Meng, F.; Li, L.; Chen, Y.; Bo, X.; et al. Metagenomic sequencing of bile from gallstone patients to identify different microbial community patterns and novel biliary bacteria. Sci. Rep. 2015, 5, 17450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, H.O. Pathogenesis of gallstones. Surg. Clin. N. Am. 1973, 53, 963–972. [Google Scholar] [CrossRef]
- Sarles, H.; Hauton, J.; Lafont, H.; Teissier, N.; Planche, N.E.; Gerolami, A. Effect of diet on the biliary cholesterol concentration in normals and gallstone patients. Clin. Chim. Act. 1968, 19, 147–155. [Google Scholar] [CrossRef]
- Admirand, W.H.; Small, D.M. The physicochemical basis of cholesterol gallstone formation in man. J. Clin. Investig. 1968, 47, 1043–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, D.M.; Rapo, S. Source of abnormal bile in patients with cholesterol gallstones. N. Engl. J. Med. 1970, 283, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Paigen, B.; Carey, M.C. Gallstone. In The Genetic Basis of Common Diseases, 2nd ed.; King, R.A., Rotter, J.I., Motulsky, A.G., Eds.; Oxford University Press: London, UK, 2002; pp. 298–335. [Google Scholar]
- Tsai, C.J.; Leitzmann, M.F.; Willett, W.C.; Giovannucci, E.L. Long-term intake of dietary fiber and decreased risk of cholecystectomy in women. Am. J. Gastroenterol. 2004, 99, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, D.K. Dynamics of cholesterol metabolism. I. Factors regulating total sterol biosynthesis and accumulation in the rat. Proc. Natl. Acad. Sci. USA 1963, 50, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Aho, V.; Arola, J.; Boyd, S.; Jokelainen, K.; Paulin, L.; Auvinen, P.; Färkkilä, M. Bile microbiota in primary sclerosing cholangitis: Impact on disease progression and development of biliary dysplasia. PLoS ONE 2017, 12, e0182924. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.B.M.S.; Fukiya, S.; Hagio, M.; Fujii, N.; Ishizuka, S.; Ooka, T.; Ogura, Y.; Hayashi, T.; Yokota, A. Bile acid is a host factor that regulates the composition of the cecal microbiota in rats. Gastroenterology 2011, 141, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Gérard, P. Metabolism of cholesterol and bile acids by the gut microbiota. Pathogens 2013, 3, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Chang, E.B. Interactions between diet, bile acid metabolism, gut microbiota, and Inflammatory Bowel Diseases. Dig. Dis. 2015, 33, 351–356. [Google Scholar] [CrossRef] [PubMed]



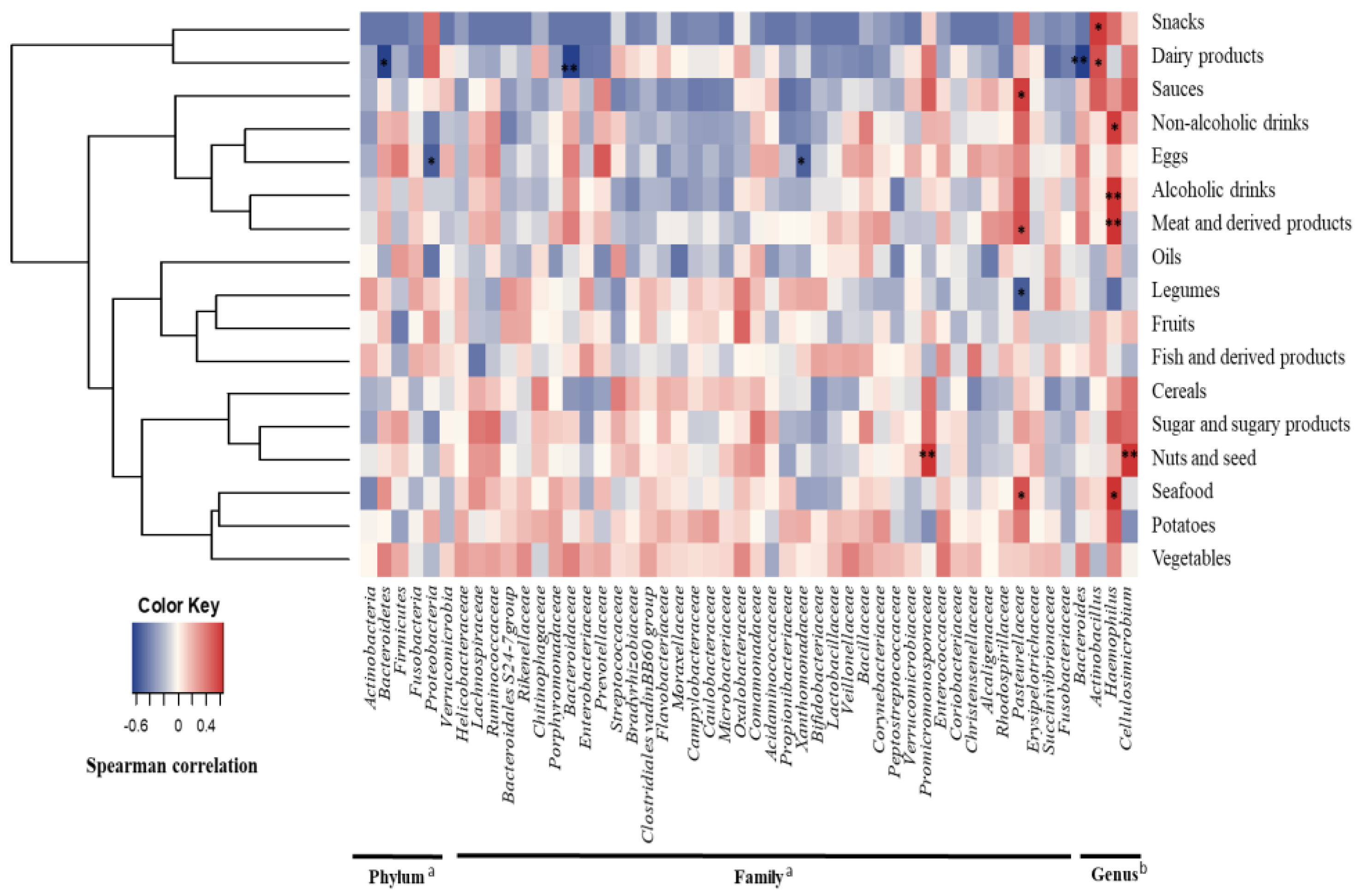{kind=link}
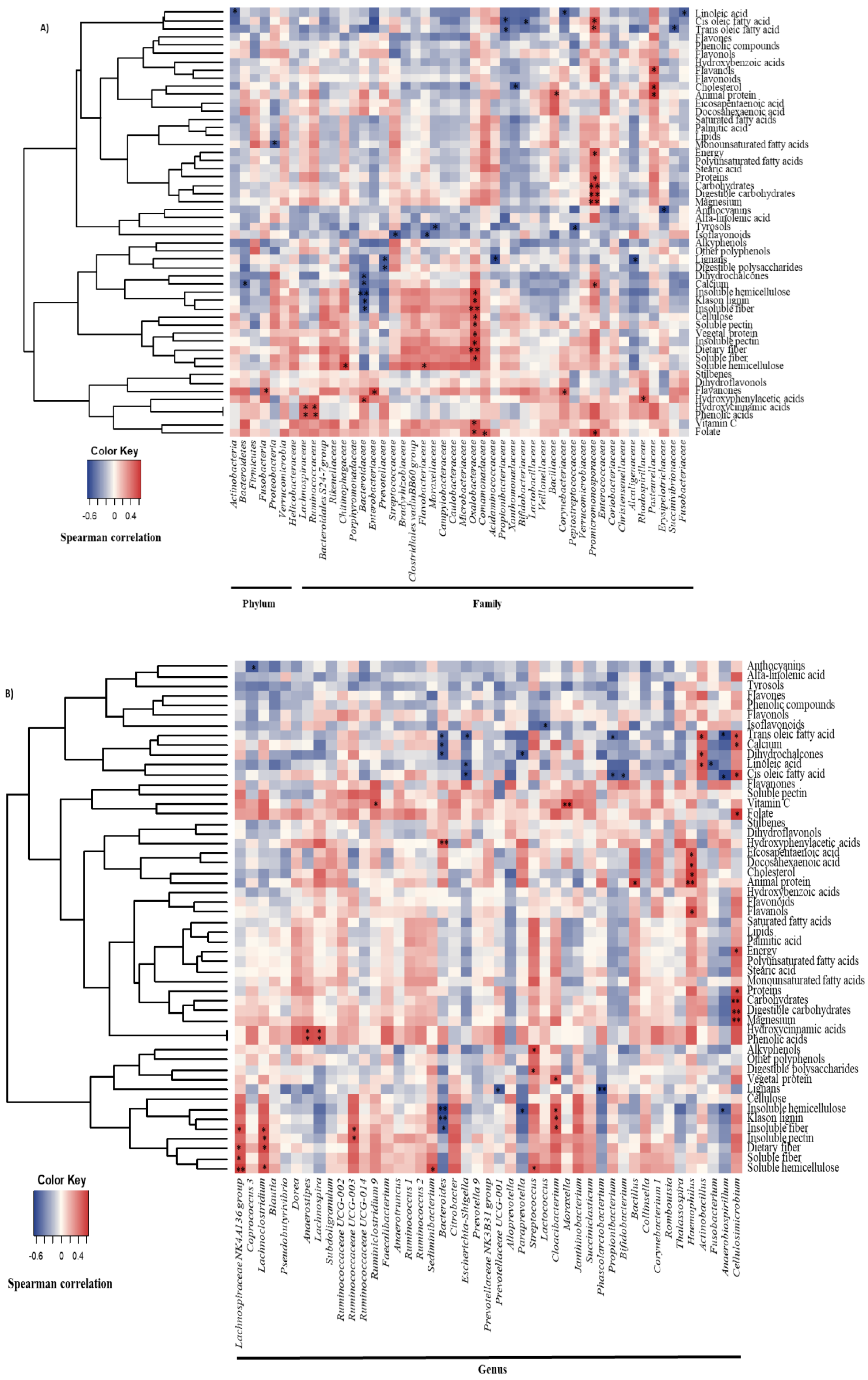{kind=link}
| Variables | Patients with Cholelithiasis (n = 14) | Control Subjects (n = 14) |
|---|---|---|
| Age (year) | 51.50 ± 14.10 | 46.50 ± 11.47 |
| Female (%) | 78.6 | 78.6 |
| BMI (kg/m2) | 26.38 ± 6.04 | 26.00 ± 3.92 |
| Drug use (%) | 50.0 | 28.6 |
| Probiotics consumption (%) | 14.3 | 35.7 |
| Vitamin and mineral supplements consumption (%) | 0.0 | 35.7 ** |
| Fiber supplementation (%) | 0.0 | 14.3 |
| Ethanol (g/day) | 3.98 ± 4.98 | 5.64 ± 6.70 |
| Current smoker (%) | 14.3 | 14.3 |
| Sedentary lifestyle (%) | 50.0 | 50.0 |
| Serum glucose (mg/dL) | 79.14 ± 17.65 | 91.00 ± 9.83 * |
| Serum triglycerides (mg/dL) | 129.14 ± 84.53 | 77.00 ± 33.63 * |
| HDL (mg/dL) | 49.93 ± 14.79 | 61.21 ± 11.18 * |
| LDL (mg/dL) | 117.07 ± 37.78 | 129.79 ± 35.22 |
| Serum total cholesterol (mg/dL) | 192.79 ± 55.33 | 206.71 ± 39.47 |
| Dietary Habits | Patients with Cholelithiasis (n = 14) | Control Subjects (n = 14) |
|---|---|---|
| Appetite a (%) | ||
| Excellent‒Good | 57.2 | - |
| Normal | 28.5 | - |
| Regular | 14.3 | - |
| Chewing problems a (%) | 0.0 | - |
| Special diet a (%) | 35.7 | - |
| Omit foodstuffs from diet a (%) | 64.3 | - |
| Legumes | 35.7 | - |
| Dairy products | 14.3 | - |
| Red meat and meat products | 64.3 | - |
| Vegetables | 14.3 | - |
| No. of subjects reporting a change in food habits a (%) | 28.6 | - |
| Food Groups (g/day) | ||
| Cereals | 121.14 ± 74.97 | 142.45 ± 68.39 |
| Dairy products | 415.86 ± 206.42 | 522.15 ± 227.64 |
| Eggs | 13.78 ± 13.38 | 18.60 ± 11.13 |
| Fats and oils | 23.30 ± 12.44 | 31.16 ± 16.99 |
| Sugar and sugary products | 16.74 ± 20.78 | 17.71 ± 17.45 |
| Vegetables | 200.17 ± 85.11 | 318.16 ± 147.14 * |
| Potatoes | 56.46 ± 30.51 | 33.27 ± 23.51 * |
| Legumes | 27.10 ± 20.82 | 32.60 ± 26.65 |
| Fruits | 244.90 ± 169.37 | 320.66 ± 151.02 |
| Meat and derived products | 129.44 ± 63.15 | 109.76 ± 34.00 |
| Fish and derived products | 60.49 ± 26.76 | 65.43 ± 31.55 |
| Seafood | 12.53 ± 14.30 | 20.90 ± 24.76 |
| Non-alcoholic drinks | 96.94 ± 142.15 | 369.62 ± 235.93 ** |
| Alcoholic drinks | 86.22 ± 110.36 | 134.96 ± 167.97 |
| Nuts and seed | 20.52 ± 55.29 | 10.72 ± 14.83 |
| Sauces | 2.82 ± 4.03 | 8.29 ± 7.82 * |
| Snack | 2.86 ± 10.69 | 2.60 ± 4.39 |
| Unadjusted | Adjusted | |||
|---|---|---|---|---|
| Dietary Components | Patients with Cholelithiasis (n = 14) | Control Subjects (n = 14) | Patients with Cholelithiasis (n = 14) | Control Subjects (n = 14) |
| Energy (kcal/day) | 1660.25 ± 659.28 | 2079.06 ± 515.84 * | - | - |
| Carbohydrates (g/day) | 170.16 ± 72.05 | 209.71 ± 51.60 | 190.12 ± 72.05 | 189.74 ± 51.60 |
| Digestible carbohydrates | 82.04 ± 41.35 | 114.03 ± 30.63 * | 91.77 ± 41.35 | 104.30 ± 30.63 |
| Digestible polysaccharides | 80.46 ± 38.54 | 84.29 ± 37.14 | 91.39 ± 38.54 | 73.36 ± 37.14 |
| Proteins (g/day) | 87.20 ± 21.78 | 100.88 ± 19.97 | 93.52 ± 21.78 | 94.56 ± 21.78 |
| Animal | 58.22 ± 16.68 | 66.05 ± 17.71 | 60.61 ± 16.68 | 63.63 ± 17.71 |
| Vegetable | 25.02 ± 16.46 | 31.78 ± 18.09 | 29.44 ± 16.46 | 27.37 ± 18.09 |
| Lipids (g/day) | 66.65 ± 36.77 | 88.06 ± 34.30 * | 78.31 ± 36.77 | 76.39 ± 34.30 |
| SFA | 18.23 ± 9.18 | 25.78 ± 10.05 | 21.04 ± 9.18 | 22.98 ± 10.05 |
| Palmitic fatty acid | 8.60 ± 4.05 | 10.73 ± 4.37 | 9.82 ± 4.05 | 9.51 ± 4.37 |
| Stearic fatty acid | 3.46 ± 2.57 | 3.82 ± 1.55 | 4.11 ± 2.57 | 3.17 ± 1.55 * |
| MUFA | 29.38 ± 13.27 | 39.10 ± 16.59 | 33.56 ± 13.27 | 34.92 ± 16.59 |
| Cis oleic fatty acid | 1.28 ± 1.01 | 1.55 ± 1.07 | 1.43 ± 1.01 | 1.41 ± 1.07 |
| Trans oleic fatty acid | 0.12 ± 0.14 | 0.14 ± 0.14 | 0.13 ± 0.14 | 0.13 ± 0.14 |
| PUFA | 13.52 ± 18.90 | 15.19 ± 10.25 * | 17.54 ± 18.90 | 11.18 ± 10.25 |
| Linoleic fatty acid | 0.19 ± 0.11 | 0.63 ± 0.95 | 0.20 ± 0.11 | 0.63 ± 0.95 |
| Alfa-linolenic fatty acid | 0.09 ± 0.08 | 0.11 ± 0.07 | 0.10 ± 0.08 | 0.10 ± 0.07 |
| Docosahexaenoic fatty acid | 0.21 ± 0.13 | 0.23 ± 0.14 | 0.20 ± 0.13 | 0.23 ± 0.14 |
| Eicosapentaenoic fatty acid | 0.09 ± 0.05 | 0.11 ± 0.08 | 0.09 ± 0.05 | 0.11 ± 0.08 |
| Folate (µg/day) | 301.26 ± 148.29 | 446.34 ± 111.00 ** | 330.25 ± 148.29 | 417.36 ± 111.00 * |
| Cholesterol (mg/day) | 243.47 ± 126.33 | 303.03 ± 86.00 | 255.53 ± 126.33 | 290.97 ± 86.00 |
| Calcium (mg/day) | 840.95 ± 278.81 | 1260.04 ± 307.79 ** | 916.85 ± 278.81 | 1184.14 ± 307.79 ** |
| Magnesium (mg/day) | 348.44 ± 270.31 | 403.52 ± 110.17 * | 403.30 ± 270.31 | 348.66 ± 110.17 |
| Vitamin C (mg/day) | 111.50 ± 61.73 | 193.63 ± 66.52 ** | 109.75 ± 61.73 | 195.39 ± 66.52 ** |
| Phenolic compounds (mg/day) | 946.11 ± 473.35 | 1780.48 ± 899.58 ** | 1025.80 ± 473.35 | 1700.78 ± 899.58 * |
| Flavonoids | 211.79 ± 141.50 | 499.54 ± 393.48 * | 248.44 ± 141.50 | 462.89 ± 393.48 |
| Anthocyanins | 7.45 ± 10.69 | 26.07 ± 35.29 * | 5.49 ± 10.69 | 28.03 ± 35.29 * |
| Dihydroflavonols | 1.04 ± 1.78 | 1.33 ± 2.84 | 1.00 ± 1.78 | 1.38 ± 2.84 |
| Dihydrochalcones | 1.78 ± 1.98 | 2.62 ± 3.97 | 1.94 ± 1.98 | 2.46 ± 3.97 |
| Flavanols | 167.01 ± 122.61 | 360.71 ± 397.60 | 206.56 ± 122.61 | 321.16 ± 397.60 |
| Flavanones | 16.87 ± 33.49 | 43.54 ± 31.14 ** | 13.55 ± 33.49 | 46.86 ± 31.14 * |
| Flavones | 0.54 ± 0.57 | 5.15 ± 4.15 ** | 0.20 ± 0.57 | 5.49 ± 4.15 ** |
| Flavonols | 17.06 ± 10.18 | 37.27 ± 18.16 ** | 16.71 ± 10.18 | 37.63 ± 18.16 * |
| Isoflavonoids | 0.03 ± 0.03 | 22.84 ± 76.64 | 3.00 ± 0.03 | 19.88 ± 76.64 |
| Lignans | 0.40 ± 0.61 | 1.14 ± 0.60 ** | 0.49 ± 0.61 | 1.05 ± 0.60 * |
| Phenolic acids | 122.64 ± 153.59 | 336.09 ± 301.89 ** | 145.93 ± 153.59 | 312.80 ± 301.89 |
| Hydroxybenzoic acids | 7.20 ± 9.24 | 40.61 ± 28.38 ** | 7.56 ± 9.24 | 40.25 ± 28.38 ** |
| Hydroxycinnamic acids | 115.37 ± 152.11 | 294.28 ± 292.73 ** | 138.21 ± 152.11 | 271.44 ± 292.73 |
| Hydroxyphenylacetic acids | 0.07 ± 0.09 | 0.98 ± 1.61 ** | 0.14 ± 0.09 | 0.91 ± 1.61 |
| Stilbenes | 0.68 ± 1.13 | 0.99 ± 1.85 | 0.64 ± 1.13 | 1.04 ± 1.85 |
| Other polyphenols | 9.61 ± 12.99 | 42.22 ± 32.60 ** | 10.65 ± 12.99 | 41.18 ± 32.60 ** |
| Alkyphenols | 4.75 ± 8.53 | 18.92 ± 29.75 | 4.56 ± 8.53 | 19.11 ± 29.75 |
| Tyrosols | 4.05 ± 6.41 | 19.34 ± 10.83 ** | 5.29 ± 6.41 | 18.11 ± 10.83 ** |
| Dietary fiber (g/day) | 20.20 ± 8.47 | 24.21 ± 9.87 | 22.05 ± 8.47 | 22.36 ± 9.87 |
| Soluble | 2.28 ± 1.18 | 2.56 ± 0.94 | 2.42 ± 1.18 | 2.41 ± 0.94 |
| Pectin | 0.74 ± 0.49 | 0.90 ± 0.43 | 0.74 ± 0.49 | 0.90 ± 0.43 |
| Hemicellulose | 1.43 ± 0.87 | 1.51 ± 0.66 | 1.57 ± 0.87 | 1.36 ± 0.66 |
| Insoluble | 12.77 ± 6.25 | 15.60 ± 7.40 | 13.44 ± 6.25 | 14.92 ± 7.40 |
| Pectin | 1.65 ± 0.84 | 1.84 ± 0.77 | 1.71 ± 0.84 | 1.77 ± 0.77 |
| Hemicellulose | 4.61 ± 3.04 | 5.13 ± 3.01 | 4.99 ± 3.04 | 4.75 ± 3.01 |
| Klason lignin | 1.68 ± 1.13 | 2.18 ± 0.85 | 1.84 ± 1.13 | 2.02 ± 0.85 |
| Cellulose | 5.10 ± 2.03 | 6.26 ± 3.08 | 5.31 ± 2.03 | 6.03 ± 3.08 |
| Predictors | R2 | β | p Value | |
|---|---|---|---|---|
| Phylum | ||||
| Actinobacteria | Linoleic fatty acid (g/day) | 0.360 | −0.600 | 0.023 |
| Family | ||||
| Bacteroidaceae | Hydroxyphenylacetic acids (mg/day) | 0.709 | 0.722 | 0.001 |
| Insoluble fiber (g/day) | −0.401 | 0.031 | ||
| Chitinophagaceae | Soluble hemicellulose (g/day) | 0.524 | 0.724 | 0.003 |
| Flavobacteriaceae | Insoluble hemicellulose (g/day) | 0.393 | 0.627 | 0.016 |
| Lachnospiraceae | Hydroxycinnamic acids (mg/day) | 0.383 | 0.619 | 0.018 |
| Oxalobacteriaceae | Insoluble pectin (g/day) | 0.728 | 0.853 | 0.000 |
| Promicromonosporaceae | Magnesium (mg/day) | 0.962 | 1.116 | 0.000 |
| Digestible carbohydrates (g/day) | −0.313 | 0.002 | ||
| Propionibacteriaceae | Trans oleic fatty acid (g/day) | 0.378 | −0.615 | 0.019 |
| Genus | ||||
| Actinobacillus | Trans oleic fatty acid (g/day) | 0.321 | 0.566 | 0.035 |
| Bacteroides | Hydroxyphenylacetic acids (mg/day) | 0.709 | 0.722 | 0.001 |
| Insoluble fiber (g/day) | −0.401 | 0.031 | ||
| Cellulosimicrobium | Magnesium (mg/day) | 0.962 | 1.161 | 0.000 |
| Digestible carbohydrates (g/day) | −0.313 | 0.002 | ||
| Cloacibacterium | Insoluble fiber (g/day) | 0.549 | 0.741 | 0.002 |
| Escherichia-Shigella | Linoleic acid (g/day) | 0.286 | −0.535 | 0.049 |
| Lachnosclostridium | Dietary fiber (g/day) | 0.316 | 0.562 | 0.036 |
| Lachnospira | Hydroxycinnamic acids (mg/day) | 0.894 | 0.946 | 0.000 |
| Moraxella | Vitamin C (mg/day) | 0.652 | 0.807 | 0.000 |
| Propionibacterium | Trans oleic fatty acid (g/day) | 0.379 | −0.616 | 0.019 |
| Ruminoclostridium 9 | Vitamin C (mg/day) | 0.430 | 0.656 | 0.011 |
| Sediminibacterium | Soluble hemicellulose (g/day) | 0.533 | 0.730 | 0.003 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Díaz, I.; Molinero, N.; Cabrera, A.; Rodríguez, J.I.; Margolles, A.; Delgado, S.; González, S. Diet: Cause or Consequence of the Microbial Profile of Cholelithiasis Disease? Nutrients 2018, 10, 1307. https://doi.org/10.3390/nu10091307
Gutiérrez-Díaz I, Molinero N, Cabrera A, Rodríguez JI, Margolles A, Delgado S, González S. Diet: Cause or Consequence of the Microbial Profile of Cholelithiasis Disease? Nutrients. 2018; 10(9):1307. https://doi.org/10.3390/nu10091307
Chicago/Turabian StyleGutiérrez-Díaz, Isabel, Natalia Molinero, Ana Cabrera, José Ignacio Rodríguez, Abelardo Margolles, Susana Delgado, and Sonia González. 2018. "Diet: Cause or Consequence of the Microbial Profile of Cholelithiasis Disease?" Nutrients 10, no. 9: 1307. https://doi.org/10.3390/nu10091307