
Distance to a River Modifies Climate Legacy on Vegetation Growth in a Boreal Riparian Forest
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
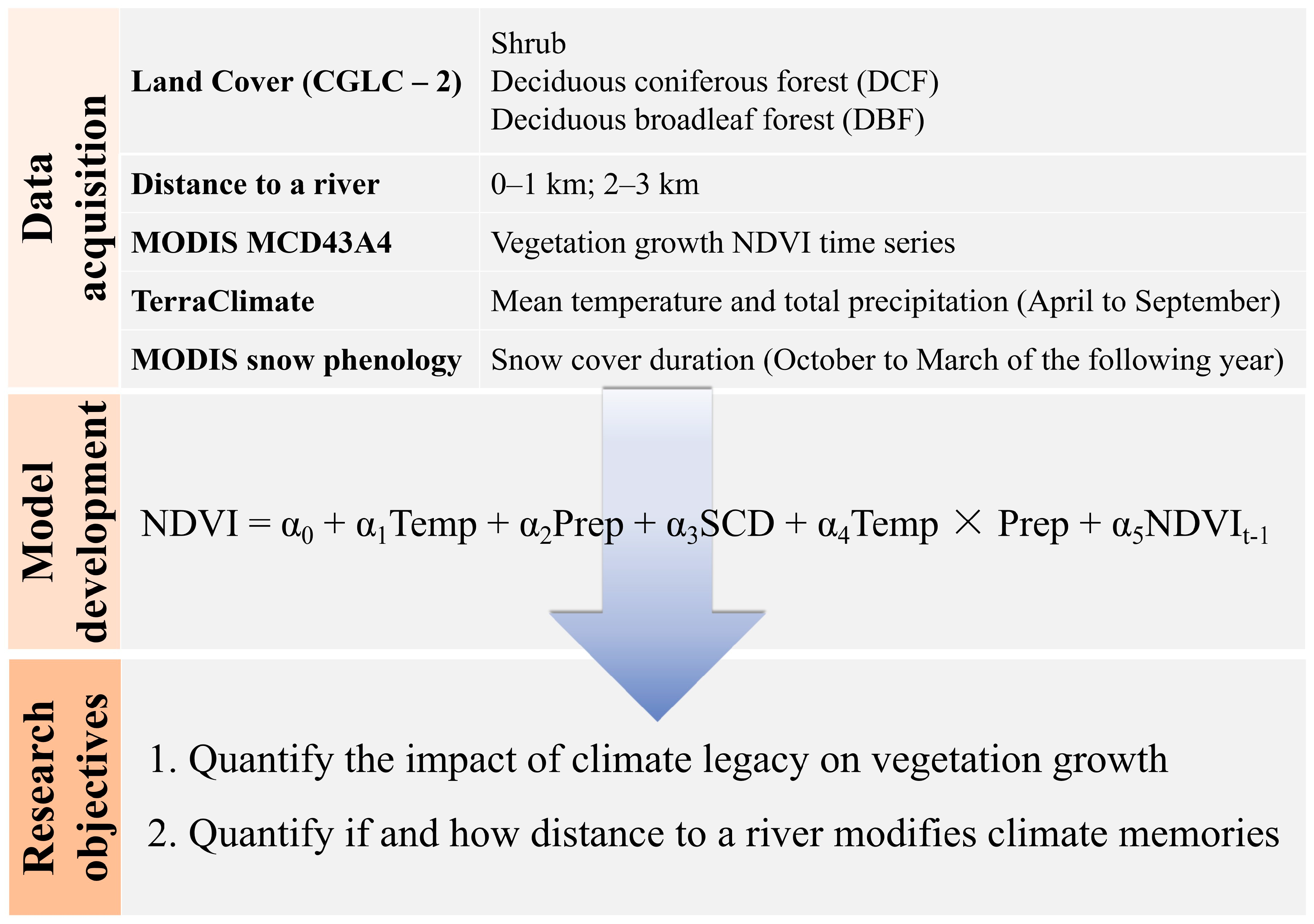
2. Materials and Methods
2.1. Study Area
2.2. Random Site Selection
2.3. MODIS Derived NDVI Time Series
2.4. Climate Data
2.5. Methodology
3. Results
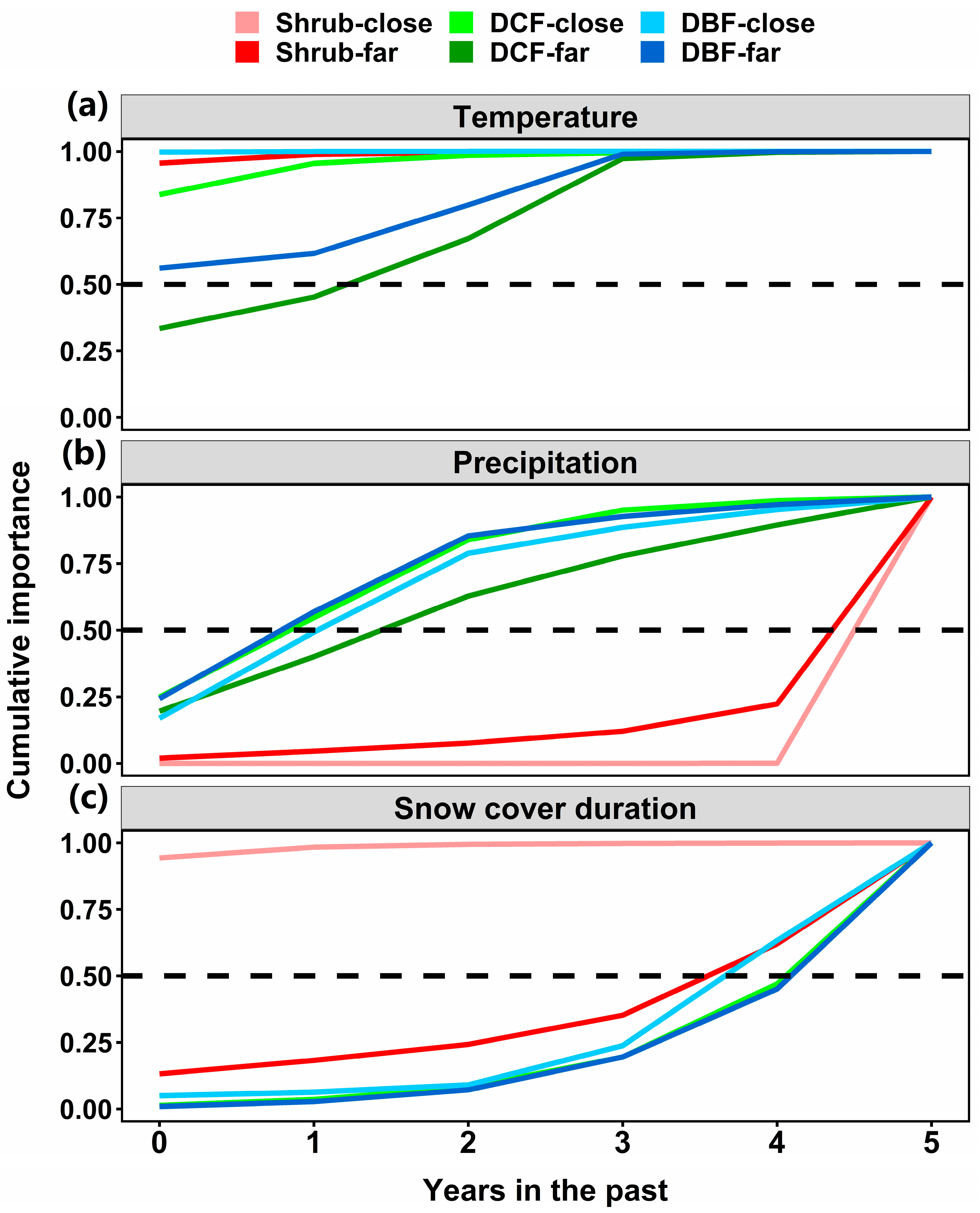
3.1. Vegetation Growth in Response to Temperature, Precipitation and Snow Cover Duration
3.2. Distance to a River Modifies the Importance and Duration of Climatic Legacy
4. Discussion
4.1. Effects of Concurrent and Antecedent Temperature and Precipitation on Vegetation Growth
4.2. Snow Cover Duration Legacy on Vegetation Growth
4.3. River Modulates Climate Legacy
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar] [CrossRef]
- Frelich, L.E.; Montgomery, R.A.; Reich, P.B. Seven Ways a Warming Climate Can Kill the Southern Boreal Forest. Forests 2021, 12, 560. [Google Scholar] [CrossRef]
- Boulanger, Y.; Taylor, A.R.; Price, D.T.; Cyr, D.; McGarrigle, E.; Rammer, W.; Sainte-Marie, G.; Beaudoin, A.; Guindon, L.; Mansuy, N. Climate change impacts on forest landscapes along the Canadian southern boreal forest transition zone. Landsc. Ecol. 2017, 32, 1415–1431. [Google Scholar] [CrossRef]
- Sun, J.; Liu, W.; Pan, Q.; Zhang, B.; Lv, Y.; Huang, J.; Han, X. Positive legacies of severe droughts in the Inner Mongolia grassland. Sci. Adv. 2022, 8, eadd6249. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Wang, L.; Smith, W.K.; Chang, Q.; Wang, H.; D’Odorico, P. Observed increasing water constraint on vegetation growth over the last three decades. Nat. Commun. 2021, 12, 3777. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Bermudez, R.; Montgomery, R.A.; Rich, R.L.; Rice, K.E.; Hobbie, S.E.; Stefanski, A. Even modest climate change may lead to major transitions in boreal forests. Nature 2022, 608, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Bayle, A.; Roy, A.; Dedieu, J.-P.; Boudreau, S.; Choler, P.; Lévesque, E. Two distinct waves of greening in northeastern Canada: Summer warming does not tell the whole story. Environ. Res. Lett. 2022, 17, 064051. [Google Scholar] [CrossRef]
- Li, Z.; Fang, H.; Tu, J.; Li, X.; Sha, Z. Phenological Shifts of the Deciduous Forests and Their Responses to Climate Variations in North America. Forests 2022, 13, 1137. [Google Scholar] [CrossRef]
- Park, T.; Ganguly, S.; Tømmervik, H.; Euskirchen, E.S.; Høgda, K.-A.; Karlsen, S.R.; Brovkin, V.; Nemani, R.R.; Myneni, R.B. Changes in growing season duration and productivity of northern vegetation inferred from long-term remote sensing data. Environ. Res. Lett. 2016, 11, 084001. [Google Scholar] [CrossRef]
- Peltier, D.M.P.; Anderegg, W.R.L.; Guo, J.S.; Ogle, K. Contemporary tree growth shows altered climate memory. Ecol. Lett. 2022, 25, 2663–2674. [Google Scholar] [CrossRef]
- Huang, C.; Liang, Y.; He, H.S.; Wu, M.M.; Liu, B.; Ma, T. Sensitivity of aboveground biomass and species composition to climate change in boreal forests of Northeastern China. Ecol. Model. 2021, 445, 109472. [Google Scholar] [CrossRef]
- Buermann, W.; Forkel, M.; O’Sullivan, M.; Sitch, S.; Friedlingstein, P.; Haverd, V.; Jain, A.K.; Kato, E.; Kautz, M.; Lienert, S.; et al. Widespread seasonal compensation effects of spring warming on northern plant productivity. Nature 2018, 562, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, A.D.; Myers-Smith, I.H.; Elmendorf, S.C.; Normand, S.; Rüger, N.; Beck, P.S.A.; Blach-Overgaard, A.; Blok, D.; Cornelissen, J.H.C.; Forbes, B.C.; et al. Plant functional trait change across a warming tundra biome. Nature 2018, 562, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Manzoni, S.; Vico, G.; Bastos, A.; de Vries, F.T.; Messori, G. Drought Legacy in Sub-Seasonal Vegetation State and Sensitivity to Climate Over the Northern Hemisphere. Geophys. Res. Lett. 2022, 49, e2022GL098700. [Google Scholar] [CrossRef]
- Nanzad, L.; Zhang, J.; Tuvdendorj, B.; Yang, S.; Rinzin, S.; Prodhan, F.A.; Sharma, T.P.P. Assessment of Drought Impact on Net Primary Productivity in the Terrestrial Ecosystems of Mongolia from 2003 to 2018. Remote Sens. 2021, 13, 2522. [Google Scholar] [CrossRef]
- Sturm, M.; Schimel, J.; Michaelson, G.; Welker, J.M.; Oberbauer, S.F.; Liston, G.E.; Fahnestock, J.; Romanovsky, V.E. Winter Biological Processes Could Help Convert Arctic Tundra to Shrubland. BioScience 2005, 55, 17–26. [Google Scholar] [CrossRef]
- Monson, R.K.; Lipson, D.L.; Burns, S.P.; Turnipseed, A.A.; Delany, A.C.; Williams, M.W.; Schmidt, S.K. Winter forest soil respiration controlled by climate and microbial community composition. Nature 2006, 439, 711–714. [Google Scholar] [CrossRef]
- Ellebjerg, S.M.; Tamstorf, M.P.; Illeris, L.; Michelsen, A.; Hansen, B.U. Inter-Annual Variability and Controls of Plant Phenology and Productivity at Zackenberg. Adv. Ecol. Res. 2008, 40, 249–273. [Google Scholar]
- Pedersen, S.H.; Liston, G.E.; Tamstorf, M.P.; Abermann, J.; Lund, M.; Schmidt, N.M. Quantifying snow controls on vegetation greenness. Ecosphere 2018, 9, e02309. [Google Scholar] [CrossRef]
- Shakhmatov, R.; Hashiguchi, S.; Maximov, T.C.; Sugimoto, A. Effects of snow manipulation on larch trees in the taiga forest ecosystem in northeastern Siberia. Prog. Earth Planet. Sci. 2022, 9, 3. [Google Scholar] [CrossRef]
- Pulliainen, J.; Luojus, K.; Derksen, C.; Mudryk, L.; Lemmetyinen, J.; Salminen, M.; Ikonen, J.; Takala, M.; Cohen, J.; Smolander, T.; et al. Patterns and trends of Northern Hemisphere snow mass from 1980 to 2018. Nature 2020, 581, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Lievens, H.; Demuzere, M.; Marshall, H.-P.; Reichle, R.H.; Brucker, L.; Brangers, I.; de Rosnay, P.; Dumont, M.; Girotto, M.; Immerzeel, W.W.; et al. Snow depth variability in the Northern Hemisphere mountains observed from space. Nat. Commun. 2019, 10, 4629. [Google Scholar] [CrossRef] [PubMed]
- Cooper, E.J. Warmer Shorter Winters Disrupt Arctic Terrestrial Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 271–295. [Google Scholar] [CrossRef]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Change Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [PubMed]
- Vilonen, L.; Ross, M.; Smith, M.D. What happens after drought ends: Synthesizing terms and definitions. New Phytol. 2022, 235, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Stampfli, A.; Zeiter, M. The impact of seed deficiency on productivity and on negative drought effect in semi-natural grassland. J. Veg. Sci. 2020, 31, 1066–1078. [Google Scholar] [CrossRef]
- Guo, J.S.; Gear, L.; Hultine, K.R.; Koch, G.W.; Ogle, K. Non-structural carbohydrate dynamics associated with antecedent stem water potential and air temperature in a dominant desert shrub. Plant Cell Environ. 2020, 43, 1467–1483. [Google Scholar] [CrossRef]
- Zeng, Z.; Wu, W.; Ge, Q.; Li, Z.; Wang, X.; Zhou, Y.; Zhang, Z.; Li, Y.; Huang, H.; Liu, G.; et al. Legacy effects of spring phenology on vegetation growth under preseason meteorological drought in the Northern Hemisphere. Agric. For. Meteorol. 2021, 310, 108630. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, X.; Wang, Y.; Bai, Y. How precipitation legacies affect broad-scale patterns of primary productivity: Evidence from the Inner Mongolia grassland. Agric. For. Meteorol. 2022, 320, 108954. [Google Scholar] [CrossRef]
- Mackie, K.A.; Zeiter, M.; Bloor, J.M.G.; Stampfli, A. Plant functional groups mediate drought resistance and recovery in a multisite grassland experiment. J. Ecol. 2019, 107, 937–949. [Google Scholar] [CrossRef]
- Stampfli, A.; Bloor, J.M.G.; Fischer, M.; Zeiter, M. High land-use intensity exacerbates shifts in grassland vegetation composition after severe experimental drought. Glob. Change Biol. 2018, 24, 2021–2034. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Schwalm, C.R.; Anderegg, W.R.L. Ghosts of the past: How drought legacy effects shape forest functioning and carbon cycling. Ecol. Lett. 2020, 23, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Itter, M.S.; D’Orangeville, L.; Dawson, A.; Kneeshaw, D.; Duchesne, L.; Finley, A.O.; Battipaglia, G. Boreal tree growth exhibits decadal-scale ecological memory to drought and insect defoliation, but no negative response to their interaction. J. Ecol. 2018, 107, 1288–1301. [Google Scholar] [CrossRef]
- Ren, Y.; Zhang, L.; Yang, K.; Li, Z.; Yin, R.; Tan, B.; Wang, L.; Liu, Y.; Li, H.; You, C.; et al. Short-term effects of snow cover manipulation on soil bacterial diversity and community composition. Sci. Total Environ. 2020, 741, 140454. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, Y.; Yang, K.; Li, Z.; Tan, B.; Liu, Y.; Li, H.; You, C.; Liu, S.; Wang, L.; et al. Immediate and legacy effects of snow exclusion on soil fungal diversity and community composition. For. Ecosyst. 2021, 8, 22. [Google Scholar] [CrossRef]
- Blume-Werry, G.; Kreyling, J.; Laudon, H.; Milbau, A.; Gilliam, F. Short-term climate change manipulation effects do not scale up to long-term legacies: Effects of an absent snow cover on boreal forest plants. J. Ecol. 2016, 104, 1638–1648. [Google Scholar] [CrossRef]
- Nguyen, U.; Glenn, E.P.; Nagler, P.L.; Scott, R.L. Long-term decrease in satellite vegetation indices in response to environmental variables in an iconic desert riparian ecosystem: The Upper San Pedro, Arizona, United States. Ecohydrology 2015, 8, 610–625. [Google Scholar] [CrossRef]
- Perry, L.G.; Andersen, D.C.; Reynolds, L.V.; Nelson, S.M.; Shafroth, P.B. Vulnerability of riparian ecosystems to elevated CO2 and climate change in arid and semiarid western North America. Glob. Change Biol. 2012, 18, 821–842. [Google Scholar] [CrossRef]
- Henriques, M.; McVicar, T.R.; Holland, K.L.; Daly, E. Riparian vegetation and geomorphological interactions in anabranching rivers: A global review. Ecohydrology 2021, 15, e2370. [Google Scholar] [CrossRef]
- Zhu, X.; Yuan, G.; Yi, X.; Du, T. Quantifying the impacts of river hydrology on riparian vegetation spatial structure: Case study in the lower basin of the Tarim River, China. Ecohydrology 2017, 10, e1887. [Google Scholar] [CrossRef]
- Costa, F.R.C.; Schietti, J.; Stark, S.C.; Smith, M.N. The other side of tropical forest drought: Do shallow water table regions of Amazonia act as large-scale hydrological refugia from drought? New Phytol. 2023, 237, 714–733. [Google Scholar] [CrossRef] [PubMed]
- Esteban, E.J.L.; Castilho, C.V.; Melgaco, K.L.; Costa, F.R.C. The other side of droughts: Wet extremes and topography as buffers of negative drought effects in an Amazonian forest. New Phytol. 2021, 229, 1995–2006. [Google Scholar] [CrossRef] [PubMed]
- He, R.-X.; Jin, H.-J.; Luo, D.-L.; Li, X.-Y.; Zhou, C.-F.; Jia, N.; Jin, X.-Y.; Li, X.-Y.; Che, T.; Yang, X.; et al. Permafrost changes in the Nanwenghe Wetlands Reserve on the southern slope of the Da Xing’anling–Yile’huli mountains, Northeast China. Adv. Clim. Change Res. 2021, 12, 696–709. [Google Scholar] [CrossRef]
- Buchhorn, M.; Lesiv, M.; Tsendbazar, N.-E.; Herold, M.; Bertels, L.; Smets, B. Copernicus Global Land Cover Layers—Collection 2. Remote Sens. 2020, 12, 1044. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- Andreadis, K.M.; Schumann, G.J.P.; Pavelsky, T.M.J.W.R.R. A simple global river bankfull width and depth database. Water Resour. Res. 2013, 49, 7164–7168. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Int. J. Remote Sens. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Forkel, M.; Carvalhais, N.; Verbesselt, J.; Mahecha, M.D.; Neigh, C.S.R.; Reichstein, M. Trend Change Detection in NDVI Time Series: Effects of Inter-Annual Variability and Methodology. Remote Sens. 2013, 5, 2113–2144. [Google Scholar] [CrossRef]
- Schaaf, C.B.; Gao, F.; Strahler, A.H.; Lucht, W.; Li, X.; Tsang, T.; Strugnell, N.C.; Zhang, X.; Jin, Y.; Muller, J.-P.; et al. First operational BRDF, albedo nadir reflectance products from MODIS. Remote Sens. Environ. 2002, 83, 135–148. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Treydte, K.; Zhang, Z.; Kang, H.; Zeng, X.; Xu, G.; Wu, Q.; Kang, S. Permafrost degradation alters the environmental signals recorded in tree-ring lignin methoxy group delta(2)H in northeastern China. Sci. Total Environ. 2023, 860, 160519. [Google Scholar] [CrossRef]
- Zhao, Q.; Hao, X.; Wang, J.; Sun, X.; Li, H. A Dataset of Snow Phenology in China Based on MODIS from 2000 to 2020. Investig. Snow Cover. Charact. Distrib. China 2022, 7, 3. [Google Scholar] [CrossRef]
- Ogle, K.; Barber, J.J.; Barron-Gafford, G.A.; Bentley, L.P.; Young, J.M.; Huxman, T.E.; Loik, M.E.; Tissue, D.T. Quantifying ecological memory in plant and ecosystem processes. Ecol. Lett. 2015, 18, 221–235. [Google Scholar] [CrossRef]
- Itter, M.S.; Vanhatalo, J.; Finley, A.O. EcoMem: An R package for quantifying ecological memory. Environ. Model. Softw. 2019, 119, 305–308. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 16 April 2022).
- Hoover, D.L.; Pfennigwerth, A.A.; Duniway, M.C. Drought resistance and resilience: The role of soil moisture–plant interactions and legacies in a dryland ecosystem. J. Ecol. 2021, 109, 3280–3294. [Google Scholar] [CrossRef]
- Muller, L.M.; Bahn, M. Drought legacies and ecosystem responses to subsequent drought. Glob. Change Biol. 2022, 28, 5086–5103. [Google Scholar] [CrossRef] [PubMed]
- Price, D.T.; Alfaro, R.I.; Brown, K.J.; Flannigan, M.D.; Fleming, R.A.; Hogg, E.H.; Girardin, M.P.; Lakusta, T.; Johnston, M.; McKenney, D.W.; et al. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 2013, 21, 322–365. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S.; et al. Detection and attribution of vegetation greening trend in China over the last 30 years. Glob. Change Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef]
- Liu, X.; Tian, Y.; Liu, S.; Jiang, L.; Mao, J.; Jia, X.; Zha, T.; Zhang, K.; Wu, Y.; Zhou, J. Time-Lag Effect of Climate Conditions on Vegetation Productivity in a Temperate Forest–Grassland Ecotone. Forests 2022, 13, 1024. [Google Scholar] [CrossRef]
- Lian, X.; Jiao, L.; Liu, Z.; Jia, Q.; Zhong, J.; Fang, M.; Wang, W. Multi-spatiotemporal heterogeneous legacy effects of climate on terrestrial vegetation dynamics in China. GISci. Remote Sens. 2022, 59, 164–183. [Google Scholar] [CrossRef]
- Li, C.; Wang, R.; Cui, X.; Wu, F.; Yan, Y.; Peng, Q.; Qian, Z.; Xu, Y. Responses of vegetation spring phenology to climatic factors in Xinjiang, China. Ecol. Indic. 2021, 124, 107286. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophys. Res. Lett. 2006, 33, L23402. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Chen, A.; Wang, K.; Li, X.; Buermann, W.; Huntingford, C.; Penuelas, J.; Xu, H.; Myneni, R.B. Seasonal biological carryover dominates northern vegetation growth. Nat. Commun. 2021, 12, 983. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Kong, D.; Shi, P.; Singh, V.P.; Sun, P. Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Shen, M.; Piao, S.; Chen, X.; An, S.; Fu, Y.H.; Wang, S.; Cong, N.; Janssens, I.A. Strong impacts of daily minimum temperature on the green-up date and summer greenness of the Tibetan Plateau. Glob. Change Biol. 2016, 22, 3057–3066. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, W.; Xue, K.; Wang, S.; Zhang, L.; Hu, R.; Zeng, H.; Xu, X.; Li, Y.; Jiang, L.; et al. Grassland changes and adaptive management on the Qinghai–Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 668–683. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J. Mediterranean dwarf shrubs and coexisting trees present different radial-growth synchronies and responses to climate. Plant Ecol. 2012, 213, 1687–1698. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Sánchez-Salguero, R.; Vicente-Serrano, S.M.; Serra-Maluquer, X.; Gutiérrez, E.; Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; et al. Drought legacies are short, prevail in dry conifer forests and depend on growth variability. J. Ecol. 2020, 108, 2473–2484. [Google Scholar] [CrossRef]
- Li, P.; Zhu, D.; Wang, Y.; Liu, D. Elevation dependence of drought legacy effects on vegetation greenness over the Tibetan Plateau. Agric. For. Meteorol. 2020, 295, 108190. [Google Scholar] [CrossRef]
- Wu, X.; Liu, H.; Li, X.; Ciais, P.; Babst, F.; Guo, W.; Zhang, C.; Magliulo, V.; Pavelka, M.; Liu, S.; et al. Differentiating drought legacy effects on vegetation growth over the temperate Northern Hemisphere. Glob. Change Biol. 2018, 24, 504–516. [Google Scholar] [CrossRef]
- Dong, B.; Yu, Y.; Pereira, P. Non-growing season drought legacy effects on vegetation growth in southwestern China. Sci. Total Environ. 2022, 846, 157334. [Google Scholar] [CrossRef]
- Jiang, P.; Liu, H.; Piao, S.; Ciais, P.; Wu, X.; Yin, Y.; Wang, H. Enhanced growth after extreme wetness compensates for post-drought carbon loss in dry forests. Nat. Commun. 2019, 10, 195. [Google Scholar] [CrossRef]
- Grippa, M.; Kergoat, L.; Le Toan, T.; Mognard, N.M.; Delbart, N.; L’Hermitte, J.; Vicente-Serrano, S.M. The impact of snow depth and snowmelt on the vegetation variability over central Siberia. Geophys. Res. Lett. 2005, 32, L21412. [Google Scholar] [CrossRef]
- Zhang, X.; Manzanedo, R.D.; D’Orangeville, L.; Rademacher, T.T.; Li, J.; Bai, X.; Hou, M.; Chen, Z.; Zou, F.; Song, F.; et al. Snowmelt and early to mid-growing season water availability augment tree growth during rapid warming in southern Asian boreal forests. Glob. Change Biol. 2019, 25, 3462–3471. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, K.C.; Pedersen, S.H.; Leffler, A.J.; Sexton, J.O.; Feng, M.; Welker, J.M. Winter snow and spring temperature have differential effects on vegetation phenology and productivity across Arctic plant communities. Glob. Change Biol. 2021, 27, 1572–1586. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tang, Y.; Xu, J.; Shu, S.; Yu, B.; Wu, J.; Huang, Y. Impact of Snow Cover Phenology on the Vegetation Green-Up Date on the Tibetan Plateau. Remote Sens. 2022, 14, 3909. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Fang, J.; Wang, X. Change in winter snow depth and its impacts on vegetation in China. Glob. Change Biol. 2010, 16, 3004–3013. [Google Scholar] [CrossRef]
- Allen, S.T.; Kirchner, J.W.; Braun, S.; Siegwolf, R.T.W.; Goldsmith, G.R. Seasonal origins of soil water used by trees. Hydrol. Earth Syst. Sci. 2019, 23, 1199–1210. [Google Scholar] [CrossRef]
- Li, Y.; Wu, X.; Huang, Y.; Li, X.; Shi, F.; Zhao, S.; Yang, Y.; Tian, Y.; Wang, P.; Zhang, S.; et al. Compensation effect of winter snow on larch growth in Northeast China. Clim. Chang. 2021, 164, 54. [Google Scholar] [CrossRef]
- Shamir, E.; Meko, D.; Touchan, R.; Lepley, K.S.; Campbell, R.; Kaliff, R.N.; Georgakakos, K.P. Snowpack- and Soil Water Content-Related Hydrologic Indices and Their Association with Radial Growth of Conifers in the Sierra Nevada, California. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005331. [Google Scholar] [CrossRef]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D.; Tierney, G.L. Colder soils in a warmer world: A snow manipulation study in a northern hardwood forest ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Wipf, S. Phenology, growth, and fecundity of eight subarctic tundra species in response to snowmelt manipulations. Plant Ecol. 2010, 207, 53–66. [Google Scholar] [CrossRef]
- García-Cervigón Morales, A.I.; Olano Mendoza, J.M.; Eugenio Gozalbo, M.; Camarero Martínez, J.J. Arboreal and prostrate conifers coexisting in Mediterranean high mountains differ in their climatic responses. Dendrochronologia 2012, 30, 279–286. [Google Scholar] [CrossRef]
- Dolores Bejarano, M.; Nilsson, C.; GonzáLez Del Tánago, M.; Marchamalo, M. Responses of riparian trees and shrubs to flow regulation along a boreal stream in northern Sweden. Freshw. Biol. 2011, 56, 853–866. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Sun, Q.; Zou, H.; Marschner, P. Distance to a River Modifies Climate Legacy on Vegetation Growth in a Boreal Riparian Forest. Remote Sens. 2023, 15, 5582. https://doi.org/10.3390/rs15235582
Li Y, Sun Q, Zou H, Marschner P. Distance to a River Modifies Climate Legacy on Vegetation Growth in a Boreal Riparian Forest. Remote Sensing. 2023; 15(23):5582. https://doi.org/10.3390/rs15235582
Chicago/Turabian StyleLi, Yingyu, Qiaoqi Sun, Hongfei Zou, and Petra Marschner. 2023. "Distance to a River Modifies Climate Legacy on Vegetation Growth in a Boreal Riparian Forest" Remote Sensing 15, no. 23: 5582. https://doi.org/10.3390/rs15235582