Mouse Chow Composition Influences Immune Responses and Food Allergy Development in a Mouse Model

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Immunization with OVA
2.3. Oral Challenge with OVA
2.4. Measurement of OVA-Specific Antibody Levels
2.5. Splenocyte Stimulation and Cytokine Read-Out
2.6. Statistic and Data Analysis
3. Results
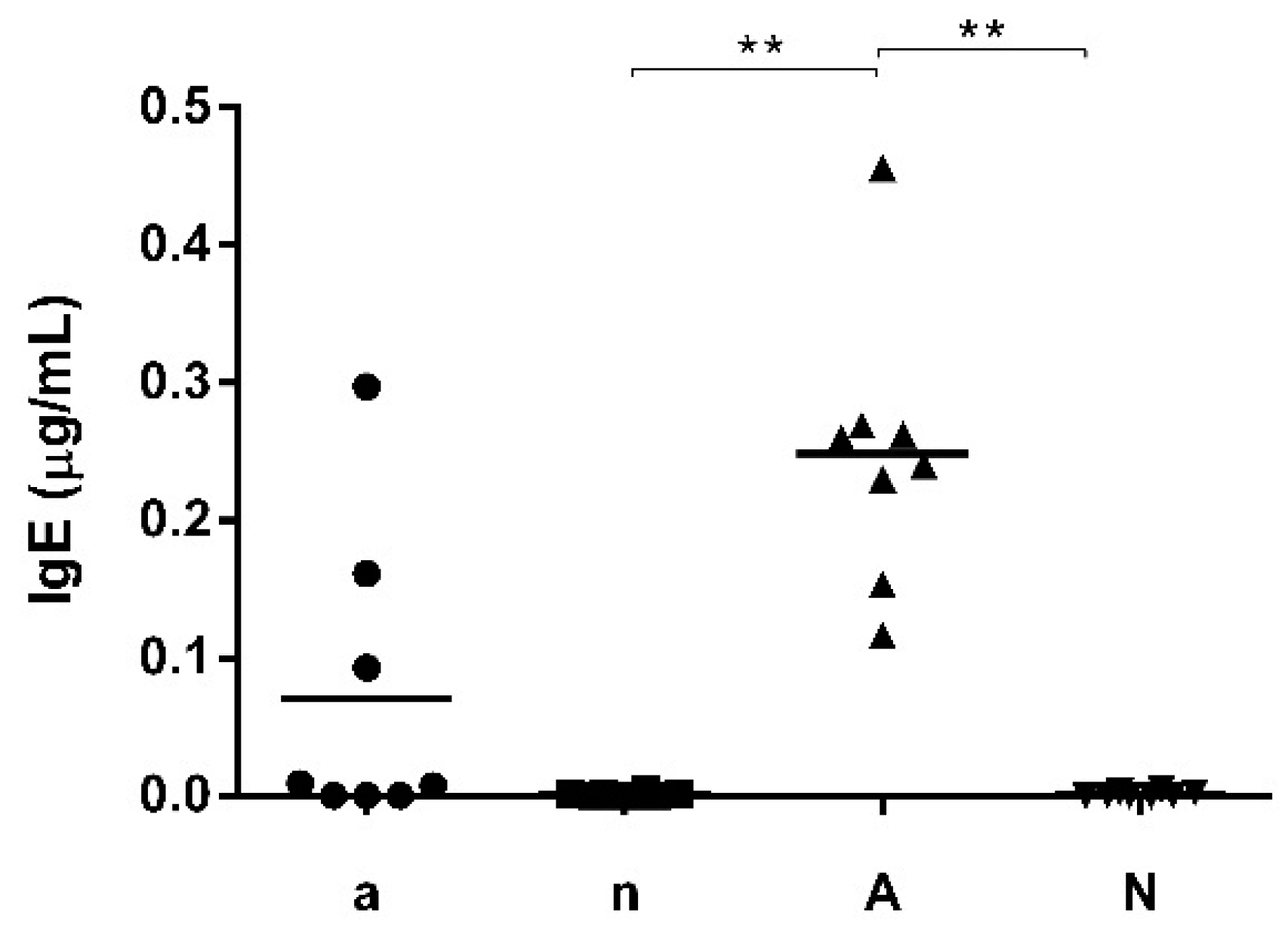
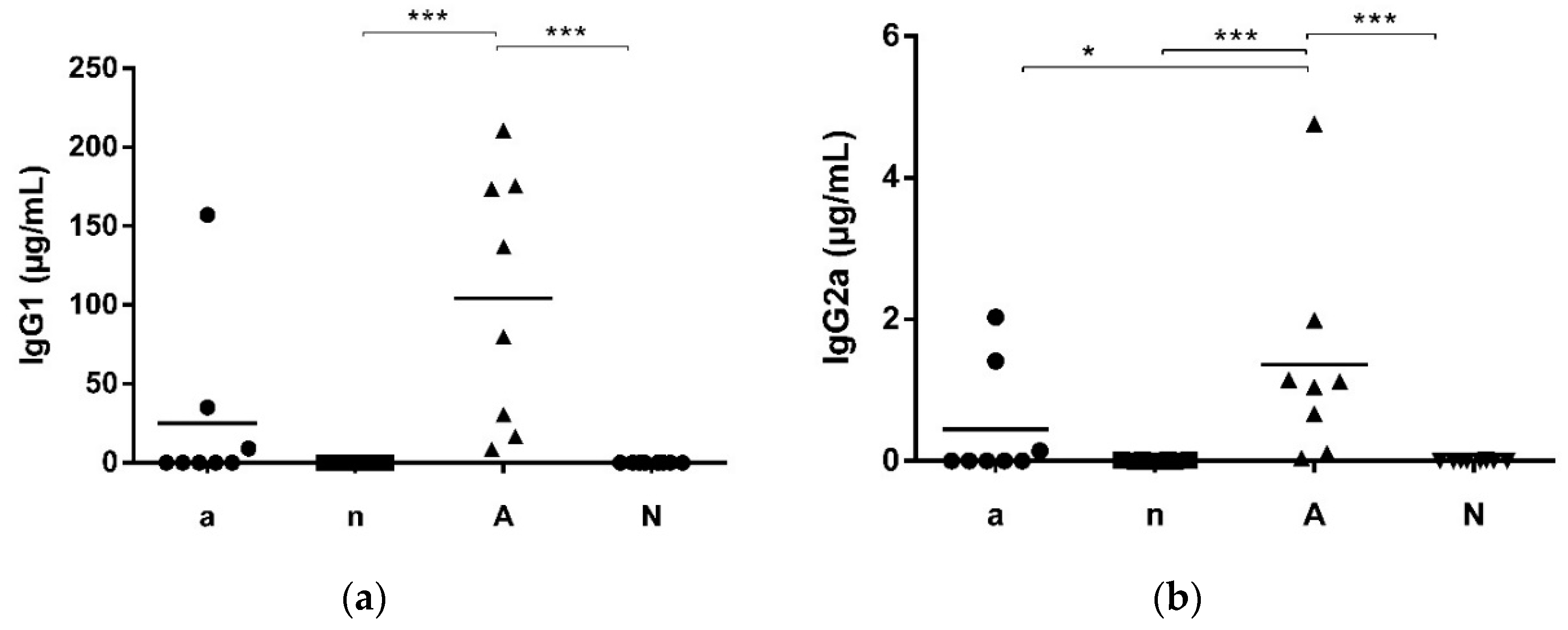
3.1. Mouse Feed Influences Systemic and Local Intestinal Antibody Production
3.2. Cytokine Production Reveal a Tolerogenic Milieu Based in Mouse Feed
3.3. Temperature Changes after Oral OVA Challenge Indicate Food Allergy Development
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Julia, V.; Macia, L.; Dombrowicz, D. The impact of diet on asthma and allergic diseases. Nat. Rev. Immunol. 2015, 15, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, L.M.; Woodward, B. A comparison of the capacity of six cold-pressed plant oils to support development of acquired immune competence in the weanling mouse: Superiority of low-linoleic-acid oils. Br. J. Nutr. 2002, 88, 171. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, J.S.; Hayek, M.G.; Reinhart, G.A.; Chew, B.P. Dietary fish oil and flaxseed oil suppress inflammation and immunity in cats. Vet. Immunol. Immunopathol. 2011, 141, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Biological significance of essential fatty acids. J. Assoc. Physicians India 2006, 54, 309–319. [Google Scholar] [PubMed]
- Hogenkamp, A.; van Vlies, N.; Fear, A.L.; van Esch, B.C.; Hofman, G.A.; Garssen, J.; Calder, P.C. Dietary fatty acids affect the immune system in male mice sensitized to ovalbumin or vaccinated with influenza. J. Nutr. 2011, 141, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, D.W.L.; Kang, J.X.; Kulka, M. N-3 polyunsaturated fatty acids inhibit Fc ε receptor I-mediated mast cell activation. J. Nutr. Biochem. 2015, 26, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Arita, M. Mediator lipidomics in acute inflammation and resolution. J. Biochem. 2012, 152, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunisawa, J.; Arita, M.; Hayasaka, T.; Harada, T.; Iwamoto, R.; Nagasawa, R.; Shikata, S.; Nagatake, T.; Suzuki, H.; Hashimoto, E.; et al. Dietary ω3 fatty acid exerts anti-allergic effect through the conversion to 17,18-epoxyeicosatetraenoic acid in the gut. Sci. Rep. 2015, 5, 9750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, T.; Arita, M.; Isobe, Y.; Iwamoto, R.; Goto, T.; Yoshioka, T.; Urabe, D.; Inoue, M.; Arai, H. Eicosapentaenoic acid is converted via ω-3 epoxygenation to the anti-inflammatory metabolite 12-hydroxy-17,18-epoxyeicosatetraenoic acid. FASEB J. 2014, 28, 586–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemsen, L.E.M. Dietary n-3 long chain polyunsaturated fatty acids in allergy prevention and asthma treatment. Eur. J. Pharmacol. 2016, 785, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Thang, C.L.; Boye, J.I.; Shi, H.N.; Zhao, X. Effects of supplementing different ratios of omega-3 and omega-6 fatty acids in western-style diets on cow’s milk protein allergy in a mouse model. Mol. Nutr. Food Res. 2013, 57, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.S.; Wang, J.; Zhang, X.; Zhang, L.; Zhang, H.P.; Wang, L.; Wood, L.G.; Wang, G. Is the consumption of fast foods associated with asthma or other allergic diseases? Respirology 2018. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.; Song, Y.; Brown, H.; Hart, P.H.; Zhang, G.B. Cellular and molecular mechanisms of vitamin D in food allergy. J. Cell. Mol. Med. 2018, 22, 3270–3277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, J.H.; Shin, Y.H.; Chung, I.H.; Kim, H.J.; Yoo, E.-G.; Yoon, J.W.; Jee, H.M.; Chang, Y.E.; Han, M.Y. The link between serum vitamin D. level, sensitization to food allergens and the severity of atopic dermatitis in infancy. J. Pediatr. 2014, 165, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhong, Y.; Shen, X.; Yang, K.; Cai, W. Maternal and early-life vitamin D deficiency enhances allergic reaction in an ovalbumin-sensitized BALB/c mouse model. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.J.; Koplin, J.J.; Ponsonby, A.-L.; Gurrin, L.C.; Wake, M.; Vuillermin, P.; Martin, P.; Matheson, M.; Lowe, A.; Robinson, M.; et al. Vitamin D insufficiency is associated with challenge-proven food allergy in infants. J. Allergy Clin. Immunol. 2013, 131, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Norizoe, C.; Akiyama, N.; Segawa, T.; Tachimoto, H.; Mezawa, H.; Ida, H.; Urashima, M. Increased food allergy and vitamin D: Randomized, double-blind, placebo-controlled trial. Pediatr. Int. 2014, 56, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwaru, B.I.; Hadkhale, K.; Hämäläinen, N.; Takkinen, H.-M.; Ahonen, S.; Ilonen, J.; Toppari, J.; Niemelä, O.; Haapala, A.-M.; Veijola, R.; et al. Vitamin D intake during the first 4 years and onset of asthma by age 5: A nested case-control study. Pediatr. Allergy Immunol. 2017, 28, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Weisse, K.; Winkler, S.; Hirche, F.; Herberth, G.; Hinz, D.; Bauer, M.; Röder, S.; Rolle-Kampczyk, U.; von Bergen, M.; Olek, S.; et al. Maternal and newborn vitamin D status and its impact on food allergy development in the German LINA cohort study. Allergy 2013, 68, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Hyppönen, E.; Berry, D.J.; Wjst, M.; Power, C. Serum 25-hydroxyvitamin D and IgE—A significant but nonlinear relationship. Allergy 2009, 64, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Rothers, J.; Wright, A.L.; Stern, D.A.; Halonen, M.; Camargo, C.A., Jr. Cord blood 25-hydroxyvitamin D levels are associated with aeroallergen sensitization in children from Tucson, Arizona. J. Allergy Clin. Immunol. 2011, 128, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Barcik, W.; Untersmayr, E.; Pali-Schöll, I.; O’Mahony, L.; Frei, R. Influence of microbiome and diet on immune responses in food allergy models. Drug. Discov. Today. Dis. Models 2015, 17, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.; Ah-Leung, S.; Drumare, M.-F.; Feraudet-Tarisse, C.; Verhasselt, V.; Wal, J.-M.; Créminon, C.; Adel-Patient, K. Peanut allergens are rapidly transferred in human breast milk and can prevent sensitization in mice. Allergy 2014, 69, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Knippels, L.M.; Penninks, A.H.; Houben, G.F. Continued expression of anti-soy protein antibodies in rats bred on a soy protein–free diet for one generation: The importance of dietary control in oral sensitization research. J. Allergy Clin. Immunol. 1998, 101, 815–820. [Google Scholar] [CrossRef]
- Diesner, S.C.; Knittelfelder, R.; Krishnamurthy, D.; Pali-Schöll, I.; Gajdzik, L.; Jensen-Jarolim, E.; Untersmayr, E. Dose-dependent food allergy induction against ovalbumin under acid-suppression: A murine food allergy model. Immunol. Lett. 2008. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://www.graphpad.com (accessed on 11 June 2018).
- Tordesillas, L.; Berin, M.C. Mechanisms of oral tolerance. Clin. Rev. Allergy Immunol. 2018, 55, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Myles, I.A. Fast food fever: Reviewing the impacts of the Western diet on immunity. Nutr. J. 2014, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Gibson, P.G. Dietary factors lead to innate immune activation in asthma. Pharmacol. Ther. 2009, 123, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Lawson, J.A.; Rennie, D.C.; Dosman, J.A.; Cammer, A.L.; Senthilselvan, A. Obesity, diet and activity in relation to asthma and wheeze among rural dwelling children and adolescents. J. Obes. 2013, 2013, 315096. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Garg, M.L.; Gibson, P.G. A high-fat challenge increases airway inflammation and impairs bronchodilator recovery in asthma. J. Allergy Clin. Immunol. 2011, 127, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Skypala, I.J.; McKenzie, R. Nutritional issues in food allergy. Clin. Rev. Allergy Immunol. 2018, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.M.; Friedman, J.E. Maternal modifiers of the infant gut microbiota: Metabolic consequences. J. Endocrinol. 2017, 235, R1–R12. [Google Scholar] [CrossRef] [PubMed]
- Kehl-Fie, T.E.; Skaar, E.P. Nutritional immunity beyond iron: A role for manganese and zinc. Curr. Opin. Chem. Biol. 2010, 14, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Damo, S.M.; Kehl-Fie, T.E.; Sugitani, N.; Holt, M.E.; Rathi, S.; Murphy, W.J.; Zhang, Y.; Betz, C.; Hench, L.; Fritz, G.; et al. Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proc. Natl. Acad. Sci. USA 2013, 110, 3841–3846. [Google Scholar] [CrossRef] [PubMed]
- Diesner, S.C.; Bergmayr, C.; Pfitzner, B.; Assmann, V.; Krishnamurthy, D.; Starkl, P.; Endesfelder, D.; Rothballer, M.; Welzl, G.; Rattei, T.; et al. A distinct microbiota composition is associated with protection from food allergy in an oral mouse immunization model. Clin. Immunol. 2016, 173, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwaru, B.I.; Erkkola, M.; Lumia, M.; Kronberg-Kippilä, C.; Ahonen, S.; Kaila, M.; Ilonen, J.; Simell, O.; Knip, M.; Veijola, R.; et al. Maternal intake of fatty acids during pregnancy and allergies in the offspring. Br. J. Nutr. 2012, 108, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. N-3 Polyunsaturated fatty acids, inflammation and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, S1505–S1519. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Calder, P.C. Can early omega-3 fatty acid exposure reduce risk of childhood allergic disease? Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Mirzakhani, H.; Al-Garawi, A.; Weiss, S.T.; Litonjua, A.A. Vitamin D and the development of allergic disease: How important is it? Clin. Exp. Allergy 2015, 45, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Fleet, J.C.; Gliniak, C.; Zhang, Z.; Xue, Y.; Smith, K.B.; McCreedy, R.; Adedokun, S.A. Serum metabolite profiles and target tissue gene expression define the effect of cholecalciferol intake on calcium metabolism in rats and mice. J. Nutr. 2008, 138, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Laboratory Animals: Fourth Revised Edition; The National Academies Press: Washington, DC, USA, 1995. [Google Scholar]
- Cantorna, M.T.; Snyder, L.; Lin, Y.-D.; Yang, L.; Vitamin, D. and 1,25(OH)2D regulation of T cells. Nutrients 2015, 7, 3011–3021. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.; Luo, W.; Sui, Y.; Li, Z.; Hua, J. Dietary n-3 PUFA protects mice from con a induced liver injury by modulating regulatory T cells and ppar-γ expression. PLoS ONE 2015, 10, e0132741. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.M.; Song, Y.; Li, X.-M.; Sampson, H.A.; Masilamani, M. Dietary elimination of soybean components enhances allergic immune response to peanuts in BALB/c mice. Int. Arch. Allergy Immunol. 2015, 166, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Masilamani, M.; Wei, J.; Sampson, H.A. Regulation of the immune response by soybean isoflavones. Immunol. Res. 2012, 54, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Masilamani, M.; Wei, J.; Bhatt, S.; Paul, M.; Yakir, S.; Sampson, H.A. Soybean isoflavones regulate dendritic cell function and suppress allergic sensitization to peanut. J. Allergy Clin. Immunol. 2011, 128, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Jarolim, E.; Untersmayr, E. Gender-medicine aspects in allergology. Allergy 2008, 63, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, F.H. 90th Anniversary commentary: The AIN-93 purified diets for laboratory rodents—The development of a landmark article in the journal of nutrition and its impact on health and disease research using rodent models. J. Nutr. 2018, 148, 1667–1670. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soy-Containing Feed (Ssniff Rat/Mouse Maintenance Fortified V1534-300) | Soy-Free Feed (Lasvendi LasQCdiet Rod16-A) |
|---|---|
| Dietary Ingredients (in Descending Amount) | Dietary Ingredients (in Descending Amount) |
| Wheat soybean products barley minerals oat hulls molasses vitamins & trace elements L-lysine HCl DL-methionine | Wheat corn gluten oats corn barley linseed oil molasses brewer’s yeast minerals vitamins/trace elements-mix |
| Energy per kg | Energy per kg |
| Gross Energy (GE) 16.2 MJ Metabolizable Energy (ME) 13.5 MJ | Gross Energy (GE) 15.9 MJ Metabolizable Energy (ME) 13.1 MJ calc. formula §14 FMVO |
| Minerals [%] | Minerals [%] |
| Calcium 1.0 Phosphorus 0.7 Sodium 0.24 Magnesium 0.22 Potassium 0.92 Ca/P = 1.43: 1 | Calcium 1.0 Phosphorus 0.65 Sodium 0.3 Magnesium 0.25 |
| Amino acids [%] | Amino acids [%] |
| Arginine 1.19 Cysteine 0.35 Histidine 0.49 Isoleucine 0.79 Leucine 1.39 Lysine 1.1 Methionine 0.38 Phenylalanine 0.89 Threonine 0.72 Tryptophan 0.25 Alanine 0.87 Aspartic acid 1.84 Glutamic acid 4.22 Glycine 0.89 Proline 1.31 Serine 1.01 Valine 0.92 Met+Cys 0.73 Phe+Tyr 1.5 | Arginine 0.8 Cysteine 0.35 Histidine 0.4 Isoleucine 0.65 Leucine 1.7 Lysine 0.9 Methionine 0.45 Phenylalanine 0.85 Threonine 0.6 Tryptophan 0.2 Tyrosine 0.6 |
| Vitamins per kg | Vitamins per kg |
| Vitamin A 25,000 IU Vitamin D3 1500 IU Vitamin E 135 mg Vitamin K (as MNB) 20 mg Thiamine (B1) 86 mg Riboflavin (B2) 32 mg Pyridoxine (B6) 31 mg Cobalamin (B12) 150 μg Biotin 710 μg Choline 1370 mg Folate 10 mg Pantothenate 59 mg Niacin 153 mg | Vitamin A 25,000 IU Vitamin D3 1800 IU Vitamin E 120 mg Vitamin K 80 mg Thiamine (B1) 100 mg Riboflavin (B2) 30 mg Pyridoxine (B6) 25 mg Cobalamin (B12) 120 mg Biotin 400 µg Choline 1500 mg Folate 6 mg Pantothenate 35 mg Niacin 80 mg |
| Trace Elements per kg | Trace Elements per kg |
| Copper 15 mg Iodine 2.1 mg Iron 186 mg Manganese 68 mg Selenium 0.3 mg Zinc 91 mg | Copper 15 mg Iodine 4 mg Iron 200 mg Manganese 120 mg Selenium 0.2 mg Zinc 75 mg Cobalt 1.5 mg |
| Fatty Acids [%] | Fatty Acids [%] |
| C 14:0 0.01 C 16:0 0.45 C 18:0 0.09 C 20:0 0.01 C 16:1 0.01 C 18:1 0.62 C 18:2 1.76 C 18:3 0.23 | C 16:0 0.5 C 18:0 0.2 C 20:0 0.01 C 18:1 0.9 C 18:2 1.9 C 18:3 0.75 |
| Group | Sensitization | Oral Challenge | Number of Mice | Mouse Chow |
|---|---|---|---|---|
| a | 200 µg OVA oral + acid suppression | 2 mg/mL OVA oral | 8 | soy-free feed |
| n | naive | 2 mg/mL OVA oral | 8 | soy-free feed |
| A | 200 µg OVA oral + acid suppression | 2 mg/mL OVA oral | 8 | soy-containing feed |
| N | naive | 2 mg/mL OVA oral | 8 | soy-containing feed |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weidmann, E.; Samadi, N.; Klems, M.; Heiden, D.; Seppova, K.; Ret, D.; Untersmayr, E. Mouse Chow Composition Influences Immune Responses and Food Allergy Development in a Mouse Model. Nutrients 2018, 10, 1775. https://doi.org/10.3390/nu10111775
Weidmann E, Samadi N, Klems M, Heiden D, Seppova K, Ret D, Untersmayr E. Mouse Chow Composition Influences Immune Responses and Food Allergy Development in a Mouse Model. Nutrients. 2018; 10(11):1775. https://doi.org/10.3390/nu10111775
Chicago/Turabian StyleWeidmann, Eleonore, Nazanin Samadi, Martina Klems, Denise Heiden, Klara Seppova, Davide Ret, and Eva Untersmayr. 2018. "Mouse Chow Composition Influences Immune Responses and Food Allergy Development in a Mouse Model" Nutrients 10, no. 11: 1775. https://doi.org/10.3390/nu10111775