
Using Satellite Imagery and Aerial Orthophotos for the Multi-Decade Monitoring of Subalpine Norway Spruce Stands Changes in Gorce National Park, Poland


Abstract
:1. Introduction
- Examining which of the land cover (LC) classes have undergone the most significant changes, using archival remote sensing data;
- Determining the characteristics of dynamics of changes occurring in selected land cover classes using GIS analyses and archival remote sensing data.
2. Study Area
3. Materials and Methods
3.1. Materials
3.2. Methods
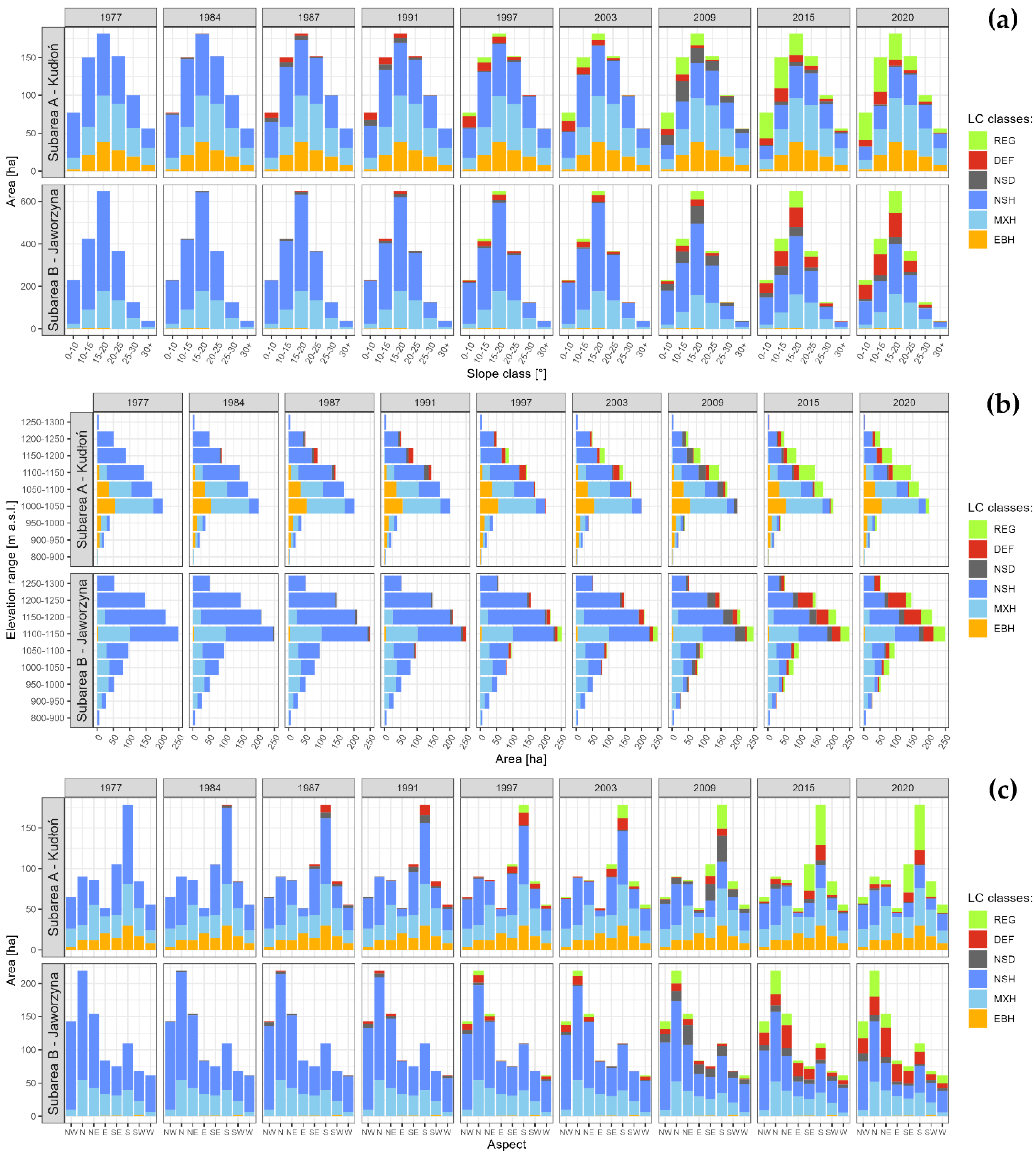
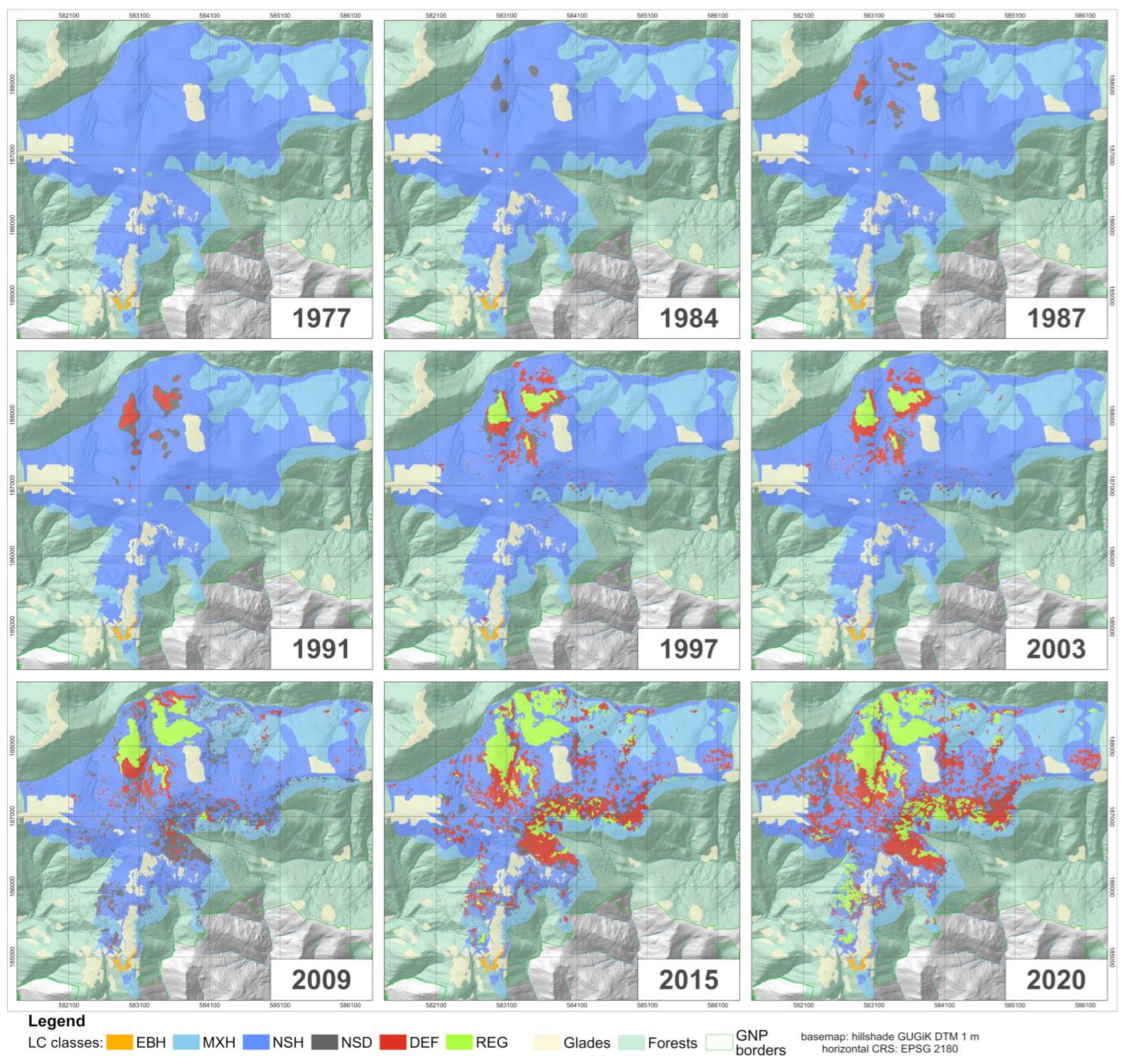
4. Results
4.1. Land Cover Class Mapping
4.2. Slope, Elevation and Aspect Analysis and Comparison of Subareas
4.3. Dynamics of Land Cover Classes
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- White, P.S.; Pickett, S.T.A. Natural Disturbance and Patch Dynamics: An Introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Elsevier: Amsterdam, The Netherlands, 1985; pp. 3–13. [Google Scholar]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest Disturbances under Climate Change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Viljur, M.; Abella, S.R.; Adámek, M.; Alencar, J.B.R.; Barber, N.A.; Beudert, B.; Burkle, L.A.; Cagnolo, L.; Campos, B.R.; Chao, A.; et al. The Effect of Natural Disturbances on Forest Biodiversity: An Ecological Synthesis. Biol. Rev. 2022, 97, 1930–1947. [Google Scholar] [CrossRef]
- Schurman, J.S.; Trotsiuk, V.; Bače, R.; Čada, V.; Fraver, S.; Janda, P.; Kulakowski, D.; Labusova, J.; Mikoláš, M.; Nagel, T.A.; et al. Large-Scale Disturbance Legacies and the Climate Sensitivity of Primary Picea abies Forests. Glob. Chang. Biol. 2018, 24, 2169–2181. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing Forest Disturbances in Europe and Their Impact on Carbon Storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Janda, P.; Trotsiuk, V.; Mikoláš, M.; Bače, R.; Nagel, T.A.; Seidl, R.; Seedre, M.; Morrissey, R.C.; Kucbel, S.; Jaloviar, P.; et al. The Historical Disturbance Regime of Mountain Norway Spruce Forests in the Western Carpathians and Its Influence on Current Forest Structure and Composition. For. Ecol. Manag. 2017, 388, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Albrich, K.; Rammer, W.; Thom, D.; Seidl, R. Trade-Offs between Temporal Stability and Level of Forest Ecosystem Services Provisioning under Climate Change. Ecol. Appl. 2018, 28, 1884–1896. [Google Scholar] [CrossRef]
- Gratzer, G.; Alemayehu Duguma, L.; Kohler, T.; Maselli, D.; Romeo, R. Mountain Forests in a Changing World; FAO/MPS and SDC: Rome, Italy, 2011. [Google Scholar]
- Szewczyk, J.; Szwagrzyk, J.; Muter, E. Tree Growth and Disturbance Dynamics in Oldgrowth Subalpine Spruce Forests of the Western Carpathians. Can. J. For. Res. 2011, 41, 938–944. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests: Vegetation Ecology of Central Europe; Springer International Publishing: Cham, Switzerland, 2017; Volume 1, ISBN 978-3-319-43042-3. [Google Scholar]
- Prinz, B. Major Hypotheses and Factors Causes of Forest Damage in Europe. Environment 1987, 29, 11–37. [Google Scholar] [CrossRef]
- Blank, L.W. A New Type of Forest Decline in Germany. Nature 1985, 314, 311–314. [Google Scholar] [CrossRef]
- Holeksa, J. Rozpad Drzewostanu i Odnowienie Świerka a Struktura i Dynamika Karpackiego Boru Górnoreglowego. Monogr. Bot. 1998, 82, 1–209. [Google Scholar] [CrossRef] [Green Version]
- Rehfuess, K.E. On the Causes of Decline of Norway Spruce (Picea abies Karst.) in Central Europe. Soil Use Manag. 1985, 1, 30–32. [Google Scholar] [CrossRef]
- Schütt, P.; Koch, W.; Blaschke, H.; Land, K.J.; Schluck, H.J.; Summerer, H. So Stirbt Der Wald. Schadbilder Und Krankheitsverlauf; BLV-Verlag: Munich, Germany, 1983. [Google Scholar]
- Manion, P. Tree Disease Concepts; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1981. [Google Scholar]
- Bałazy, R. Forest Dieback Process in the Polish Mountains in the Past and Nowadays—Literature Review on Selected Topics. Folia For. Pol. 2020, 62, 184–198. [Google Scholar] [CrossRef]
- Synek, M.; Janda, P.; Mikoláš, M.; Nagel, T.A.; Schurman, J.S.; Pettit, J.L.; Trotsiuk, V.; Morrissey, R.C.; Bače, R.; Čada, V.; et al. Contrasting Patterns of Natural Mortality in Primary Picea Forests of the Carpathian Mountains. For. Ecol. Manag. 2020, 457, 117734. [Google Scholar] [CrossRef]
- Jandl, R. Climate-Induced Challenges of Norway Spruce in Northern Austria. Trees For. People 2020, 1, 100008. [Google Scholar] [CrossRef]
- Honkaniemi, J.; Rammer, W.; Seidl, R. Norway Spruce at the Trailing Edge: The Effect of Landscape Configuration and Composition on Climate Resilience. Landsc. Ecol. 2020, 35, 591–606. [Google Scholar] [CrossRef]
- Jansen, S.; Konrad, H.; Geburek, T. The Extent of Historic Translocation of Norway Spruce Forest Reproductive Material in Europe. Ann. For. Sci. 2017, 74, 56. [Google Scholar] [CrossRef]
- Hais, M.; Langhammer, J.; Jirsová, P.; Dvořák, L. Dynamics of Forest Disturbance in Central Part of the Šumava Mountains between 1985 and 2007 Based on Landsat TM/ETM+ Satellite Data. Acta Univ. Carol. Geogr. 2008, 43, 53–62. [Google Scholar] [CrossRef]
- van der Knaap, W.O.; van Leeuwen, J.F.N.; Fahse, L.; Szidat, S.; Studer, T.; Baumann, J.; Heurich, M.; Tinner, W. Vegetation and Disturbance History of the Bavarian Forest National Park, Germany. Veg. Hist. Archaeobot. 2019, 29, 277–295. [Google Scholar] [CrossRef]
- Obladen, N.; Dechering, P.; Skiadaresis, G.; Tegel, W.; Keßler, J.; Höllerl, S.; Kaps, S.; Hertel, M.; Dulamsuren, C.; Seifert, T.; et al. Tree Mortality of European Beech and Norway Spruce Induced by 2018-2019 Hot Droughts in Central Germany. Agric. For. Meteorol. 2021, 307, 108482. [Google Scholar] [CrossRef]
- Krejza, J.; Cienciala, E.; Světlík, J.; Bellan, M.; Noyer, E.; Horáček, P.; Štěpánek, P.; Marek, M.V. Evidence of Climate-Induced Stress of Norway Spruce along Elevation Gradient Preceding the Current Dieback in Central Europe. Trees 2020, 35, 103–119. [Google Scholar] [CrossRef]
- Stereńczak, K.; Mielcarek, M.; Kamińska, A.; Kraszewski, B.; Piasecka, Ż.; Miścicki, S.; Heurich, M. Influence of Selected Habitat and Stand Factors on Bark Beetle Ips typographus (L.) Outbreak in the Białowieża Forest. For. Ecol. Manag. 2020, 459, 117826. [Google Scholar] [CrossRef]
- Zajączkowski, G.; Jabłoński, M.; Jabłoński, T.; Szmidla, H.; Kowalska, A.; Małachowska, J.; Piwnicki, J. Raport o Stanie Lasów w Polsce 2020; PGL Lasy Państwowe: Warszawa, Poland, 2020; pp. 1–119. [Google Scholar]
- Koprowski, M.; Zielski, A. Dendrochronology of Norway Spruce (Picea abies (L.) Karst.) from Two Range Centres in Lowland Poland. Trees 2006, 20, 383–390. [Google Scholar] [CrossRef]
- Jurek, E.; Olszowska, G.; Olszowski, J. Zamieranie Drzewostanów Świerkowych w Rejonie Gór Izerskich. Sylwan 1983, 127, 13–20. [Google Scholar]
- Jarocińska, A.; Zagajewski, B.; Ochtyra, A.; Marcinkowska, A.; Kycko, M.; Pabjanek, P. Przebieg Klęski Ekologicznej w Karkonoszach i Górach Izerskich Na Podstawie Analizy Zdjęć Satelitarnych Landsat. In Proceedings of the 25 lat po klęsce ekologicznej w Karkonoszach i Górach Izerskich—Obawy a Rzeczywistość; Knapik, R., Ed.; Karkonoski Park Narodowy: Jelenia Góra, Poland, 2014; pp. 47–62. [Google Scholar]
- Ochtyra, A. Forest Disturbances in Polish Tatra Mountains for 1985–2016 in Relation to Topography, Stand Features, and Protection Zone. Forests 2020, 11, 579. [Google Scholar] [CrossRef]
- Capecki, Z. Masowe Wystąpienie Zasnui Wysokogórskiej w Gorcach. Sylwan 1982, 126, 41–50. [Google Scholar]
- Honowski, J.; Huflejt, T. O Występowaniu Zasnui Wysokogórskiej Cephalcia falleni (Dalm.), w Gorczańskim Parku Narodowym. Pol. Pismo Entomol. 1988, 58, 433–445. [Google Scholar]
- Loch, J.; Wężyk, P.; Małek, S.; Pająk, M. Natural Regeneration versus Tree Planting in the Subalpine Spruce Forest Plagiothecio-Piceetum tatricum of the Gorce National Park (Southern Poland). Nat. Conserv. 2001, 58, 5–15. [Google Scholar]
- Grodzki, W.; Loch, J.; Armatys, P. Występowanie Kornika Drukarza Ips typographus L. w Uszkodzonych Przez Wiatr Drzewostanach Świerkowych Masywu Kudłonia w Gorczańskim Parku Narodowym Occurrence of Ips typographus L. in Wind-Damaged Norway Spruce Stands of Kudłoń Massif in the Gorce Natio. Ochr. Beskidów Zachodnich 2006, 1, 125–137. [Google Scholar]
- Chwistek, K. Zmiany Składu Gatunkowego i Struktury Drzewostanów Gorczańskiego Parku Narodowego w Latach 1992–2007. Ochr. Beskidów Zachodnich 2010, 3, 79–92. [Google Scholar]
- O’Brien, L.; Schuck, A.; Fraccaroli, C.; Pötzelsberger, E.; Winkel, G.; Lindner, M. Protecting Old-Growth Forests in Europe—A Review of Scientific Evidence to Inform Policy Implementation; European Forest Institute: Joensuu, Finland, 2021. [Google Scholar]
- Loch, J. Świerczyny górnoreglowe Gorczańskiego Parku Narodowego-stan zachowania i dynamika zmian. Probl. Zagospod. Ziem Górskich 2002, 48, 185–193. [Google Scholar]
- Krzystek, P.; Serebryanyk, A.; Schnörr, C.; Červenka, J.; Heurich, M. Large-Scale Mapping of Tree Species and Dead Trees in Sumava National Park and Bavarian Forest National Park Using Lidar and Multispectral Imagery. Remote Sens. 2020, 12, 661. [Google Scholar] [CrossRef]
- Wężyk, P.; Hawryło, P.; Janus, B.; Weidenbach, M.; Szostak, M. Forest Cover Changes in Gorce NP (Poland) Using Photointerpretation of Analogue Photographs and GEOBIA of Orthophotos and NDSM Based on Image-Matching Based Approach. Eur. J. Remote Sens. 2018, 51, 501–510. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R. Mapping the Forest Disturbance Regimes of Europe. Nat. Sustain. 2021, 4, 63–70. [Google Scholar] [CrossRef]
- Meigs, G.W.; Kennedy, R.E.; Cohen, W.B. A Landsat Time Series Approach to Characterize Bark Beetle and Defoliator Impacts on Tree Mortality and Surface Fuels in Conifer Forests. Remote Sens. Environ. 2011, 115, 3707–3718. [Google Scholar] [CrossRef]
- Migas-Mazur, R.; Kycko, M.; Zwijacz-Kozica, T.; Zagajewski, B. Assessment of Sentinel-2 Images, Support Vector Machines and Change Detection Algorithms for Bark Beetle Outbreaks Mapping in the Tatra Mountains. Remote Sens. 2021, 13, 3314. [Google Scholar] [CrossRef]
- Miczyński, J. Klimat Rządzi Przyrodą. In Gorczański Park Narodowy. Przyroda i Krajobraz pod Ochroną; Gorczański Park Narodowy: Poręba Wielka, Poland, 2015; pp. 35–38. [Google Scholar]
- Protection Plan of Gorce National Park, Journal of Laws of the Republic of Poland, Item 579. 2022; pp. 4–5. Available online: https://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20220000579/O/D20220579.pdf (accessed on 6 February 2023).
- Tomasiewicz, J. Bory Regla Górnego. In Gorczański Park Narodowy. Przyroda i Krajobraz pod Ochroną; Gorczański Park Narodowy: Poręba Wielka, Poland, 2015; pp. 82–89. [Google Scholar]
- European Council. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora 1992. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A01992L0043-20130701 (accessed on 6 February 2023).
- European Parliament and the Council Directive 2009/147/EC of the European Parliament and of the Council of 30 November 2009 on the Conservation of Wild Birds 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A02009L0147-20190626 (accessed on 6 February 2023).
- QGIS Association QGIS Geographic Information System 2020. Available online: https://qgis.org/site/ (accessed on 6 February 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating Outbreak of Bark Beetles in the Czech Republic: Drivers, Impacts, and Management Implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Faccoli, M.; Bernardinelli, I. Composition and Elevation of Spruce Forests Affect Susceptibility to Bark Beetle Attacks: Implications for Forest Management. Forests 2014, 5, 88–102. [Google Scholar] [CrossRef]
- Jonášová, M.; Matějková, I. Natural Regeneration and Vegetation Changes in Wet Spruce Forests after Natural and Artificial Disturbances. Can. J. For. Res. 2007, 37, 1907–1914. [Google Scholar] [CrossRef]
- Zeppenfeld, T.; Svoboda, M.; DeRose, R.J.; Heurich, M.; Müller, J.; Čížková, P.; Starý, M.; Bače, R.; Donato, D.C. Response of Mountain Picea abies Forests to Stand-Replacing Bark Beetle Outbreaks: Neighbourhood Effects Lead to Self-Replacement. J. Appl. Ecol. 2015, 52, 1402–1411. [Google Scholar] [CrossRef]
- Kausrud, K.; Økland, B.; Skarpaas, O.; Grégoire, J.-C.; Erbilgin, N.; Stenseth, N. Chr. Population Dynamics in Changing Environments: The Case of an Eruptive Forest Pest Species. Biol. Rev. 2012, 87, 34–51. [Google Scholar] [CrossRef] [PubMed]
- Jakuš, R.; Edwards-Jonášová, M.; Cudlín, P.; Blaženec, M.; Ježík, M.; Havlíček, F.; Moravec, I. Characteristics of Norway Spruce Trees (Picea abies) Surviving a Spruce Bark Beetle (Ips typographus L.) Outbreak. Trees 2011, 25, 965–973. [Google Scholar] [CrossRef]
- Seidl, R.; Müller, J.; Hothorn, T.; Bässler, C.; Heurich, M.; Kautz, M. Small Beetle, Large-Scale Drivers: How Regional and Landscape Factors Affect Outbreaks of the European Spruce Bark Beetle. J. Appl. Ecol. 2016, 53, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Klopcic, M.; Poljanec, A.; Gartner, A.; Boncina, A. Factors Related to Natural Disturbances in Mountain Norway Spruce (Picea abies) Forests in the Julian Alps. Écoscience 2009, 16, 48–57. [Google Scholar] [CrossRef]
- Bałazy, R.; Hycza, T.; Kamińska, A.; Osińska-Skotak, K. Factors Affecting the Health Condition of Spruce Forests in Central European Mountains-Study Based on Multitemporal RapidEye Satellite Images. Forests 2019, 10, 943. [Google Scholar] [CrossRef]
- Despres, T.; VaItkov, L.; Bace, R.; Cada, V.; Janda, P.; Mikolas, M.; Schurman, J.S.; Trotsiuk, V.; Svoboda, M. Past Disturbances and Intraspecific Competition as Drivers of Spatial Pattern in Primary Spruce Forests. Ecosphere 2017, 8, e02037. [Google Scholar] [CrossRef]
- Wężyk, P.; Mansberger, R. Techniki Fotogrametrii Cyfrowej i GIS w Ocenie Degradacji Drzewostanów Świerkowych w Masywie Kudłonia w Gorcach. Arch. Fotogram. Kartogr. Teledetekcji 1998, 8, 21. [Google Scholar]
- Wężyk, P.; Mucha, M.; Szostak, M. Mapa Użytkowania i Pokrycia Fragmentu Gorczańskiego Parku Narodowego Opracowana w Oparciu o Fotointerpretację Cyfrowych Ortofotomap Lotniczych CIR z 2011 Roku. Rocz. Geomatyki 2017, 15, 119–131. [Google Scholar]
- Stych, P.; Lastovicka, J.; Hladky, R.; Paluba, D. Evaluation of the Influence of Disturbances on Forest Vegetation Using the Time Series of Landsat Data: A Comparison Study of the Low Tatras and Sumava National Parks. ISPRS Int. J. Geo-Inf. 2019, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, P.; Kuemmerle, T.; Baumann, M.; Radeloff, V.C.; Abrudan, I.V.; Lieskovsky, J.; Munteanu, C.; Ostapowicz, K.; Hostert, P. Forest Disturbances, Forest Recovery, and Changes in Forest Types across the Carpathian Ecoregion from 1985 to 2010 Based on Landsat Image Composites. Remote Sens. Environ. 2014, 151, 72–88. [Google Scholar] [CrossRef]
- Bässler, C.; Seifert, L.; Müller, J. The BIOKLIM Project in the National Park Bavarian Forest: Lessons from a Biodiversity Survey. Silva Gabreta 2015, 21, 81–93. [Google Scholar]
- Tanona, M.; Czarnota, P. Natural Disturbances of the Structure of Norway Spruce Forests in Europe and Their Impact on the Preservation of Epixylic Lichen Diversity: A Review. Ecol. Quest. 2019, 30, 7–17. [Google Scholar] [CrossRef]
- Thorn, S.; Bässler, C.; Svoboda, M.; Müller, J. Effects of Natural Disturbances and Salvage Logging on Biodiversity—Lessons from the Bohemian Forest. For. Ecol. Manag. 2017, 388, 113–119. [Google Scholar] [CrossRef]
- Kulakowski, D.; Seidl, R.; Holeksa, J.; Kuuluvainen, T.; Nagel, T.A.; Panayotov, M.; Svoboda, M.; Thorn, S.; Vacchiano, G.; Whitlock, C.; et al. A Walk on the Wild Side: Disturbance Dynamics and the Conservation and Management of European Mountain Forest Ecosystems. For. Ecol. Manag. 2017, 388, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.A.; Zenner, E.K.; Brang, P. Research in Old-Growth Forests and Forest Reserves: Implications for Integrated Forest Management. In Integrative Approaches as an Opportunity for the Conservation of Forest Biodiversity; Kraus, D., Krumm, F., Eds.; European Forest Institute: Joensuu, Finland, 2013; pp. 44–50. ISBN 978-952-5980-07-3. [Google Scholar]
- Nováková, M.H.; Edwards-Jonášová, M. Restoration of Central-European Mountain Norway Spruce Forest 15 Years after Natural and Anthropogenic Disturbance. For. Ecol. Manag. 2015, 344, 120–130. [Google Scholar] [CrossRef]
- Zielonka, T. When Does Dead Wood Turn into a Substrate for Spruce Replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Małek, S.; Barszcz, J.; Majsterkiewicz, K. Changes in the Threat of Spruce Stand Disintegration in the Beskid Śla̧ski and Żywiecki Mts. in the Years 2007-2010. J. For. Sci. 2012, 58, 519–529. [Google Scholar] [CrossRef]
- Jarosz, S. Badania Geograficzno-Leśne w Gorcach. Pract. Rol.-Śne 1935, 16, 1–125. [Google Scholar]
- Chwistek, K. Historia Lasów i Leśnictwa w Gorcach. Wierchy 2002, 68, 135–160. [Google Scholar]
- Świerz-Zaleski, T. Rezerwat Leśny w Gorcach Imienia Władysława Orkana. Ochr. Przyr. 1930, 10, 54–58. [Google Scholar]
- Lang, S.; Schöpfer, E.; Langanke, T. Combined Object-Based Classification and Manual Interpretation–Synergies for a Quantitative Assessment of Parcels and Biotopes. Geocarto Int. 2009, 24, 99–114. [Google Scholar] [CrossRef]
- Blaschke, T.; Hay, G.J.; Kelly, M.; Lang, S.; Hofmann, P.; Addink, E.; Queiroz Feitosa, R.; van der Meer, F.; van der Werff, H.; van Coillie, F.; et al. Geographic Object-Based Image Analysis—Towards a New Paradigm. ISPRS J. Photogramm. Remote Sens. 2014, 87, 180–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year of Acquisition | Spectral Composition | Ground Sampling Distance [m] | Source |
|---|---|---|---|
| 1977 | B&W | 0.50 | GNP archive * |
| 1987 | B&W | 0.75 | GNP archive * |
| 1997 | CIR | 0.25 | GNP archive * |
| 2003 | B&W | 0.25 | GUGiK |
| 2009 | RGB | 0.25 | GUGiK |
| 2015 | RGB and CIR | 0.25 | GUGiK |
| 2020 | RGB and CIR | 0.05 | GNP archive |
| LC Class Code | Description |
|---|---|
| NSH | Mature, healthy forest stands dominated by the Norway spruce. |
| EBH | Healthy, forest stands dominated by the European beech. |
| MXH | Mature, healthy mixed forest stands, typically European beech- Norway spruce, European beech-silver fir, or silver fir-Norway spruce mixtures, with sycamore (Acer pseudoplatanus L.) and rowan (Sorbus aucuparia L.) admixtures. |
| NSD | Areas with standing, dead and severely damaged groups of Norway spruce trees. |
| DEF | Deforested areas with fallen deadwood, forest gaps. |
| REG | Young forests on post-forest decline areas. |
| LC Class | 1977–1984 | 1984–1987 | 1987–1991 | 1991–1997 | 1997–2003 | 2003–2009 | 2009–2015 | 2015–2020 | Total Changes Compared to 1977 |
|---|---|---|---|---|---|---|---|---|---|
| [ha] | |||||||||
| EBH | * | * | * | * | * | * | * | −0.02 | −0.02 |
| MXH | * | * | * | −2.18 | −0.11 | −11.23 | 0.60 | 0.66 | −12.26 |
| NSH | −6.68 | −29.20 | −16.16 | −8.70 | −11.00 | −88.87 | −18.33 | −6.26 | −185.21 |
| NSD | 4.98 | 14.14 | 6.73 | −19.16 | −1.93 | 80.27 | −62.13 | −14.90 | 8.01 |
| REG | ** | ** | ** | 16.88 | 18.52 | 31.19 | 56.99 | 24.86 | 148.43 |
| DEF | 1.70 | 15.06 | 9.43 | 13.16 | −5.49 | −11.35 | 22.87 | −4.35 | 41.04 |
| LC Class | 1977–1984 | 1984–1987 | 1987–1991 | 1991–1997 | 1997–2003 | 2003–2009 | 2009–2015 | 2015–2020 | Total Changes Compared to 1977 |
|---|---|---|---|---|---|---|---|---|---|
| [ha] | |||||||||
| EBH | * | * | * | * | * | * | * | * | 0.0 |
| MXH | 0.0 | 0.0 | 0.0 | −1.44 | −0.79 | −22.25 | 0.68 | 1.44 | −22.35 |
| NSH | −7.53 | −10.07 | −16.09 | −33.93 | −3.02 | −111.05 | −89.85 | −58.20 | −329.74 |
| NSD | 7.26 | 6.96 | 6.12 | −1.85 | −6.52 | 104.01 | −53.59 | −15.14 | 47.25 |
| REG | ** | ** | ** | 18.31 | 3.28 | 25.10 | 49.70 | 33.55 | 129.94 |
| DEF | 0.27 | 3.14 | 9.97 | 18.92 | 7.04 | 4.20 | 93.07 | 38.34 | 174.95 |
| 1977–1984 | 1984–1987 | 1987–1991 | 1991–1997 | 1997–2003 | 2003–2009 | 2009–2015 | 2015–2020 | |
|---|---|---|---|---|---|---|---|---|
| [%] | ||||||||
| Subarea A— Kudłoń | 1.86 | 8.13 | 4.50 | 8.36 | 5.15 | 31.01 | 22.39 | 7.10 |
| Subarea B— Jaworzyna | 1.64 | 2.20 | 3.51 | 8.11 | 2.25 | 29.05 | 31.26 | 15.98 |
| EBH | MXH | NSH | NSD | REG | DEF | |
|---|---|---|---|---|---|---|
| [%] | ||||||
| Subarea A— Kudłoń | 0.00 | 2.05 | 25.77 | 28.41 | 20.65 | 11.61 |
| Subarea B— Jaworzyna | 0.00 | 2.90 | 35.93 | 21.95 | 14.16 | 19.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krawczyk, W.; Wężyk, P. Using Satellite Imagery and Aerial Orthophotos for the Multi-Decade Monitoring of Subalpine Norway Spruce Stands Changes in Gorce National Park, Poland. Remote Sens. 2023, 15, 951. https://doi.org/10.3390/rs15040951
Krawczyk W, Wężyk P. Using Satellite Imagery and Aerial Orthophotos for the Multi-Decade Monitoring of Subalpine Norway Spruce Stands Changes in Gorce National Park, Poland. Remote Sensing. 2023; 15(4):951. https://doi.org/10.3390/rs15040951
Chicago/Turabian StyleKrawczyk, Wojciech, and Piotr Wężyk. 2023. "Using Satellite Imagery and Aerial Orthophotos for the Multi-Decade Monitoring of Subalpine Norway Spruce Stands Changes in Gorce National Park, Poland" Remote Sensing 15, no. 4: 951. https://doi.org/10.3390/rs15040951