
Assessing the Current and Future Potential Distribution of Solanum rostratum Dunal in China Using Multisource Remote Sensing Data and Principal Component Analysis

Abstract
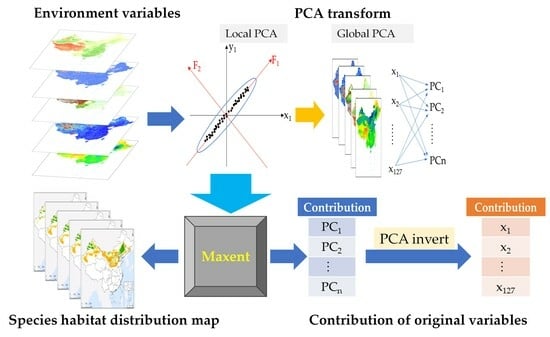
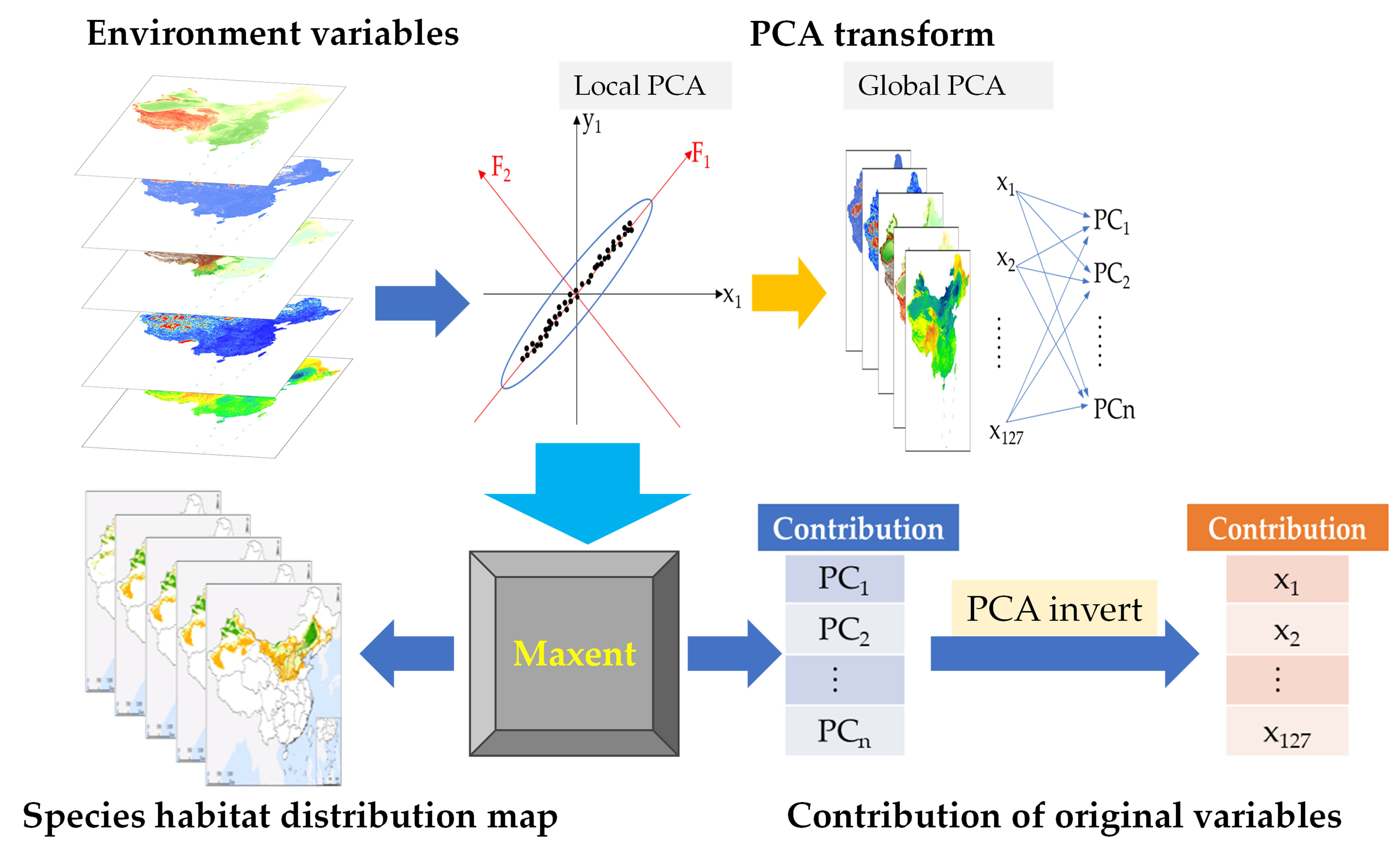
:
1. Introduction
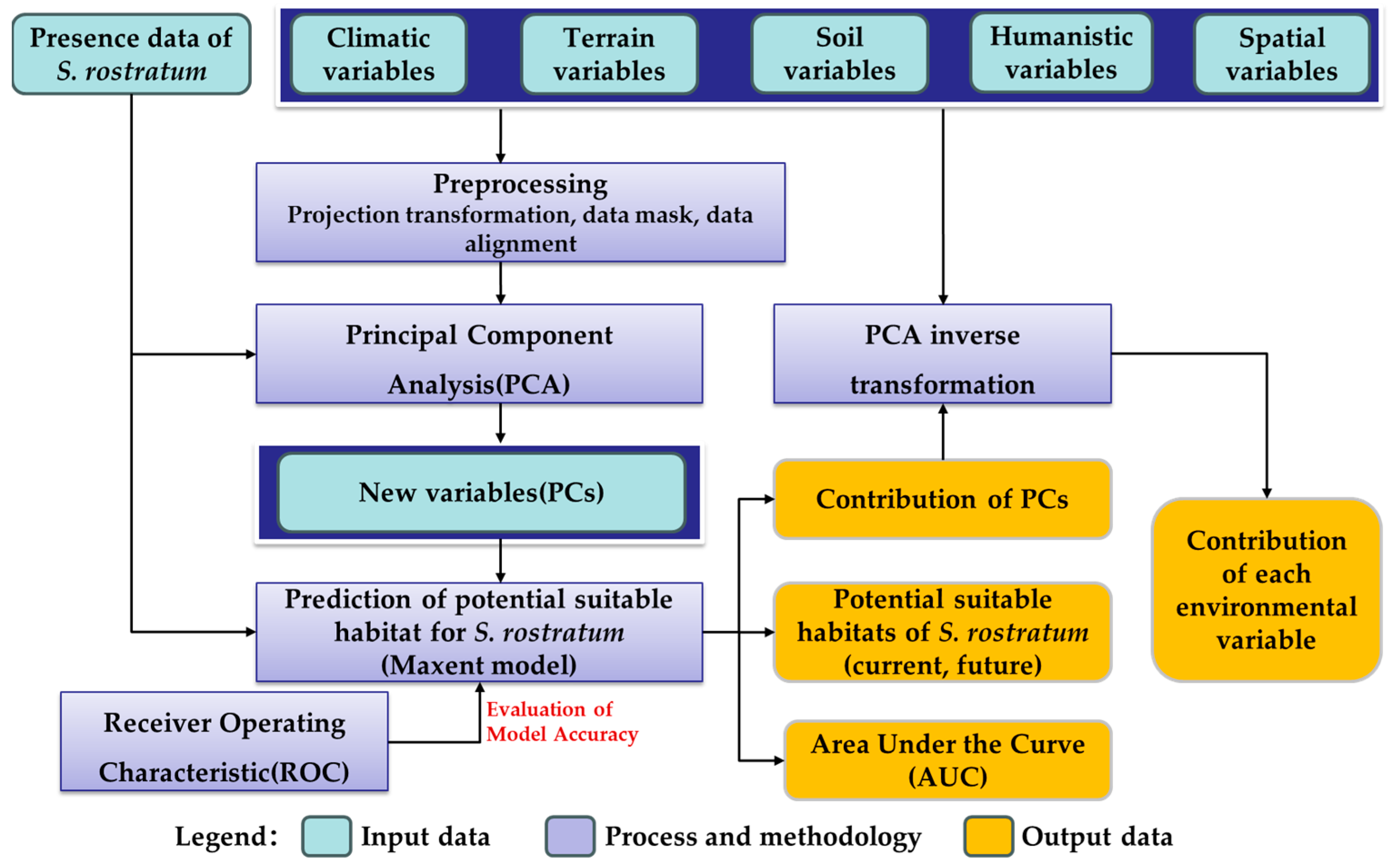
2. Materials and Methods
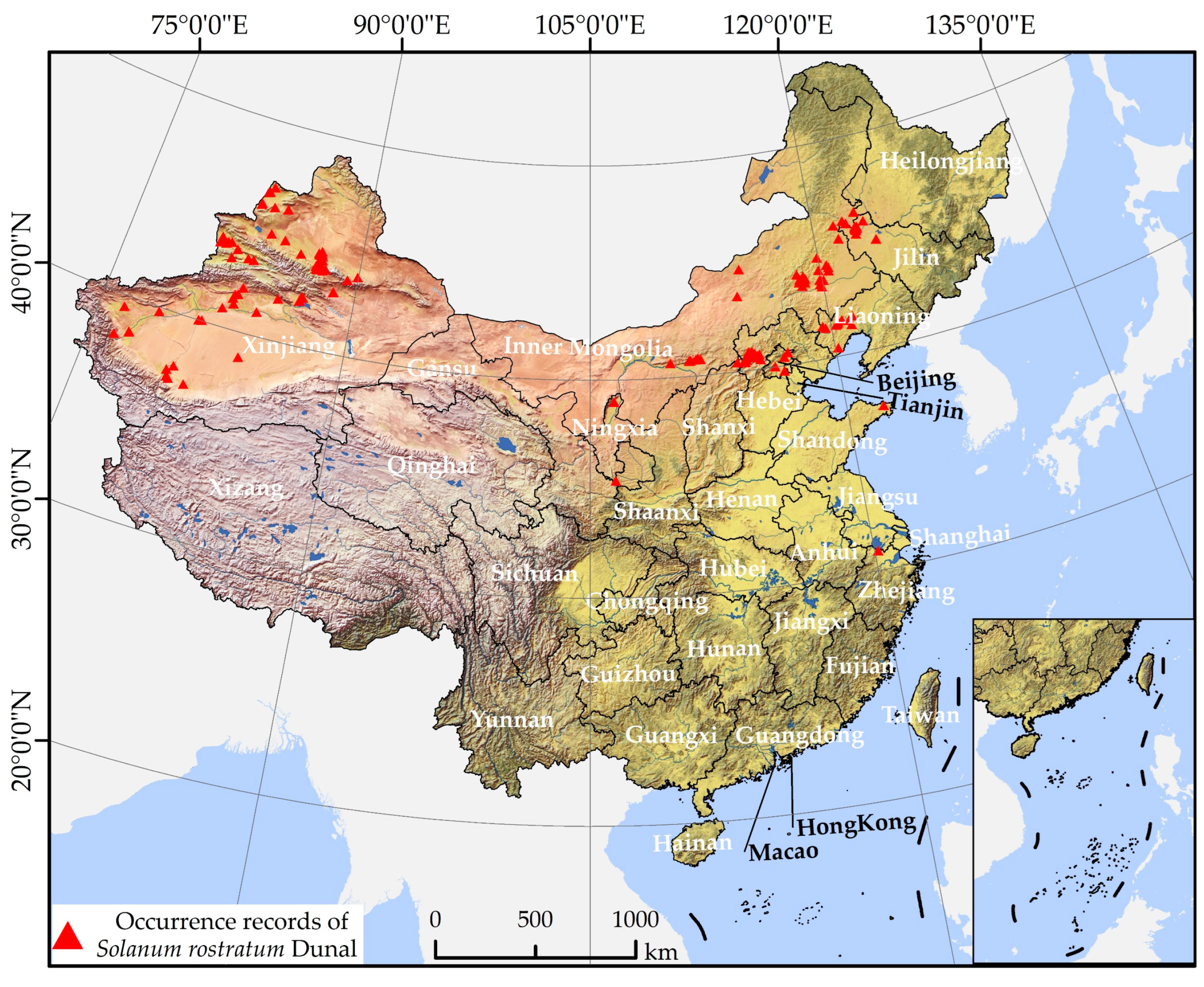
2.1. Presence Data
2.2. Environmental Variables
2.3. Methodology
2.3.1. Principal Component Analysis
2.3.2. Maxent Model
2.3.3. Evaluation of Model Accuracy
2.3.4. Spatial Geometric Center Analysis
3. Results
3.1. Model Accuracy
3.2. The Spatial and Temporal Dynamics of Potential Habitats of S. rostratum
3.2.1. Current Climate Scenarios
3.2.2. Future Climate Scenarios
3.2.3. Temporal and Spatial Dissemination Trends
3.3. Factors Influencing the Suitable Habitat of S. rostratum
3.4. Impact Analysis on Model Prediction Accuracy
3.4.1. Impact of Different Combinations of Environmental Variables
3.4.2. Impact of Decorrelation Methods
4. Discussion
4.1. The Distribution of Potential Suitable Habitats under Contemporary Climate Scenarios
4.2. The Spatial Distribution of S. rostratum Is Influenced by Multiple Factors
4.3. The Future Development of Potential Suitable Habitats for S. rostratum
4.4. The Advantages of Using the PCA Method in Species Distribution Modeling
4.5. Limitations and Suggestions
4.5.1. Limitations
4.5.2. Suggestions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pysek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, B.; Aldridge, D.C.; González Moreno, P.; Pergl, J.; Pizarro, M.; Pyšek, P.; Thuiller, W.; Yesson, C.; Vilà, M. Protected areas offer refuge from invasive species spreading under climate change. Glob. Change Biol. 2017, 23, 5331–5343. [Google Scholar] [CrossRef]
- Kumar Rai, P.; Singh, J.S. Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol. Indic. 2020, 111, 106020. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Sun, S.; Wang, N.; Fan, P.; You, C.; Wang, R.; Zheng, P.; Wang, H. Dynamics of the distribution of invasive alien plants (Asteraceae) in China under climate change. Sci. Total Environ. 2023, 903, 166260. [Google Scholar] [CrossRef]
- Sittaro, F.; Hutengs, C.; Vohland, M. Which factors determine the invasion of plant species? Machine learning based habitat modelling integrating environmental factors and climate scenarios. Int. J. Appl. Earth Obs. Geoinf. 2023, 116, 103158. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Ziska, L.; Dukes, J.S. Invasive Species and Global Climate Change; CABI: Oxfordshire, UK, 2022. [Google Scholar] [CrossRef]
- IPCC. Climate change 2021: The physical science basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2021; Volume 2, Available online: https://www.ipcc.ch/report/ar6/wg1/downloads/report/IPCC_AR6_WGI_FrontMatter.pdf (accessed on 3 August 2023).
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; van Kleunen, M.; Kühn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Glob. Change Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef]
- Guan, G.; Gao, D.; Li, W.; Ye, J.; Xin, X.; Li, S. Solanum rostratum: A quarantine weed. Plant Quar. 1984, 25–28. [Google Scholar] [CrossRef]
- Wu, X.-W.; Luo, J.; Chen, J.-K.; Li, B. Spatial patterns of invasive alien plants in china and its relationship with environmental and anthropological factors. J. Plant Ecol. 2006, 30, 576–584. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Anderson, C.J.; Heins, D.; Pelletier, K.C.; Knight, J.F. Improving Machine Learning Classifications of Phragmites australis Using Object-Based Image Analysis. Remote Sens. 2023, 15, 989. [Google Scholar] [CrossRef]
- Dmitriev, P.A.; Kozlovsky, B.L.; Kupriushkin, D.P.; Dmitrieva, A.A.; Rajput, V.D.; Chokheli, V.A.; Tarik, E.P.; Kapralova, O.A.; Tokhtar, V.K.; Minkina, T.M.; et al. Assessment of Invasive and Weed Species by Hyperspectral Imagery in Agrocenoses Ecosystem. Remote Sens. 2022, 14, 2442. [Google Scholar] [CrossRef]
- Jochems, L.W.; Brandt, J.; Monks, A.; Cattau, M.; Kolarik, N.; Tallant, J.; Lishawa, S. Comparison of Different Analytical Strategies for Classifying Invasive Wetland Vegetation in Imagery from Unpiloted Aerial Systems (UAS). Remote Sens. 2021, 13, 4733. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef] [PubMed]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, Z.; Qiao, H.; Wang, R.; Wei, H.; Wang, L.; Gu, W.; Li, X. Challenges and Development Trend of Species Distribution Model. Adv. Earth Sci. 2020, 35, 1292–1305. [Google Scholar] [CrossRef]
- Huang, X.; He, F.; Peng, Y.; Chen, S. Distribution of Alien Invasive Species Ambrosia artemisiifolia in Guangxi Area. Weed Sci. 2023, 41, 29–34. [Google Scholar] [CrossRef]
- Marmion, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. The performance of state-of-the-art modelling techniques depends on geographical distribution of species. Ecol. Model. 2009, 220, 3512–3520. [Google Scholar] [CrossRef]
- Dittrich, A.; Roilo, S.; Sonnenschein, R.; Cerrato, C.; Ewald, M.; Viterbi, R.; Cord, A.F. Modelling Distributions of Rove Beetles in Mountainous Areas Using Remote Sensing Data. Remote Sens. 2020, 12, 80. [Google Scholar] [CrossRef]
- Dong, J.; Guo, M.; Wang, X.; Yang, X.; Zhang, Y.; Zhang, P. Dramatic loss of seagrass Zostera marina L. suitable habitat under projected climate change in coastal areas of the Bohai Sea and Shandong peninsula, China. J. Exp. Mar. Biol. Ecol. 2023, 565, 151915. [Google Scholar] [CrossRef]
- Keyghobadi, M.; Piri Sahragard, H.; Pahlavan Rad, M.R.; Karami, P.; Yari, R. Application of Generalized Additive Model and Classification and Regression Tree to Estimate Potential Habitat Distribution of Range plant species (Case Study: Khazri Rangelands of Beyaz Plain, Southern Khorasan). Iran. J. Range Desert Res. 2020, 27, 561–576. [Google Scholar] [CrossRef]
- Aidoo, O.F.; Souza, P.G.C.; Da Silva, R.S.; Júnior, P.A.S.; Picanço, M.C.; Osei-Owusu, J.; Sétamou, M.; Ekesi, S.; Borgemeister, C. A machine learning algorithm-based approach (MaxEnt) for predicting invasive potential of Trioza erytreae on a global scale. Ecol. Inform. 2022, 71, 101792. [Google Scholar] [CrossRef]
- Fan, J.; Li, H.; Yang, Z.; Zhu, G. Selecting the best native individual model to predict potential distribution of Cabomba caroliniana in China. Biodivers. Sci. 2019, 27, 140–148. [Google Scholar] [CrossRef]
- Briscoe Runquist, R.D.; Lake, T.; Tiffin, P.; Moeller, D.A. Species distribution models throughout the invasion history of Palmer amaranth predict regions at risk of future invasion and reveal challenges with modeling rapidly shifting geographic ranges. Sci. Rep. 2019, 9, 2426. [Google Scholar] [CrossRef] [PubMed]
- Panda, R.M.; Behera, M.D.; Roy, P.S. Assessing distributions of two invasive species of contrasting habits in future climate. J. Environ. Manag. 2018, 213, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Hamit, S.; Abdushalih, N.; Jiesisi, A.; Hua, S.; Yilihar, V. Impact of human activities on potential distribution of Solanum rostratum Dunal in Xinjiang. Acta Ecol. Sin. 2019, 39, 629–636. [Google Scholar] [CrossRef]
- Teng, M.; Liu, J.; Lu, Y.; Cheng, X.; Wang, Y. Simulation of the distribution of wild Alligator sinensis (Crocodylia: Alligatoridae) in China under climate change. Acta Ecol. Sin. 2023, 43, 1–11. [Google Scholar] [CrossRef]
- Shi, X.; Wang, J.; Zhang, L.; Chen, S.; Zhao, A.; Ning, X.; Fan, G.; Wu, N.; Zhang, L.; Wang, Z. Prediction of the potentially suitable areas of Litsea cubeba in China based on future climate change using the optimized MaxEnt model. Ecol. Indic. 2023, 148, 110093. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; El-Kassaby, Y.A.; Luo, D.; Guo, J.; He, X.; Zhao, G.; Tian, X.; Qiu, J.; Feng, Z.; et al. Planning Ginkgo biloba future fruit production areas under climate change: Application of a combinatorial modeling approach. For. Ecol. Manag. 2023, 533, 120861. [Google Scholar] [CrossRef]
- Yao, Z.; Han, Q.; Lin, B. Prediction of distribution area of main noxious and miscellaneous weeds in Xinjiang based on MaxEnt model. Acta Ecol. Sin. 2023, 43, 1–14. [Google Scholar] [CrossRef]
- Ju, X.; Lin, J.; Wu, J.; Chen, J.; Guan, J.; Li, M.; Zheng, J. Prediction of potential living area of typical locusts in Xinjiang based on species distribution model. Acta Ecol. Sin. 2022, 42, 8605–8617. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, W.; Ye, H.; Lu, L. Study on the Identification of Habitat Suitability Areas for the Dominant Locust Species Dasyhippus Barbipes in Inner Mongolia. Remote Sens. 2023, 15, 1718. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, J.; Wang, M.; Li, Z.; Lin, S.; Chen, H. Potential distribution prediction of Amaranthus palmeri S. Watson in China under current and future climate scenarios. Ecol. Evol. 2022, 12, e9505. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.; Xiong, Q.; Qiu, X.; Zhang, Y. Dynamics of invasive alien plant species in China under climate change scenarios. Ecol. Indic. 2021, 129, 107919. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Uddin, M.P.; Mamun, M.A.; Hossain, M.A. PCA-based Feature Reduction for Hyperspectral Remote Sensing Image Classification. Tech. Rev. IETE 2021, 38, 377–396. [Google Scholar] [CrossRef]
- Jiang, J.; Ma, J.; Chen, C.; Wang, Z.; Cai, Z.; Wang, L. SuperPCA: A Superpixelwise PCA Approach for Unsupervised Feature Extraction of Hyperspectral Imagery. IEEE Trans. Geosci. Remote Sens. 2018, 56, 4581–4593. [Google Scholar] [CrossRef]
- Uddin, M.P.; Mamun, M.A.; Afjal, M.I.; Hossain, M.A. Information-theoretic feature selection with segmentation-based folded principal component analysis (PCA) for hyperspectral image classification. Int. J. Remote Sens. 2021, 42, 286–321. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, W.; Yuan, Z.; Song, Z.; Wang, Z.; Gao, J.; Fu, W.; Zhang, G. Chromosome-level genome assembly and annotation of the prickly nightshade Solanum rostratum Dunal. Sci. Data 2023, 10, 341. [Google Scholar] [CrossRef]
- Lin, Y.; Tan, D.Y. The potential and exotic invasive plant: Solanum rostratum. Acta Phytotaxon. Sin. 2007, 45, 675–685. [Google Scholar] [CrossRef]
- Yan, W.; Wang, J.; Zheng, Y. Advances in Hazard status and Control technology of Solanum rostratum. Terr. Ecosyst. Conserv. 2022, 2, 73–79. [Google Scholar] [CrossRef]
- Bao, H.; Fu, J.; Wang, Y.; Luo, J.; Liu, D.; Shao, R.; Zhang, L. Distribution pattern and spread impact factors of invasive plant buffalobur in Inner Mongolia. Inn. Mong. Environ. Sci. 2022, 34, 104–108. [Google Scholar] [CrossRef]
- Zhao, X.-Y.; Ma, X.-D.; Xu, Z.-W. Non-native Solanum rostratum and Its Ecological Significance Appeared in Urumqi, Xinjiang. Adv. Earth Sci. 2007, 22, 167–170. [Google Scholar] [CrossRef]
- Chen, T.; Liu, Z.; Lou, A. Phenotypic variation in populations of Solanum rostratum in different distribution areas in China. Chin. J. Plant Ecol. 2013, 37, 344–353. [Google Scholar] [CrossRef]
- Guo, J.; Cao, W.; Zhang, Y.; Gao, Y.; Wang, Y. Prediction of the potential distribution area of Solanum rostratum in northeast China. Pratacultural Sci. 2019, 36, 2476–2484. [Google Scholar] [CrossRef]
- Wang, Q.; Cheng, M.; Xiao, X.; Yuan, H.; Zhu, J.; Fan, C.; Zhang, J. An image segmentation method based on deep learning for damage assessment of the invasive weed Solanum rostratum Dunal. Comput. Electron. Agric. 2021, 188, 106320. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H. Survey on major alien invasive plants and their prevention and control in Jizhou District, Tianjin, China. Sci. Technol. Tianjin Agric. For. 2017, 27–29. [Google Scholar] [CrossRef]
- Che, J.; Liu, Q.; Hu, B. Alien invasive weed Solanum rostratum Dunal. Weed Sci. 2006, 58–60. [Google Scholar] [CrossRef]
- Xiang, J.; Li, C.-N.; Liu, Q.-R.; Zhou, Y.-L.; Sun, L.; Mao, C.-M.; Liang, Q.-J. Ecological state of invasive alien plant Solanum rostratum in Beijing. Chin. J. Ecol. 2011, 30, 453–458. [Google Scholar] [CrossRef]
- Technical Guidance for Prevention and Control of Solanum rostratum Dunal. Available online: http://www.reea.agri.cn/stzybh/202307/t20230707_8004181.htm (accessed on 13 September 2023).
- Wu, Z.-G.; Fang, Y.; Qin, M.; Qin, Y.-J.; Wang, C.; Zhao, T.; Li, Z.-H. Potential economic loss assessment on maize industry of China caused by buffalobur (Solanum rostratum). J. China Agric. Univ. 2015, 20, 138–145. [Google Scholar] [CrossRef]
- Wang, H. Study on the Invasion Characteristics of Solanum rostratum Dunal. Master’s Thesis, Inner Mongolia Normal University, Hohhot, China, 2017. [Google Scholar]
- Zhang, L.; Lou, A. Phenotypic variation in floral traits of an invasive plant (Solanum rostratum) and its impact on reproductive fitness. Sci. Sin. Vitae 2022, 52, 1281–1291. [Google Scholar] [CrossRef]
- Qiu, J.; Shalimu, D.; Tan, D. Reproductive characteristics of the invasive species Solanum rostratum indifferent habitats of Xinjiang, China. Biodivers. Sci. 2013, 21, 590–600. [Google Scholar] [CrossRef]
- Shi, K.; Shao, H.; Han, C.; Zokir, T. Diversity of the Rhizosphere Soil Fungi of the Invasive Plant (Solanum rostratum Dunal) and the Allelopathic Potential of Their Secondary Metabolites. Chin. J. Soil Sci. 2022, 53, 548–557. [Google Scholar] [CrossRef]
- Abu-Nassar, J.; Matzrafi, M. Effect of Herbicides on the Management of the Invasive Weed Solanum rostratum Dunal (Solanaceae). Plants 2021, 10, 284. [Google Scholar] [CrossRef]
- Wang, Q.; Cheng, M.; Huang, S.; Cai, Z.; Zhang, J.; Yuan, H. A deep learning approach incorporating YOLO v5 and attention mechanisms for field real-time detection of the invasive weed Solanum rostratum Dunal seedlings. Comput. Electron. Agric. 2022, 199, 107194. [Google Scholar] [CrossRef]
- Zhong, G.; Shen, W.; Wan, F.; Wang, J. Potential distribution areas of Solanum rostratum in China: A prediction with GARP niche model. Chin. J. Ecol. 2009, 28, 162–166. [Google Scholar] [CrossRef]
- Lv, F. Studies on the Potential Suitable Distribution Areas and Chemical Constituents of the Invasive Plant Solanum rostratum. Master’s Thesis, Shenyang Agriculture University, Shenyang, China, 2020. [Google Scholar]
- Wang, R.; Tang, Y.; Zhang, Z.; Wan, F. The distribution pattern and early monitoring for preventing further expansion of Solanum rostratum in China. J. Biosaf. 2018, 27, 284–289. [Google Scholar] [CrossRef]
- Su, H.; Sihake, N.; Bazhabaike, M. Study on the invasion status of the alien invasive plant Solanum rostratum Dunal in Xinjiang. Xinjiang Agric. Sci. Technol. 2022, 262, 21–22. [Google Scholar] [CrossRef]
- Tian, Z.; Zeng, J.; Wang, Y.; Zhu, Q. Distribution and Risk Assessment of Invasive Plant Solanum rostratum Dunal in Ningxia. Ningxia J. Agric. For. Sci. Technol. 2021, 62, 61–64. [Google Scholar] [CrossRef]
- Qu, Z. Occurrence and Control Status of Exotic Invasive Plant Solanum rostratum in Liaoning Province. Agric. Sci. Technol. Equip. 2021, 14–15. [Google Scholar] [CrossRef]
- Chen, J.; Ma, F.-Z.; Zhang, Y.-J.; Wang, C.-B.; Xu, H.-G. Spatial point pattern analysis of Solanum rostratum Dunal in different habitats. J. South. Agric. 2020, 51, 342–349. [Google Scholar] [CrossRef]
- Yimingniyazi, A. Effect of Solanum rostratum Invasion on Distribution, Life History and Habits of Leptinotarsa decemlineata. Ph.D. Thesis, Xinjiang Agricultural University, Urumqi, China, 2015. [Google Scholar]
- Song, Z.; Tan, D.; Zhou, G. Distribution and Community Characteristics of Invasive Solanum rostratum Dunal. in Xinjiang. Arid. Res. 2013, 30, 129–134. [Google Scholar] [CrossRef]
- He, J.; Khasbag, A.; Mong, E.; Hu, M. Solanum rostratum Dunal: A Newly Invaded Alien Plant of Inner Mongolia. J. Inn. Mong. Norm. Univ. Nat. Sci. Ed. 2011, 40, 288–290. [Google Scholar] [CrossRef]
- Shalim, D. Pollination Biology and Germination Characteristics of Solanum rostratum. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2011. [Google Scholar]
- Wang, C.; Hong, Y.; Sun, L. Analysis on the biological characteristics and control methods of the Solanum rostratum Dunal occurring in Baicheng City. Jilin Agric. 2010, 83. [Google Scholar]
- National Plant Specimen Resource Center. Available online: https://www.cvh.ac.cn (accessed on 3 August 2023).
- Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 3 August 2023).
- For the First Time in East China, the Invasive Species Solanum rostratum Dunal Has Been Documented in Our Region. Available online: http://www.wuxing.gov.cn/art/2022/8/26/art_1229518306_3888984.html (accessed on 3 August 2023).
- Baidu Coordinate Retrieval System. Available online: http://api.map.baidu.com/lbsapi/getpoint/index.html (accessed on 3 August 2023).
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Zhu, C.; Qu, Y.; Wang, H. Predicting the potential distribution of an invasive species, Erigeron canadensis L., in China with a maximum entropy model. Glob. Ecol. Conserv. 2020, 21, e00822. [Google Scholar] [CrossRef]
- Zeng, Y.; Low, B.W.; Yeo, D.C.J. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish. Ecol. Model. 2016, 341, 5–13. [Google Scholar] [CrossRef]
- Ikotun, A.M.; Ezugwu, A.E.; Abualigah, L.; Abuhaija, B.; Heming, J. K-means clustering algorithms: A comprehensive review, variants analysis, and advances in the era of big data. Inf. Sci. 2023, 622, 178–210. [Google Scholar] [CrossRef]
- Cap, O.M.; Rez, A.P.E.; Lozano, J.A. An efficient K-means clustering algorithm for tall data. Data Min. Knowl. Discov. 2020, 34, 776–811. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Harris, R.; Jarvis, C. Statistics for Geography and Environmental Science; Routledge: London, UK, 2014. [Google Scholar] [CrossRef]
- National Statistical Yearbook 2022. Available online: https://data.stats.gov.cn/easyquery.htm?cn=C01 (accessed on 3 August 2023).
- Liu, D.; Zhang, X. Occurrence Prediction of Pine Wilt Disease Based on CA–Markov Model. Forests 2022, 13, 1736. [Google Scholar] [CrossRef]
- Zhang, S.; Wei, S.; Zhang, C.; Huang, H.; Cui, H.; Li, X.; Wang, J. Research Advances on Seed Dormancy and Germination of Buffalobur (Solanum rostratum). Weed Sci. 2011, 29, 5–9. [Google Scholar] [CrossRef]
- Halvorsen, R.; Mazzoni, S.; Bryn, A.; Bakkestuen, V. Opportunities for improved distribution modelling practice via a strict maximum likelihood interpretation of MaxEnt. Ecography 2015, 38, 172–183. [Google Scholar] [CrossRef]
- Halvorsen, R.; Mazzoni, S.; Dirksen, J.W.; Næsset, E.; Gobakken, T.; Ohlson, M. How important are choice of model selection method and spatial autocorrelation of presence data for distribution modelling by MaxEnt? Ecol. Model. 2016, 328, 108–118. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Wheeler, D.C. Diagnostic tools and a remedial method for collinearity in geographically weighted regression. Environ. Plan. A 2007, 39, 2464–2481. [Google Scholar] [CrossRef]
- Cruz-Cárdenas, G.; López-Mata, L.; Villaseñor, J.L.; Ortiz, E. Potential species distribution modeling and the use of principal component analysis as predictor variables. Rev. Mex. Biodivers. 2014, 85, 189–199. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Number of Features | Year | Remark | Source |
|---|---|---|---|---|
| Climatic variables | 19 | Current:1970–2020; Future:2021–2040, 2041–2060. | 19 bioclimatic factors | WorldClim database (https://www.worldclim.org/ (accessed on 3 August 2023)) |
| Terrain variables | 3 | / | Elevation, slope, and aspect | WorldClim database (https://www.worldclim.org/ (accessed on 3 August 2023)) |
| Soil variables | 98 | 2010–2018 | Grid data for various soil properties at different depths | National Tibetan Plateau Data Center (http://data.tpdc.ac.cn (accessed on 3 August 2023)) |
| Humanistic variables | 5 | 2020 | GDP, population, distances to roads and water bodies, building density | GDP, population: Resource and Environmental Science and Data Center (https://www.resdc.cn/Default.aspx (accessed on 3 August 2023)) Roads and water bodies: OpenStreetMap (https://www.openstreetmap.org (accessed on 3 August 2023)) Building: Geoservice of the Earth Observation Center (EOC) of the German Aerospace Center (DLR) (https://download.geoservice.dlr.de/WSF2019 (accessed on 3 August 2023)) |
| Spatial variables | 2 | / | Longitude and latitude | Extracted from Elevation |
| Simulation Scenario | Number of Input Variables | Number of New Variables | PCs Name |
|---|---|---|---|
| current 1 (1970–2020) | 127 a | 11 | pc1, pc2, pc3……, pc11 |
| current 2 (1970–2020) | 24 b | 4 | pc1, pc2, pc3, pc4 |
| future 1 (2021–2040: SSP1-2.6, SSP5-8.5) | 24 b | 4 | pc1, pc2, pc3, pc4 |
| future 2 (2041–2060: SSP1-2.6, SSP5-8.5) | 24 b | 4 | pc1, pc2, pc3, pc4 |
| Cluster Names | Contribution Rate (%) | Variables | Cluster Names | Contribution Rate (%) | Variables |
|---|---|---|---|---|---|
| C1 | 40.99 | total potassium density, total potassium, total nitrogen, total nitrogen density, total phosphorus, total phosphorus density | C7 | 8.80 | soil organic carbon, soil organic carbon density |
| C2 | 12.70 | coarse fragment content, sand (0.05–2 mm) | C8 | 2.15 | cation exchange capacity |
| C3 | 7.08 | pH | C9 | 2.05 | geographical longitude, geographical latitude |
| C4 | 7.14 | bio3, bio6, bio11, bio12, bio13, bio15, bio16, bio18 | C10 | 1.27 | distance from building ups, gross domestic product (GDP) within grid, population count within each grid cell (1 square kilometer) |
| C5 | 4.53 | bio4, bio5, bio7, bio10 | C11 | 1.06 | elevation, aspect, slope |
| C6 | 4.12 | bulk density, thickness |
| ID | The Types of Input Environmental Variables | Training AUC | Test AUC | Mean AUC | Remark |
|---|---|---|---|---|---|
| 1 | A+B+C+D+E | 0.950 | 0.980 | 0.941 | current 1 |
| 2 | A+B+D+E | 0.943 | 0.895 | 0.934 | |
| 3 | A+B+E | 0.934 | 0.929 | 0.925 | current 2 |
| 4 | A+B+C+E | 0.944 | 0.956 | 0.935 | |
| 5 | B+C+D+E | 0.948 | 0.971 | 0.933 | |
| 6 | A+B+C+D | 0.945 | 0.939 | 0.934 |
| ID | Method | Number of input Variables | Number of Variables after Decorrelation | Training AUC | Test AUC | Mean AUC | Remark |
|---|---|---|---|---|---|---|---|
| 1 | PCA | 127 | 11 | 0.950 | 0.980 | 0.941 | / |
| 2 | Spearman Correlation | 127 | 73 | 0.811 | 0.874 | 0.763 | 0.75 [29] |
| 3 | 70 | 0.897 | 0.911 | 0.861 | 0.6 | ||
| 4 | 62 | 0.889 | 0.947 | 0.866 | 0.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.; Yang, T.; Wang, K.; Huang, W. Assessing the Current and Future Potential Distribution of Solanum rostratum Dunal in China Using Multisource Remote Sensing Data and Principal Component Analysis. Remote Sens. 2024, 16, 271. https://doi.org/10.3390/rs16020271
Huang T, Yang T, Wang K, Huang W. Assessing the Current and Future Potential Distribution of Solanum rostratum Dunal in China Using Multisource Remote Sensing Data and Principal Component Analysis. Remote Sensing. 2024; 16(2):271. https://doi.org/10.3390/rs16020271
Chicago/Turabian StyleHuang, Tiecheng, Tong Yang, Kun Wang, and Wenjiang Huang. 2024. "Assessing the Current and Future Potential Distribution of Solanum rostratum Dunal in China Using Multisource Remote Sensing Data and Principal Component Analysis" Remote Sensing 16, no. 2: 271. https://doi.org/10.3390/rs16020271