


Combining Hyperspectral, LiDAR, and Forestry Data to Characterize Riparian Forests along Age and Hydrological Gradients
 ,
,
Abstract
:1. Introduction
- (1)
- Explore changing biophysical characteristics along age gradients.
- (2)
- Explore changing biophysical characteristics between river reaches with differing geomorphic features and hydrological connectivity.
- (3)
- Assess the use of random forest classifiers to predict forest connectivity in riparian forests.
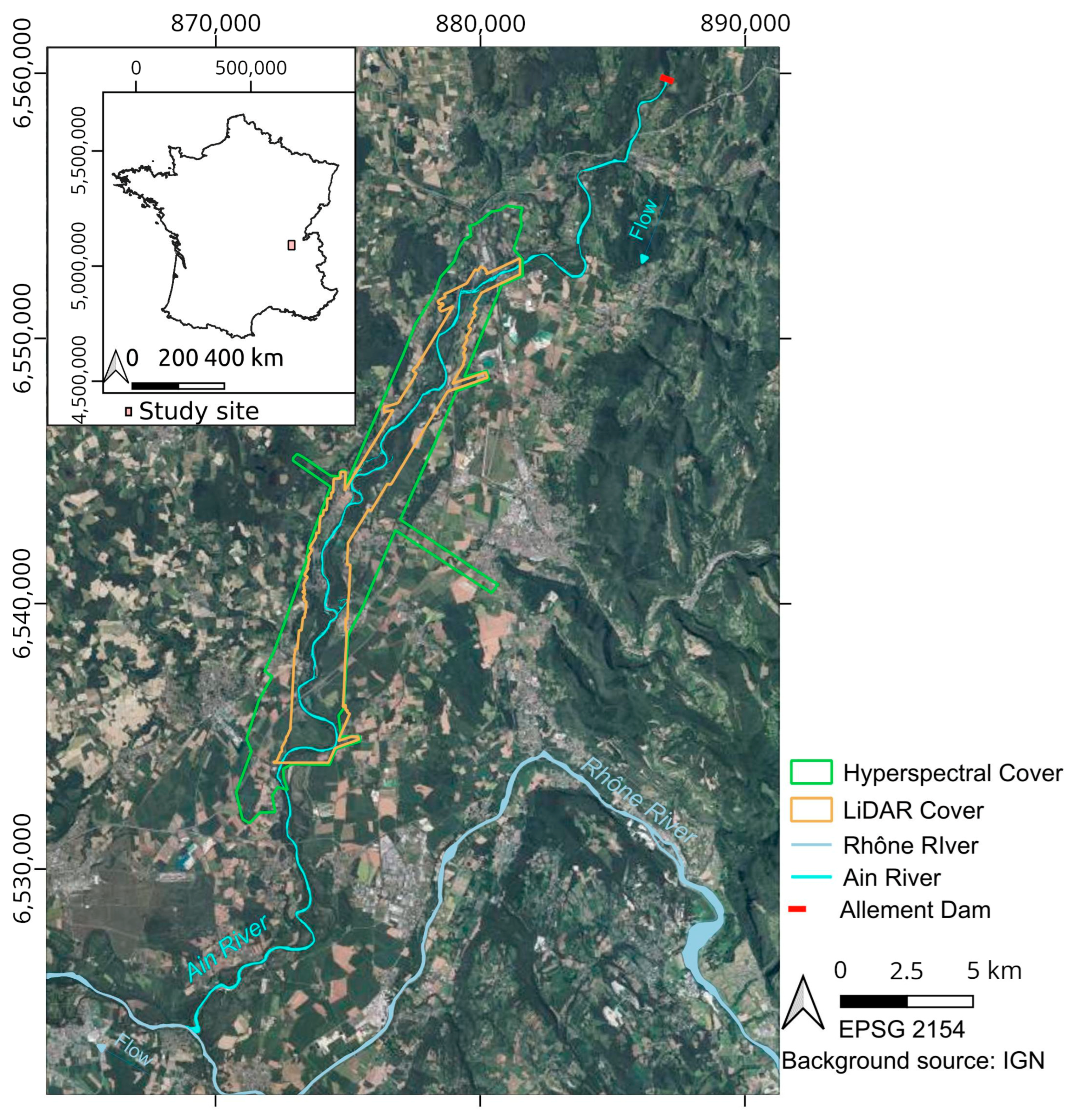
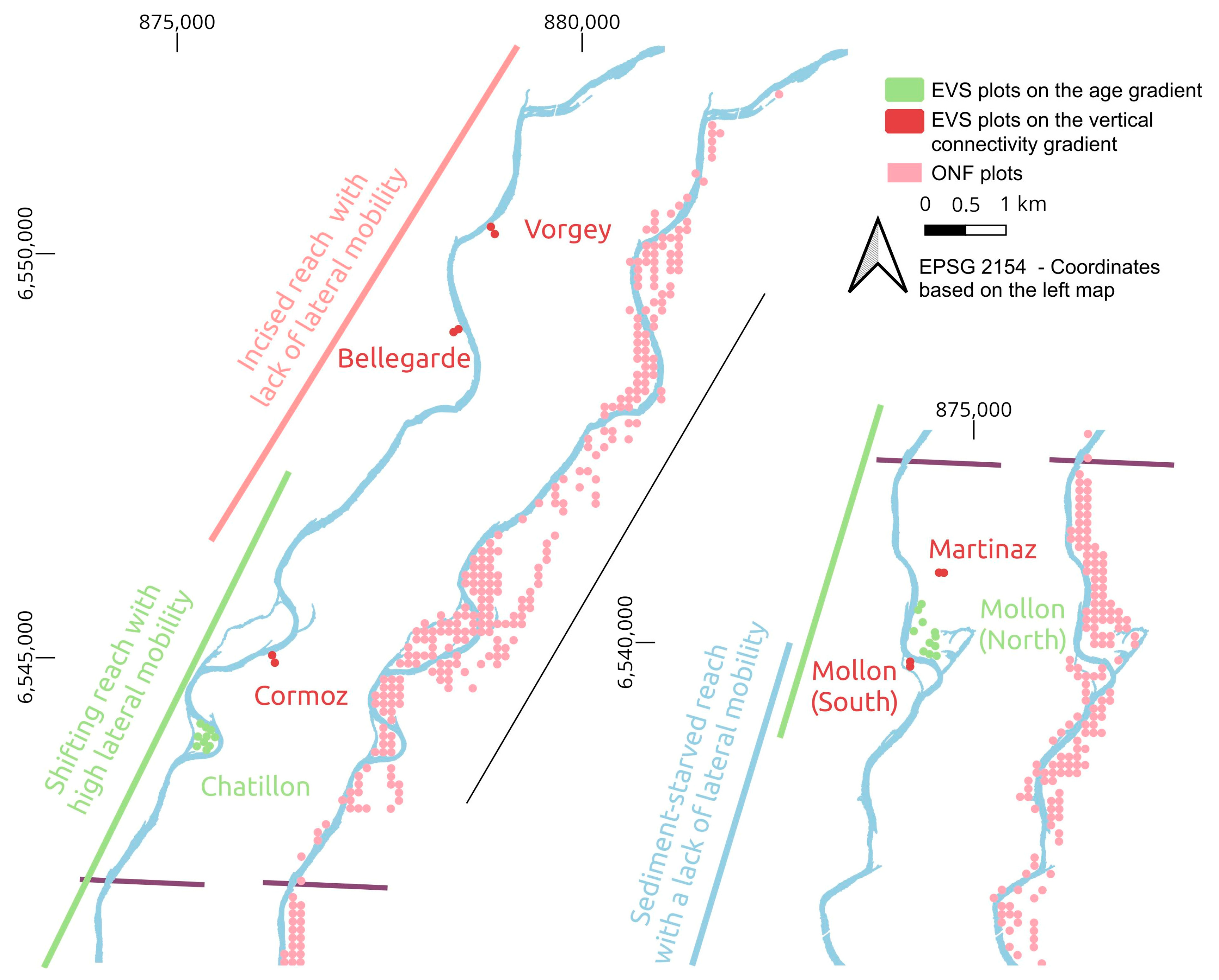
2. Study Site
3. Materials
3.1. Remote Sensing Information
3.1.1. Hyperspectral Imagery
3.1.2. LiDAR Data
3.1.3. Series of Aerial Photos Since the 1940s
3.2. Field Calibration Data
3.2.1. Vegetation Survey during the Airborne Campaign in 2015
3.2.2. The Extensive Vegetation Surveys Performed in 2008 and 2017 by ONF
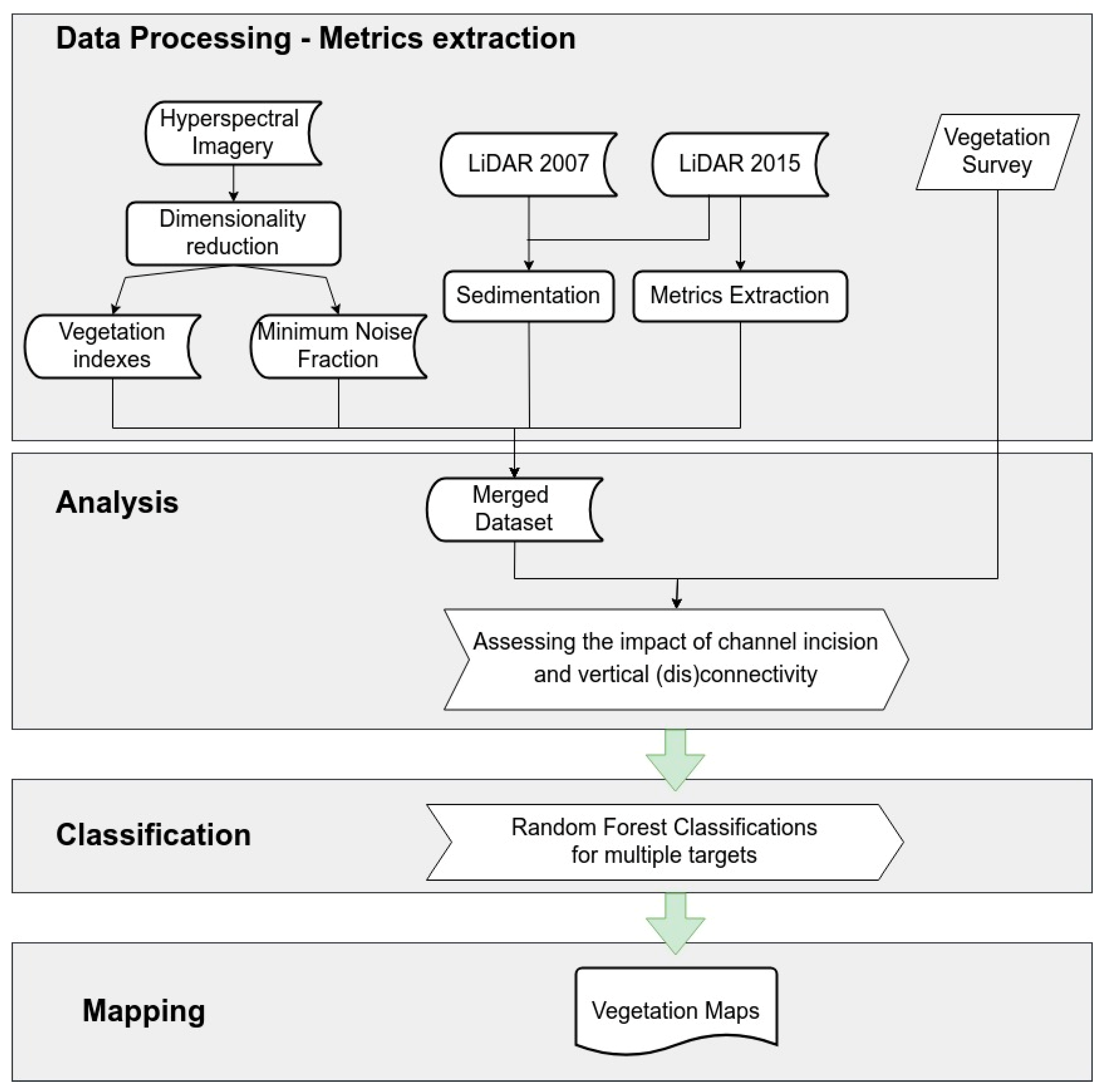
4. Methods
4.1. Data Processing: Extracting Forest Indicators from Hyperspectral and LiDAR Data
4.2. Data Analysis: Studying the Riparian Forest by Assessing the Impact of Channel Incision and Vertical (Dis) Connection to the River System
4.3. Random Forest Classifications of Forest Connectivity and Resulting Maps
5. Results
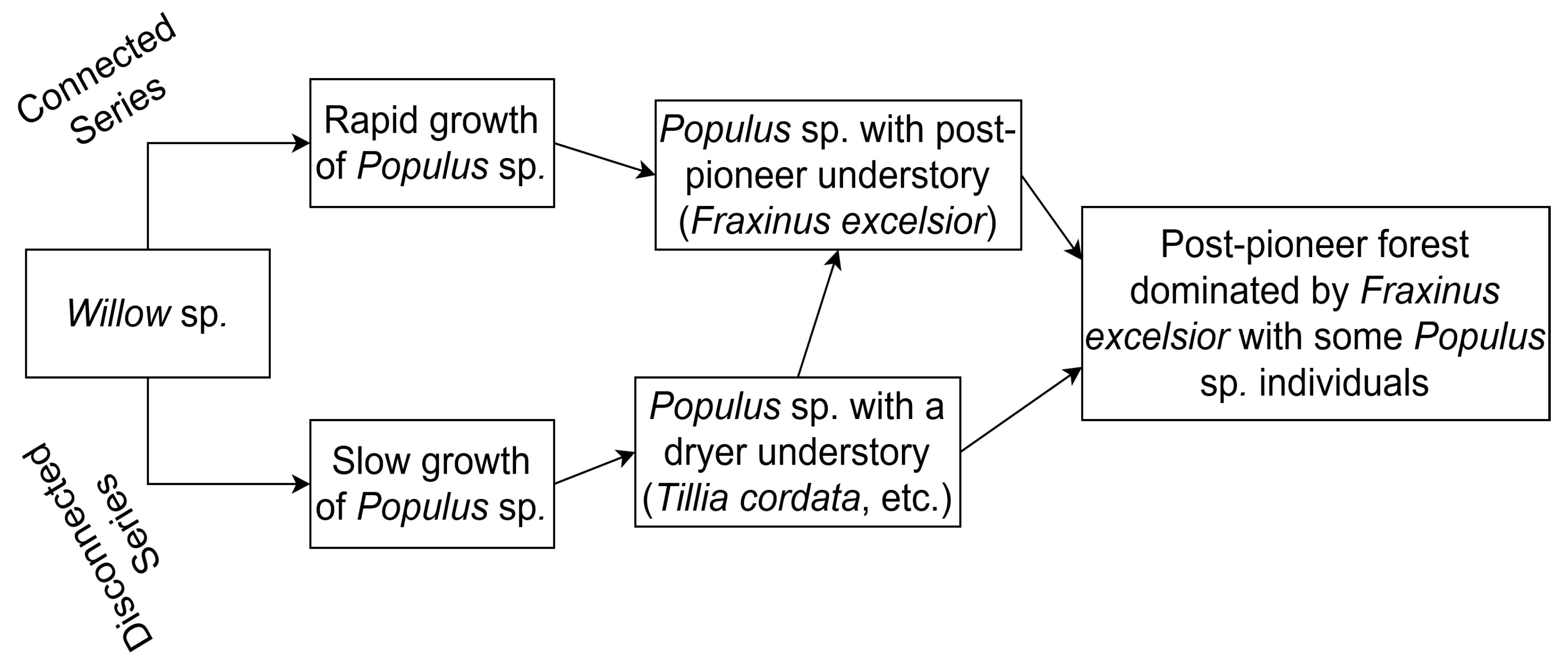
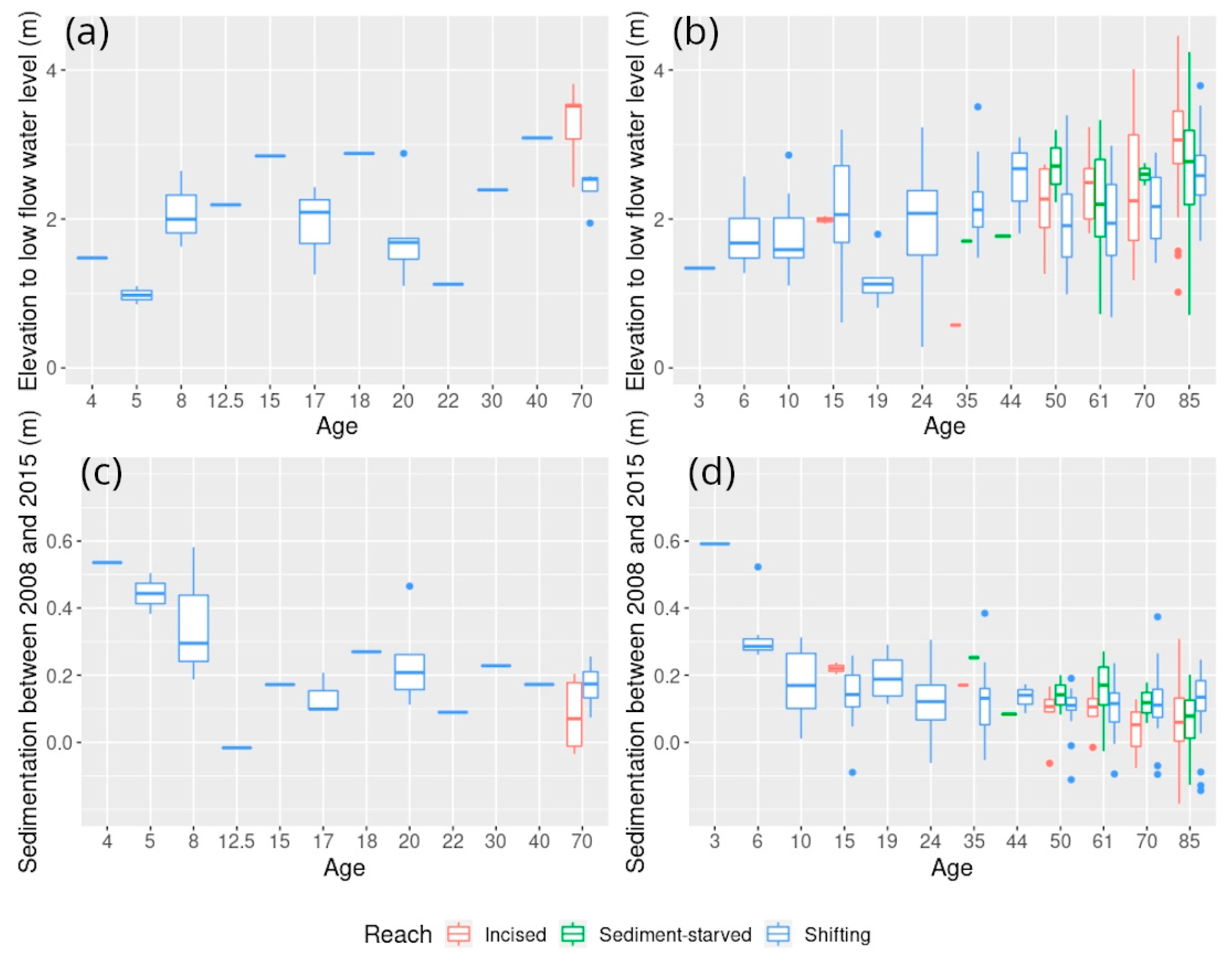
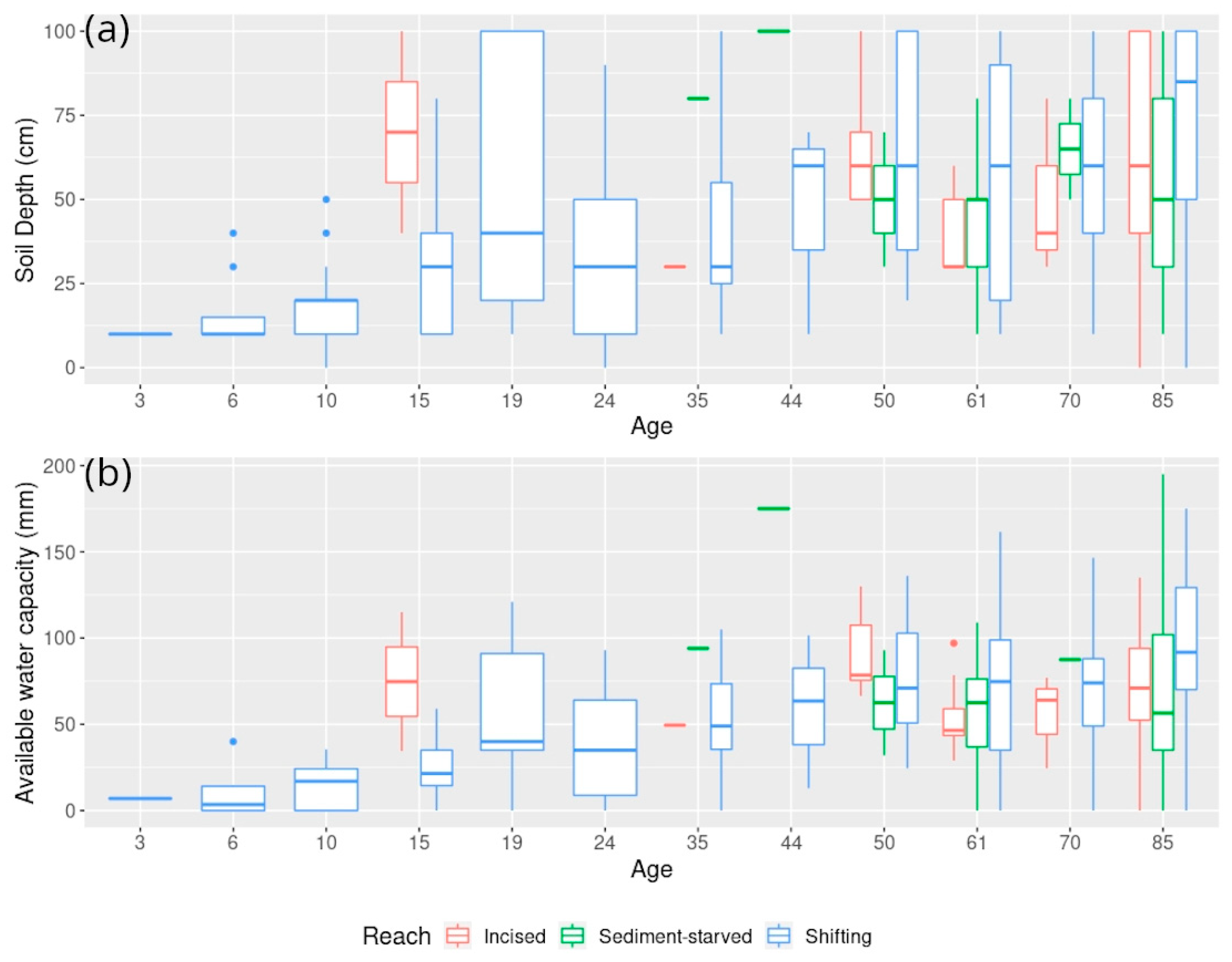
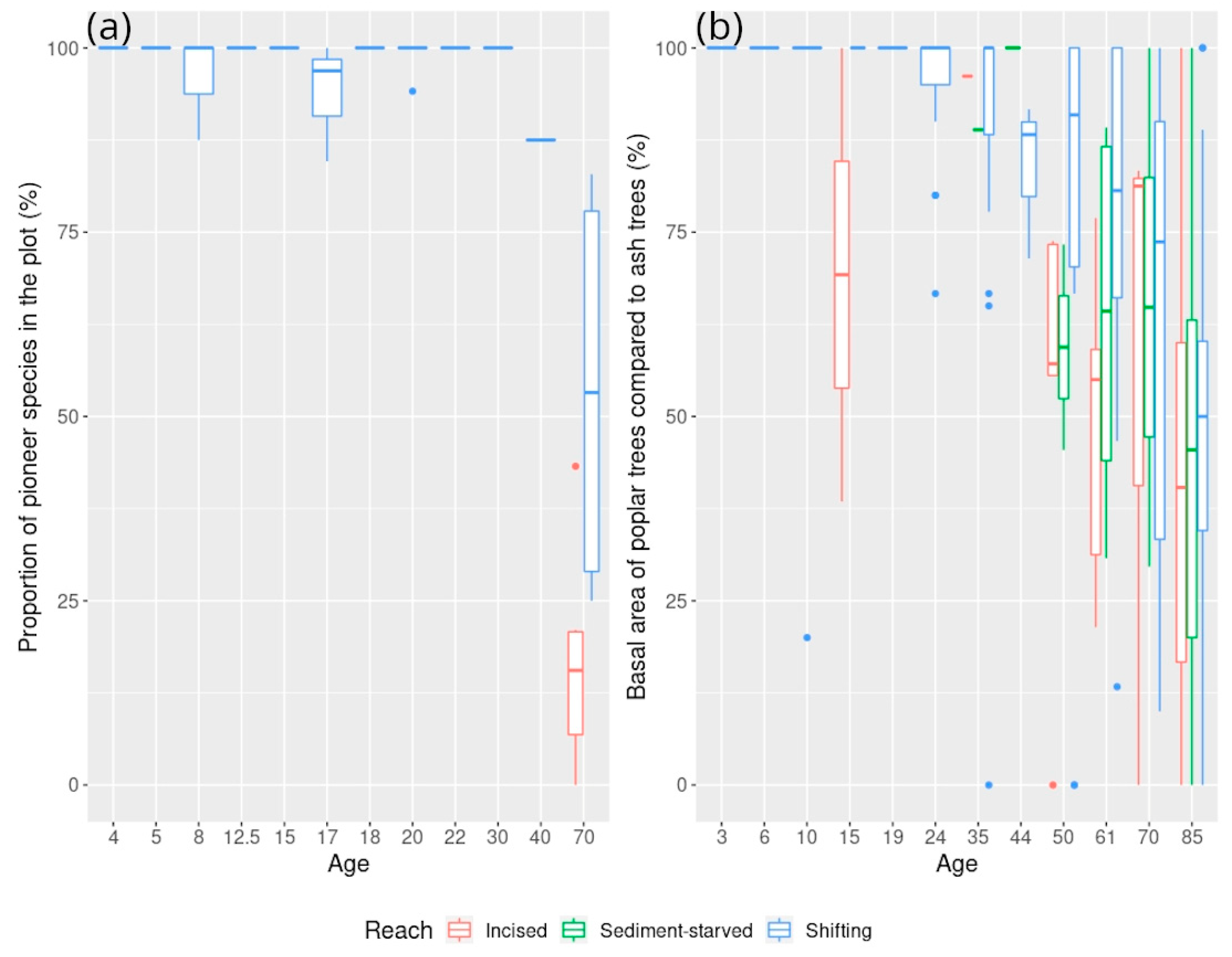
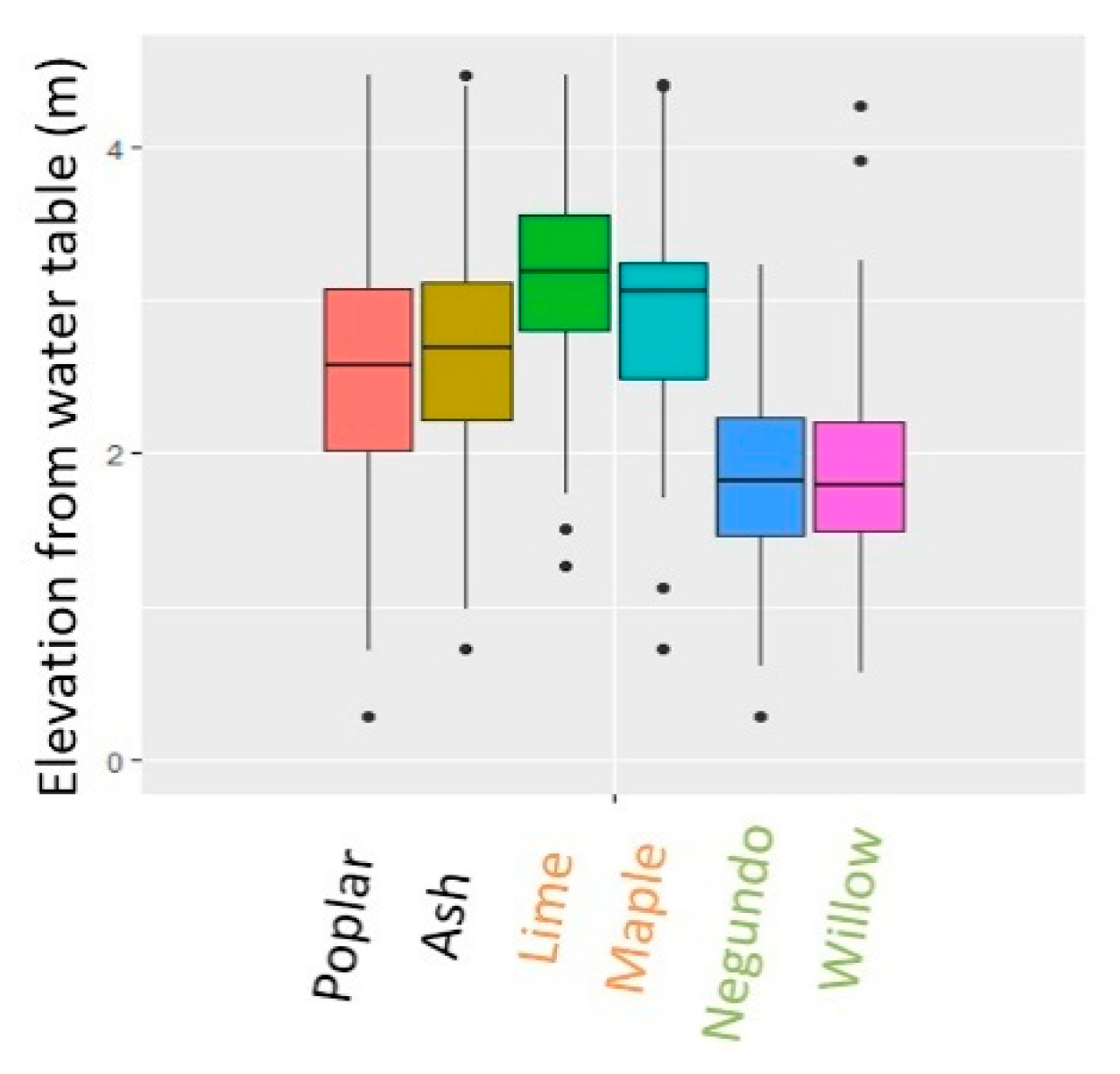
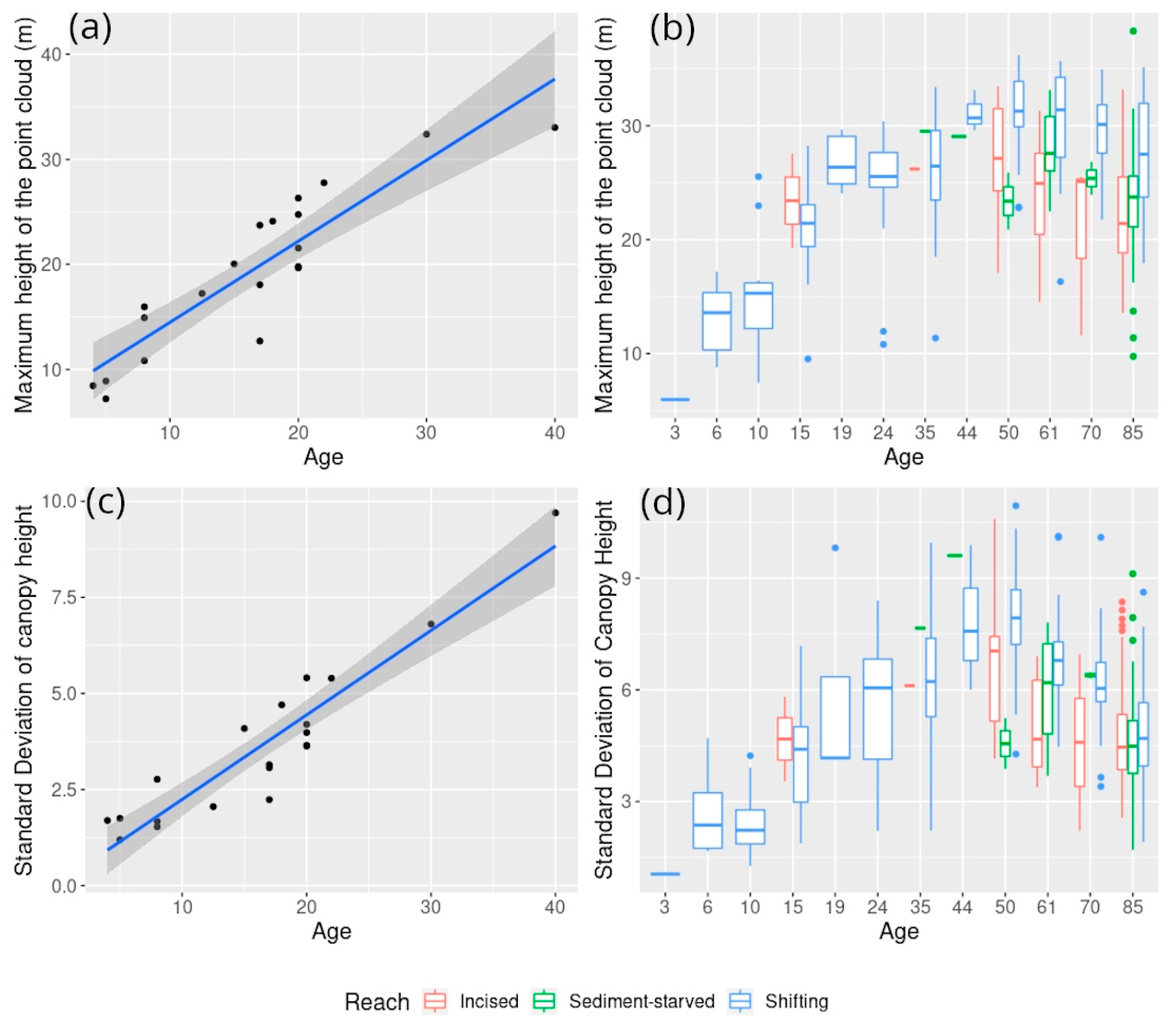
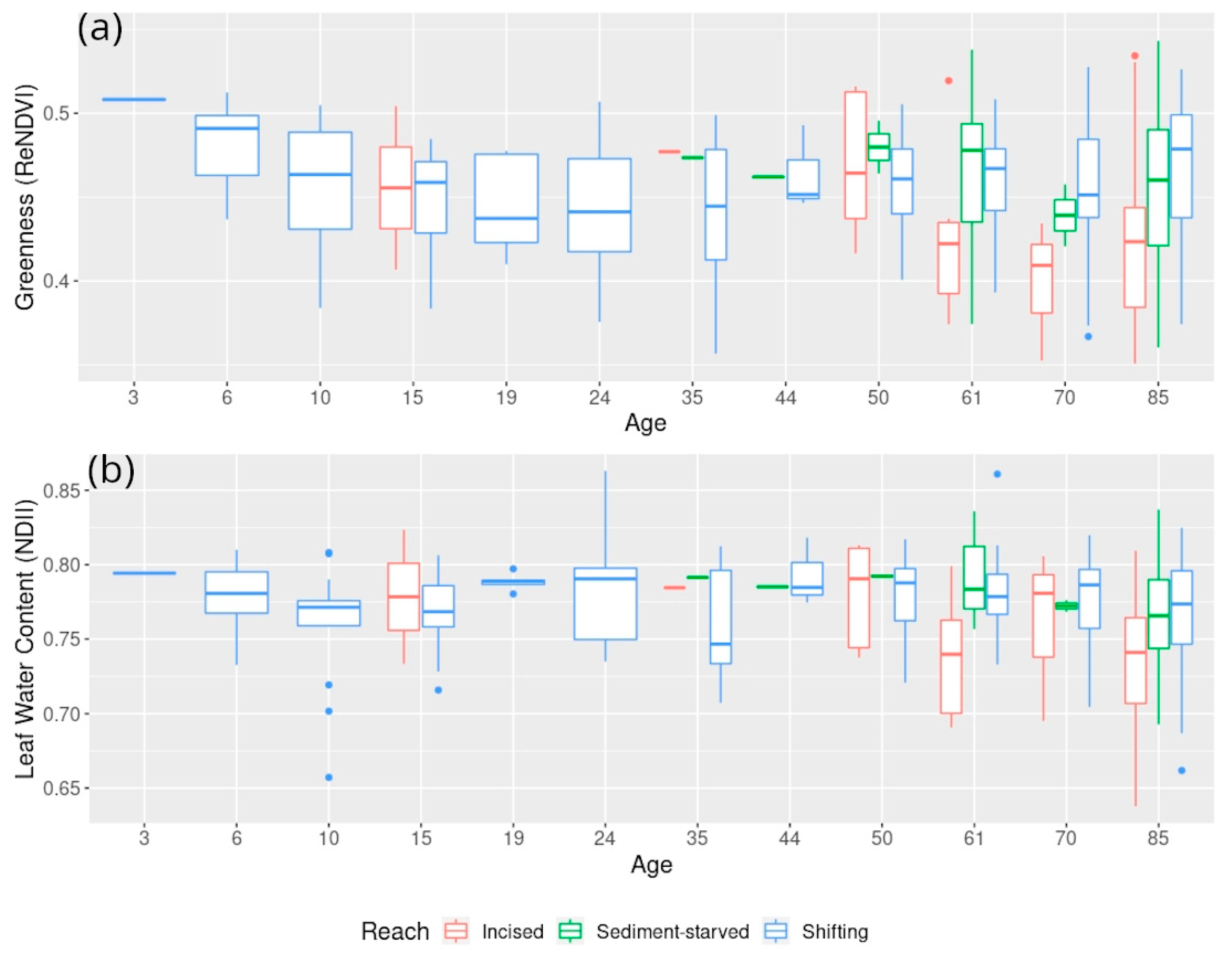
5.1. Exploring Forest Characteristics and Their Evolution along the Age Gradient at Varying Degrees of Hydrological Connectivity
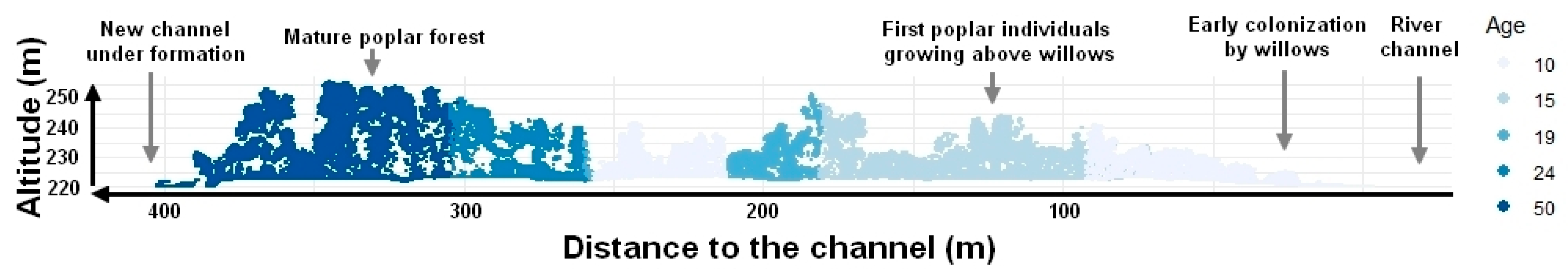
5.1.1. Characterization of Hydrological and Sedimentological Changes
5.1.2. Associated Changes in Species Composition According to Field Surveys
5.1.3. Associated Changes in Forest Structure and Reflectance
5.2. Can We Predict and Map the Shift in Forest Composition, Structure, and Reflectance That Results from Vertical (Dis)connection of the Riparian Forest Due to Channel Incision?
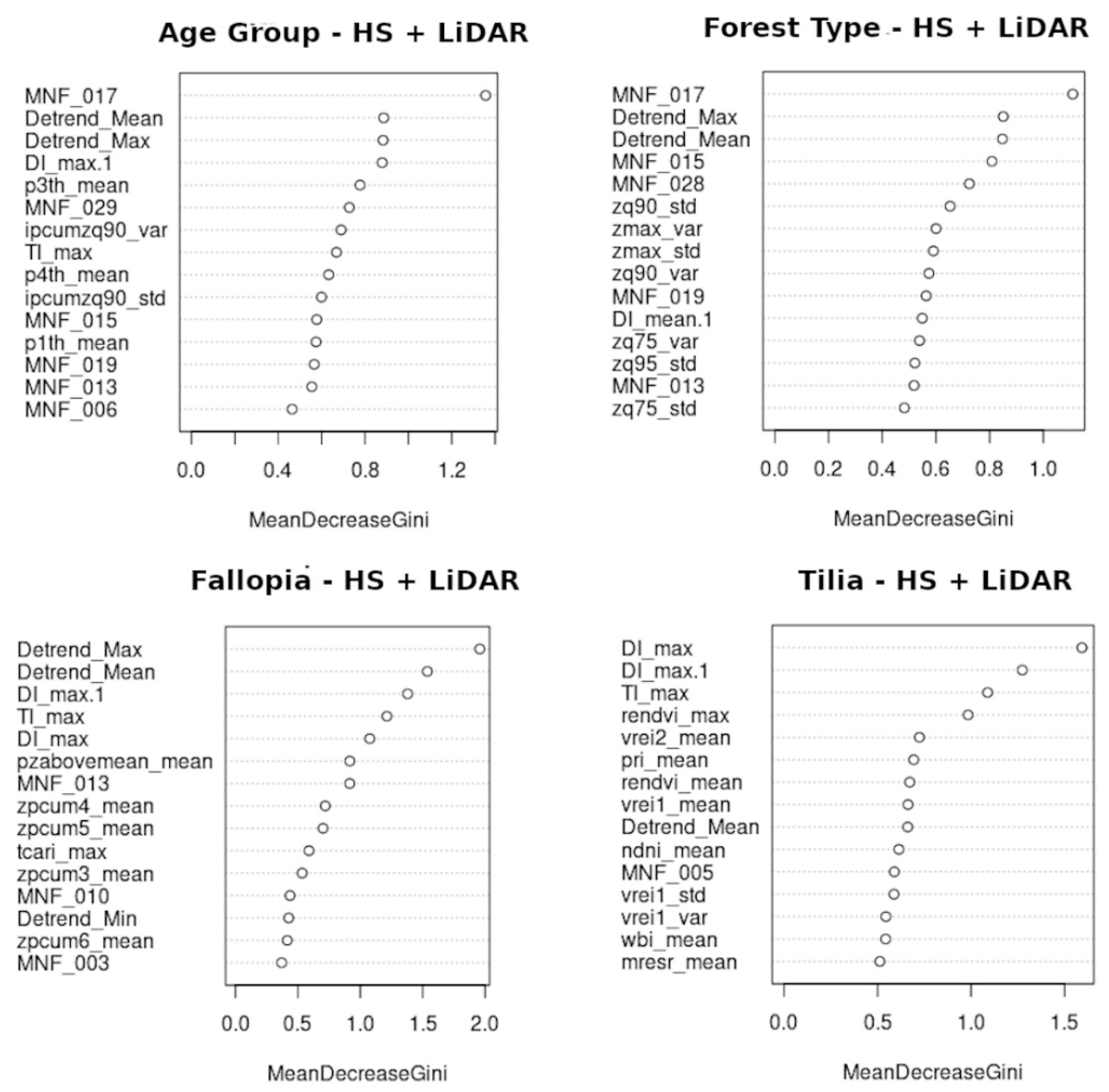
5.2.1. Random Forest Classifications
5.2.2. Mapping Indicators of Riparian Forest Connectivity across the Lower Ain River
6. Discussion
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. List of Indexes and Metrics Extracted from the LiDAR and Hyperspectral Data and Their Abbreviations
Appendix A.1. Narrowband Hyperspectral Indexes
| 1 | MSI |
| 2 | NMDI |
| 3 | WBI |
| 4 | NDWI |
| 5 | NDII |
| 6 | CAI |
| 7 | LCAI |
| 8 | PSRI |
| 9 | PRI |
| 10 | MCARI |
| 11 | MRENDVI |
| 12 | MRESR |
| 13 | MTVI |
| 14 | MTVI2 |
| 15 | RENDVI |
| 16 | TCARI |
| 17 | TVI |
| 18 | VREI1 |
| 19 | VREI2 |
| 20 | ARI1 |
| 21 | ARI2 |
| 22 | CRI1 |
| 23 | CRI2 |
| 24 | NDLI |
| 25 | NDNI |
Appendix A.2. Topographic Indexes Derived from LiDAR Data
| 1 | Elevation relative to low-flow water level (Detrend) |
| 2 | Catchment area (CA) |
| 3 | Catchment slope (CS) |
| 4 | Modified catchment area (MCA) |
| 5 | Topographic wetness index (TWI) |
| 6 | Multiresolution index of ridge top flatness (MRRTF) |
| 7 | Multiresolution index of valley bottom flatness (MRVBF) |
| 8 | Direct insolation (DI) |
| 9 | Diffuse insolation (DI.1) |
| 10 | Total insolation (TI) |
| 11 | Duration of insolation (DoI) |
| 12 | Topographic position index (TPI) |
Appendix A.3. Structural Indexes Derived from LiDAR Data
| 1 | Maximum height (zmax) |
| 2 | Mean height (zmean) |
| 3 | Entropy of height distribution (zentropy) |
| 4 | Percentage of returns above zmean (pzabovemean) |
| 5 | 5th percentile of height distribution (zq5) |
| 6 | 10th percentile of height distribution (zq10) |
| 7 | 15th percentile of height distribution (zq15) |
| 8 | 20th percentile of height distribution (zq20) |
| 9 | 25th percentile of height distribution (zq25) |
| 10 | 30th percentile of height distribution (zq30) |
| 11 | 35th percentile of height distribution (zq35) |
| 12 | 40th percentile of height distribution (zq40) |
| 13 | 45th percentile of height distribution (zq45) |
| 14 | 50th percentile of height distribution (zq50) |
| 15 | 55th percentile of height distribution (zq55) |
| 16 | 60th percentile of height distribution (zq60) |
| 17 | 65th percentile of height distribution (zq65) |
| 18 | 70th percentile of height distribution (zq70) |
| 19 | 75th percentile of height distribution (zq75) |
| 20 | 80th percentile of height distribution (zq80) |
| 21 | 85th percentile of height distribution (zq85) |
| 22 | 90th percentile of height distribution (zq90) |
| 23 | 95th percentile of height distribution (zq95) |
| 24 | Cumulative percentage of returns in the 1st layer (zpcum1) |
| 25 | Cumulative percentage of returns in the 2nd layer (zpcum2) |
| 26 | Cumulative percentage of returns in the 3rd layer (zpcum3) |
| 27 | Cumulative percentage of returns in the 4th layer (zpcum4) |
| 28 | Cumulative percentage of returns in the 5th layer (zpcum5) |
| 29 | Cumulative percentage of returns in the 6th layer (zpcum6) |
| 30 | Cumulative percentage of returns in the 7h layer (zpcum7) |
| 31 | Cumulative percentage of returns in the 8th layer (zpcum8) |
| 32 | Cumulative percentage of returns in the 9th layer (zpcum9) |
| 33 | Total intensity (itot) |
| 34 | Max intensity (imax) |
| 35 | Mean intensity (imean) |
| 36 | Percentage of intensity returned by points classified as ground (ipground) |
| 37 | Percentage of intensity returned below the 10th percentile (ipcumzq10) |
| 38 | Percentage of intensity returned below the 30th percentile ipcumzq30 |
| 39 | Percentage of intensity returned below the 50th percentile ipcumzq50 |
| 40 | Percentage of intensity returned below the 70th percentile ipcumzq70 |
| 41 | Percentage of intensity returned below the 90th percentile ipcumzq90 |
| 42 | Percentage of intensity returned by 1st returns (p1th) |
| 43 | Percentage of intensity returned by 2nd returns (p2th) |
| 44 | Percentage of intensity returned by 3rd returns (p3th) |
| 45 | Percentage of intensity returned by 4th returns (p4th) |
| 46 | Percentage of intensity returned by 5th returns (p5th) |
| 47 | Percentage of returns classified as ground per square meter (pground) |
| 48 | Points per square meter (n) |
References
- Naiman, R.J.; Decamps, H.; Pastor, J.; Johnston, C. The Potential Importance of Boundaries to Fluvial Ecosystems. J. N. Am. Benthol. Soc. 1988, 7, 289–306. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H.; Pollock, M. The Role of Riparian Corridors in Maintaining Regional Biodiversity. Ecol. Appl. A Publ. Ecol. Soc. Am. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Riis, T.; Kelly-Quinn, M.; Aguiar, F.C.; Manolaki, P.; Bruno, D.; Bejarano, M.D.; Clerici, N.; Fernandes, M.R.; Franco, J.C.; Pettit, N.; et al. Global Overview of Ecosystem Services Provided by Riparian Vegetation. BioScience 2020, 70, 501–514. [Google Scholar] [CrossRef]
- Poole, G.C.; Berman, C.H. An Ecological Perspective on In-Stream Temperature: Natural Heat Dynamics and Mechanisms of Human-CausedThermal Degradation. Environ. Manag. 2001, 27, 787–802. [Google Scholar] [CrossRef]
- Roth, T.R.; Westhoff, M.C.; Huwald, H.; Huff, J.A.; Rubin, J.F.; Barrenetxea, G.; Vetterli, M.; Parriaux, A.; Selker, J.S.; Parlange, M.B. Stream Temperature Response to Three Riparian Vegetation Scenarios by Use of a Distributed Temperature Validated Model. Environ. Sci. Technol. 2010, 44, 2072–2078. [Google Scholar] [CrossRef] [PubMed]
- Wondzell, S.M.; Diabat, M.; Haggerty, R. What Matters Most: Are Future Stream Temperatures More Sensitive to Changing Air Temperatures, Discharge, or Riparian Vegetation? J. Am. Water Resour. Assoc. 2019, 55, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Dosskey, M.G.; Vidon, P.; Gurwick, N.P.; Allan, C.J.; Duval, T.P.; Lowrance, R. The Role of Riparian Vegetation in Protecting and Improving Chemical Water Quality in Streams1. J. Am. Water Resour. Assoc. 2010, 46, 261–277. [Google Scholar] [CrossRef]
- Tabacchi, E.; Lambs, L.; Guilloy, H.; Planty-Tabacchi, A.-M.; Muller, E.; Décamps, H. Impacts of Riparian Vegetation on Hydrological Processes. Hydrol. Process. 2000, 14, 2959–2976. [Google Scholar] [CrossRef]
- Décamps, H. How a Riparian Landscape Finds Form and Comes Alive. Landsc. Urban Plan. 2001, 57, 169–175. [Google Scholar] [CrossRef]
- González del Tánago, M.; Martínez-Fernández, V.; Aguiar, F.C.; Bertoldi, W.; Dufour, S.; García de Jalón, D.; Garófano-Gómez, V.; Mandzukovski, D.; Rodríguez-González, P.M. Improving River Hydromorphological Assessment through Better Integration of Riparian Vegetation: Scientific Evidence and Guidelines. J. Environ. Manag. 2021, 292, 112730. [Google Scholar] [CrossRef]
- Francis, R.A.; Gurnell, A.M.; Petts, G.E.; Edwards, P.J. Survival and Growth Responses of Populus nigra, Salix elaeagnos and Alnus incana Cuttings to Varying Levels of Hydric Stress. For. Ecol. Manag. 2005, 210, 291–301. [Google Scholar] [CrossRef]
- Bravard, J.-P.; Amoros, C.; Pautou, G.; Bornette, G.; Bournaud, M.; Creuzé des Châtelliers, M.; Gibert, J.; Peiry, J.-L.; Perrin, J.-F.; Tachet, H. River Incision in South-East France: Morphological Phenomena and Ecological Effects. Regul. Rivers: Res. Manag. 1997, 13, 75–90. [Google Scholar] [CrossRef]
- Comiti, F.; Da Canal, M.; Surian, N.; Mao, L.; Picco, L.; Lenzi, M.A. Channel Adjustments and Vegetation Cover Dynamics in a Large Gravel Bed River over the Last 200 years. Geomorphology 2011, 125, 147–159. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Merritt, D.M.; Pepin, D.M. Homogenization of Regional River Dynamics by Dams and Global Biodiversity Implications. Proc. Natl. Acad. Sci. USA 2007, 104, 5732–5737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, M.L.; Lines, G.C.; Auble, G.T. Channel Incision and Patterns of Cottonwood Stress and Mortality along the Mojave River, California. J. Arid Environ. 2000, 44, 399–414. [Google Scholar] [CrossRef]
- Décamps, H.; Fortuné, M.; Gazelle, F.; Pautou, G. Historical Influence of Man on the Riparian Dynamics of a Fluvial Landscape. Landsc. Ecol. 1988, 1, 163–173. [Google Scholar] [CrossRef] [Green Version]
- O’Briain, R. Climate Change and European Rivers: An Eco-Hydromorphological Perspective. Ecohydrology 2019, 12, e2099. [Google Scholar] [CrossRef]
- Rivaes, R.; Rodríguez-González, P.M.; Albuquerque, A.; Pinheiro, A.N.; Egger, G.; Ferreira, M.T. Riparian Vegetation Responses to Altered Flow Regimes Driven by Climate Change in Mediterranean Rivers. Ecohydrology 2013, 6, 413–424. [Google Scholar] [CrossRef]
- Stella, J.C.; Rodríguez-González, P.M.; Dufour, S.; Bendix, J. Riparian Vegetation Research in Mediterranean-Climate Regions: Common Patterns, Ecological Processes, and Considerations for Management. Hydrobiologia 2013, 719, 291–315. [Google Scholar] [CrossRef]
- Pautou, G.; Girel, J. La Végétation de La Basse Plaine de l’Ain: Organisation Spatiale et Évolution. In Recherches Interdisciplinaires sur Les Écosystèmes de la Basse Plaine de l’Ain (France): Potentialités Évolutives et Gestion; Documents de Cartographie Ecologique; Laboratoire de biologie végétale: Grenoble, France, 1986; pp. 75–96. [Google Scholar]
- Planty-Tabacchi, A.-M.; Tabacchi, E.; Naiman, R.J.; Deferrari, C.; Décamps, H. Invasibility of Species-Rich Communities in Riparian Zones. Conserv. Biol. 1996, 10, 598–607. [Google Scholar] [CrossRef]
- Tabacchi, E.; Planty-Tabacchi, A.-M.; Salinas, M.J.; Décamps, H. Landscape Structure and Diversity in Riparian Plant Communities: A Longitudinal Comparative Study. Regul. Rivers Res. Manag. 1996, 12, 367–390. [Google Scholar] [CrossRef]
- Carbonneau, P.; Fonstad, M.A.; Marcus, W.A.; Dugdale, S.J. Making Riverscapes Real. Geomorphology 2012, 137, 74–86. [Google Scholar] [CrossRef]
- Carbonneau, P.E.; Piégay, H. Introduction: The Growing Use of Imagery in Fundamental and Applied River Sciences. In Fluvial Remote Sensing for Science and Management; John Wiley & Sons, Ltd.: Chichester, UK, 2012; pp. 1–18. [Google Scholar]
- Dufour, S.; Rodríguez-González, P.M.; Laslier, M. Tracing the Scientific Trajectory of Riparian Vegetation Studies: Main Topics, Approaches and Needs in a Globally Changing World. Sci. Total Environ. 2019, 653, 1168–1185. [Google Scholar] [CrossRef]
- Carbonneau, P.E.; Dietrich, J.T. Cost-Effective Non-Metric Photogrammetry from Consumer-Grade SUAS: Implications for Direct Georeferencing of Structure from Motion Photogrammetry. Earth Surf. Process. Landf. 2017, 42, 473–486. [Google Scholar] [CrossRef] [Green Version]
- Huylenbroeck, L.; Laslier, M.; Dufour, S.; Georges, B.; Lejeune, P.; Michez, A. Using Remote Sensing to Characterize Riparian Vegetation: A Review of Available Tools and Perspectives for Managers. J. Environ. Manag. 2020, 267, 110652. [Google Scholar] [CrossRef] [PubMed]
- Farid, A.; Goodrich, D.C.; Sorooshian, S. Using Airborne Lidar to Discern Age Classes of Cottonwood Trees in a Riparian Area. West. J. Appl. For. 2006, 21, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Bertoldi, W.; Gurnell, A.M.; Drake, N.A. The Topographic Signature of Vegetation Development along a Braided River: Results of a Combined Analysis of Airborne Lidar, Color Air Photographs, and Ground Measurements. Water Resour. Res. 2011, 47, W06525. [Google Scholar] [CrossRef]
- Henshaw, A.J.; Gurnell, A.M.; Bertoldi, W.; Drake, N.A. An Assessment of the Degree to Which Landsat TM Data Can Support the Assessment of Fluvial Dynamics, as Revealed by Changes in Vegetation Extent and Channel Position, along a Large River. Geomorphology 2013, 202, 74–85. [Google Scholar] [CrossRef]
- Corenblit, D.; Steiger, J.; Charrier, G.; Darrozes, J.; Garófano-Gómez, V.; Garreau, A.; González, E.; Gurnell, A.M.; Hortobágyi, B.; Julien, F.; et al. Populus nigra L. Establishment and Fluvial Landform Construction: Biogeomorphic Dynamics within a Channelized River. Earth Surf. Process. Landf. 2016, 41, 1276–1292. [Google Scholar] [CrossRef]
- Husson, E. Images from Unmanned Aircraft Systems for Surveying Aquatic and Riparian Vegetation. PhD Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2016. [Google Scholar]
- Vautier, F.; Corenblit, D.; Hortobágyi, B.; Fafournoux, L.; Steiger, J. Monitoring and Reconstructing Past Biogeomorphic Succession within Fluvial Corridors Using Stereophotogrammetry: Stereophotogrammetry. Earth Surf. Process. Landf. 2016, 41, 1448–1463. [Google Scholar] [CrossRef]
- Bywater-Reyes, S.; Wilcox, A.C.; Diehl, R.M. Multiscale Influence of Woody Riparian Vegetation on Fluvial Topography Quantified with Ground-Based and Airborne Lidar. J. Geophys. Res. Earth Surf. 2017, 122, 1218–1235. [Google Scholar] [CrossRef]
- Lallias-Tacon, S.; Liébault, F.; Piégay, H. Use of Airborne LiDAR and Historical Aerial Photos for Characterising the History of Braided River Floodplain Morphology and Vegetation Responses. Catena 2017, 149, 742–759. [Google Scholar] [CrossRef]
- Räpple, B.; Piégay, H.; Stella, J.C.; Mercier, D. What Drives Riparian Vegetation Encroachment in Braided River Channels at Patch to Reach Scales? Insights from Annual Airborne Surveys (Drôme River, SE France, 2005–2011). Ecohydrology 2017, 10, e1886. [Google Scholar] [CrossRef]
- Hortobágyi, B. Multi-Scale Interactions between Riparian Vegetation and Hydrogeomorphic Processes (The Lower Allier River). Doctoral Dissertation, Université Clermont Auvergne, Clermont-Ferrand, France, 2018. [Google Scholar]
- Laslier, M. Suivi Des Impacts d’un Arasement de Barrage Sur La Végétation Riveraine Par Télédétection à Très Haute Résolution Spatiale et Temporelle. Doctoral Dissertation, Université Rennes 2, Rennes, France, 2018. [Google Scholar]
- Milani, G.; Kneubühler, M.; Tonolla, D.; Doering, M.; Wiesenberg, G.L.B.; Schaepman, M.E. Remotely Sensing Variation in Ecological Strategies and Plant Traits of Willows in Perialpine Floodplains. J. Geophys. Res. Biogeosciences 2019, 124, 2090–2106. [Google Scholar] [CrossRef] [Green Version]
- Kibler, C.L.; Schmidt, E.C.; Roberts, D.A.; Stella, J.C.; Kui, L.; Lambert, A.M.; Singer, M.B. A Brown Wave of Riparian Woodland Mortality Following Groundwater Declines during the 2012–2019 California Drought. Environ. Res. Lett. 2021, 16, 084030. [Google Scholar] [CrossRef]
- Dufour, S.; Muller, E.; Straatsma, M.; Corgne, S. Image Utilisation for the Study and Management of Riparian Vegetation: Overview and Applications. In Fluvial Remote Sensing for Science and Management; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 215–239. ISBN 978-1-119-94079-1. [Google Scholar]
- Michez, A.; Piégay, H.; Jonathan, L.; Claessens, H.; Lejeune, P. Mapping of Riparian Invasive Species with Supervised Classification of Unmanned Aerial System (UAS) Imagery. Int. J. Appl. Earth Obs. Geoinf. 2016, 44, 88–94. [Google Scholar] [CrossRef]
- Michez, A.; Piégay, H.; Toromanoff, F.; Brogna, D.; Bonnet, S.; Lejeune, P.; Claessens, H. LiDAR Derived Ecological Integrity Indicators for Riparian Zones: Application to the Houille River in Southern Belgium/Northern France. Ecol. Indic. 2013, 34, 627–640. [Google Scholar] [CrossRef]
- Demarchi, L.; Bizzi, S.; Piégay, H. Hierarchical Object-Based Mapping of Riverscape Units and in-Stream Mesohabitats Using LiDAR and VHR Imagery. Remote Sens. 2016, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Bizzi, S.; Piégay, H.; Demarchi, L.; Van de Bund, W.; Weissteiner, C.J.; Gob, F. LiDAR-Based Fluvial Remote Sensing to Assess 50–100-Year Human-Driven Channel Changes at a Regional Level: The Case of the Piedmont Region, Italy. Earth Surf. Process. Landf. 2019, 44, 471–489. [Google Scholar] [CrossRef]
- Roberts, D.; Roth, K.; Perroy, R. Hyperspectral Vegetation Indices. In Hyperspectral Remote Sensing of Vegetation; CRC Press: Boca Raton, FL, USA, 2011; pp. 309–327. [Google Scholar] [CrossRef]
- Gao, B. NDWI—A Normalized Difference Water Index for Remote Sensing of Vegetation Liquid Water from Space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Spectral Reflectance Changes Associated with Autumn Senescence of Aesculus Hippocastanum L. and Acer platanoides L. Leaves. Spectral Features and Relation to Chlorophyll Estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Hunt, E.R.; Rock, B.N. Detection of Changes in Leaf Water Content Using Near- and Middle-Infrared Reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar] [CrossRef]
- Penuelas, J.; Frederic, B.; Filella, I. Semi-Empirical Indices to Assess Carotenoids/Chlorophyll-a Ratio from Leaf Spectral Reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Sims, D.A.; Gamon, J.A. Relationships between Leaf Pigment Content and Spectral Reflectance across a Wide Range of Species, Leaf Structures and Developmental Stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Pascucci, S.; Pignatti, S.; Casa, R.; Darvishzadeh, R.; Huang, W. Special Issue “Hyperspectral Remote Sensing of Agriculture and Vegetation”. Remote Sens. 2020, 12, 3665. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Tree Species Classification in the Southern Alps Based on the Fusion of Very High Geometrical Resolution Multispectral/Hyperspectral Images and LiDAR Data. Remote Sens. Environ. 2012, 123, 258–270. [Google Scholar] [CrossRef]
- He, K.S.; Rocchini, D.; Neteler, M.; Nagendra, H. Benefits of Hyperspectral Remote Sensing for Tracking Plant Invasions. Divers. Distrib. 2011, 17, 381–392. [Google Scholar] [CrossRef]
- Shoot, C.; Andersen, H.-E.; Moskal, L.M.; Babcock, C.; Cook, B.D.; Morton, D.C. Classifying Forest Type in the National Forest Inventory Context with Airborne Hyperspectral and Lidar Data. Remote Sens. 2021, 13, 1863. [Google Scholar] [CrossRef]
- Demarchi, L.; Kania, A.; Ciężkowski, W.; Piórkowski, H.; Oświecimska-Piasko, Z.; Chormański, J. Recursive Feature Elimination and Random Forest Classification of Natura 2000 Grasslands in Lowland River Valleys of Poland Based on Airborne Hyperspectral and LiDAR Data Fusion. Remote Sens. 2020, 12, 1842. [Google Scholar] [CrossRef]
- da Silva, A.R.; Demarchi, L.; Sikorska, D.; Sikorski, P.; Archiciński, P.; Jóźwiak, J.; Chormański, J. Multi-Source Remote Sensing Recognition of Plant Communities at the Reach Scale of the Vistula River, Poland. Ecol. Indic. 2022, 142, 109160. [Google Scholar] [CrossRef]
- Dutta, D.; Wang, K.; Lee, E.; Goodwell, A.; Woo, D.K.; Wagner, D.; Kumar, P. Characterizing Vegetation Canopy Structure Using Airborne Remote Sensing Data. IEEE Trans. Geosci. Remote Sens. 2017, 55, 1160–1178. [Google Scholar] [CrossRef]
- Richter, R.; Reu, B.; Wirth, C.; Doktor, D.; Vohland, M. The Use of Airborne Hyperspectral Data for Tree Species Classification in a Species-Rich Central European Forest Area. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 464–474. [Google Scholar] [CrossRef]
- Shendryk, I.; Broich, M.; Tulbure, M.G.; McGrath, A.; Keith, D.; Alexandrov, S.V. Mapping Individual Tree Health Using Full-Waveform Airborne Laser Scans and Imaging Spectroscopy: A Case Study for a Floodplain Eucalypt Forest. Remote Sens. Environ. 2016, 187, 202–217. [Google Scholar] [CrossRef]
- Rollet, A.J.; Piégay, H.; Dufour, S.; Bornette, G.; Persat, H. Assessment of Consequences of Sediment Deficit on a Gravel River Bed Downstream of Dams in Restoration Perspectives: Application of a Multicriteria, Hierarchical and Spatially Explicit Diagnosis. River Res. Appl. 2014, 30, 939–953. [Google Scholar] [CrossRef]
- Piégay, H.; Bornette, G.; Citterio, A.; Hérouin, E.; Moulin, B.; Statiotis, C. Channel Instability as a Control on Silting Dynamics and Vegetation Patterns within Perifluvial Aquatic Zones. Hydrol. Process. 2000, 14, 3011–3029. [Google Scholar] [CrossRef]
- Dufour, S. Contrôles Naturels Anthropiques de La Structure et de La Dynamique Des Forêts Riveraines. PhD Thesis, Université Jean Moulin Lyon III, Lyon, France, 2005. [Google Scholar]
- Dumas, S. Les Habitats Forestiers de La Basse Vallée de l’Ain: Étude et Analyse; Office National des Forêts: Saint Denis, France, 2004; p. 38. [Google Scholar]
- Dumas, S.; Perrin, V. Le Suivi de La Forêt Alluviale de La Basse Vallée de l’Ain: Inventaire de Niveau II de 2006; Office National des Forêts: Saint Denis, France, 2006; p. 66. [Google Scholar]
- Dumas, S. Inventaire Des Boisements Forestiers de La Basse Vallée de l’Ain; Office National des Forêts: Saint Denis, France, 2017; p. 39. [Google Scholar]
- Lejot, J.; Piégay, H.; Hunter, P.D.; Moulin, B.; Gagnage, M. Utilisation de la télédétection pour la caractérisation des corridors fluviaux: Exemples d’applications et enjeux actuels. Géomorphol. Relief Process. Environ. 2011, 17, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Lague, D.; Feldmann, B. Topo-Bathymetric Airborne LiDAR for Fluvial-Geomorphology Analysis. In Remote Sensing of Geomorphology; Paolo Tarolli, S.M.M., Ed.; Developments in Earth Surface Processes; Elsevier: Amsterdam, The Netherlands, 2020; Volume 23, pp. 25–54. [Google Scholar]
- Roussel, J.-R.; Auty, D.; Coops, N.; Tompalski, P.; Goodbody, T.R.H.; Meador, A.S.; Bourdon, J.-F.; De Boissieu, F.; Achim, A. LidR: An R Package for Analysis of Airborne Laser Scanning (ALS) Data. Remote Sens. Environ. 2020, 251, 112061. [Google Scholar] [CrossRef]
- Roux, C.; Alber, A.; Bertrand, M.; Vaudor, L.; Piégay, H. “FluvialCorridor”: A New ArcGIS Toolbox Package for Multiscale Riverscape Exploration. Geomorphology 2015, 242, 29–37. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Böhner, J. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red Edge Spectral Measurements from Sugar Maple Leaves. Int. J. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Hardisky, M.; Klemas, V. Smart, and The Influence of Soil Salinity, Growth Form, and Leaf Moisture on the Spectral Radiance of Spartina Alterniflora Canopies. Photogramm. Eng. Remote Sens. 1983, 48, 77–84. [Google Scholar]
- Wang, L.; Qu, J.J. NMDI: A Normalized Multi-Band Drought Index for Monitoring Soil and Vegetation Moisture with Satellite Remote Sensing. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef]
- Dufour, S.; Rinaldi, M.; Piégay, H.; Michalon, A. How Do River Dynamics and Human Influences Affect the Landscape Pattern of Fluvial Corridors? Lessons from the Magra River, Central–Northern Italy. Landsc. Urban Plan. 2015, 134, 107–118. [Google Scholar] [CrossRef]
- Hamada, Y.; Stow, D.A.; Coulter, L.L.; Jafolla, J.C.; Hendricks, L.W. Detecting Tamarisk Species (Tamarix spp.) in Riparian Habitats of Southern California Using High Spatial Resolution Hyperspectral Imagery. Remote Sens. Environ. 2007, 109, 237–248. [Google Scholar] [CrossRef]
- Belcore, E.; Latella, M. Riparian Ecosystems Mapping at Fine Scale: A Density Approach Based on Multi-Temporal UAV Photogrammetric Point Clouds. Remote Sens. Ecol. Conserv. 2022, 8, 644–655. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Data Type | Multi-Date Acquisition | Species Identification | Structural Information | Temporal Dynamics | Topography and Hydrological Connectivity |
|---|---|---|---|---|---|---|
| [28] | LiDAR | X | ||||
| [29] | RGB imagery + LiDAR | X | X | |||
| [30] | Landsat imagery | X | X | |||
| [31] | RGB imagery + photogrammetry | X | X | X | X | |
| [32] | RGB imagery | X | ||||
| [33] | RGB imagery + photogrammetry | X | X | X | X | |
| [34] | LiDAR | X | X | |||
| [35] | RGB imagery + LiDAR | X | X | X | X | |
| [36] | RGB imagery + LiDAR | X | X | X | X | |
| [37] | RGB imagery + LiDAR + photogrammetry | X | X | X | ||
| [38] | RGB imagery + LiDAR | X | X | X | X | X |
| [39] | Hyperspectral imagery | X | X | |||
| [40] | Hyperspectral imagery + Landsat imagery | X | X | X |
| Type of Data | Years of Acquisition | Spatial Resolution | Spectral Information |
|---|---|---|---|
| Aerial Photographs | 1945, 1954, 1963, 1971, 1980, 1991, 1996, 2000, 2005, 2009, 2012 | 0.5 × 0.5–1 × 1 m | Color RGB imagery since 2000, black and white before |
| LiDAR | 2008 | 1.8 pts/m2 | NIR Laser, but only topographic points were available |
| LiDAR | 2015 | 18.6 pts/m2 per laser | NIR laser + green laser |
| Hyperspectral | 2015 | 1 × 1 m | 361 spectral bands (380–2500 nm) |
| Characteristic | 2015 Vegetation Plots (EVS Lab) | 2017 Vegetation Plots (ONF Survey) |
|---|---|---|
| Species composition | X | X |
| Tree diameter | 10 m radius | |
| if diameter > 30 cm | ||
| 5 m radius | ||
| if diameter > 7.5 cm | ||
| Tree height (in 5-m classes) | 10 m radius | |
| if diameter > 30 cm | ||
| 5 m radius | ||
| if diameter > 7.5 cm | ||
| Basal area | X | |
| Understory cover | 5 m radius | |
| Grass cover | 5 m radius | |
| Dead trees | 10 m radius | 5 m radius |
| if diameter > 30 cm | ||
| 5 m radius | ||
| if diameter > 7.5 cm | ||
| Soil depth | X | X |
| Soil humidity | X | |
| Organic matter | X | |
| Age | X |
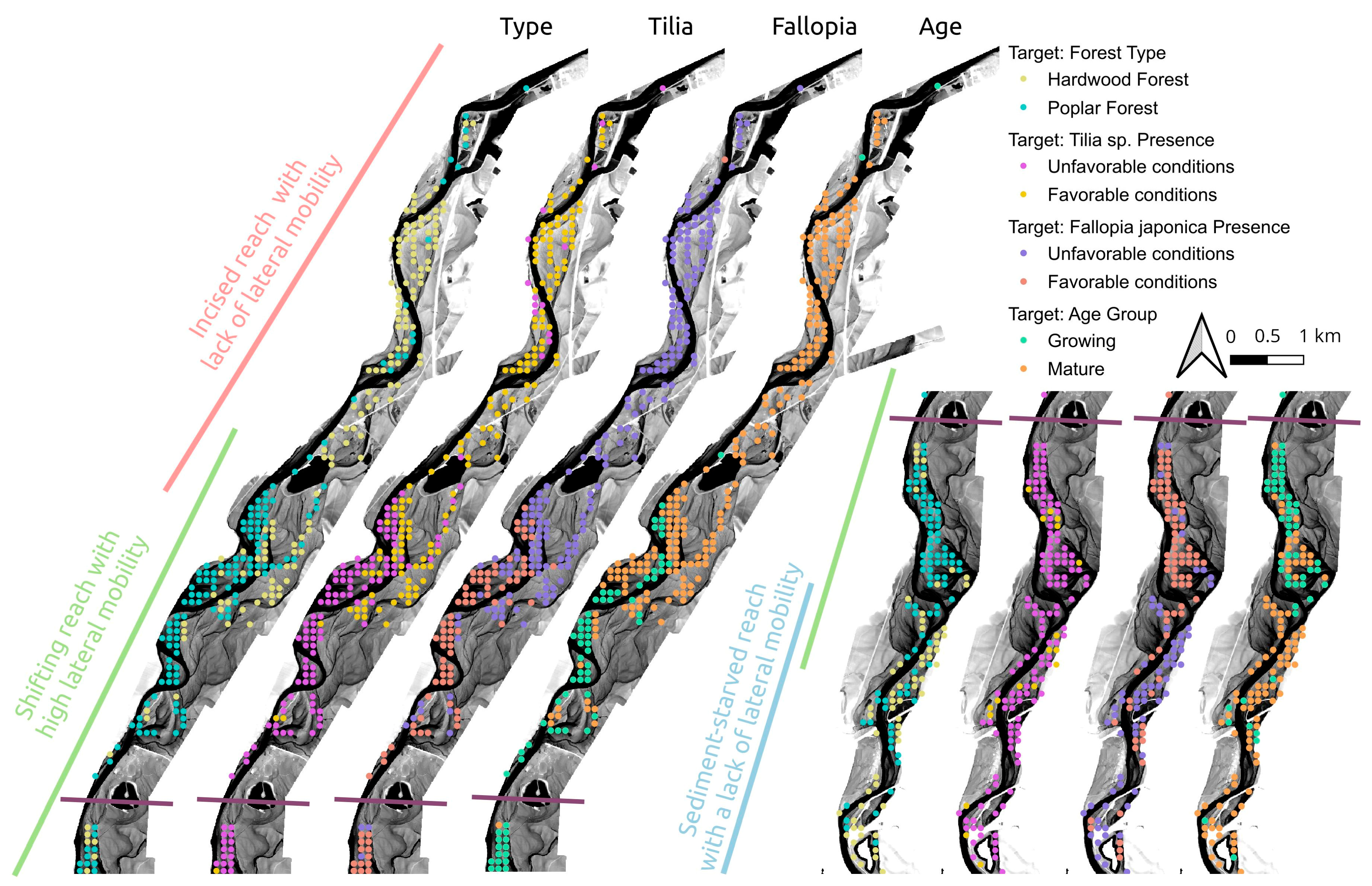
| Classification Target | Classes | Site Conditions |
|---|---|---|
| Age group | Growing and mature | Identifies the presence of lateral mobility and forest rejuvenation |
| Forest type | Poplar forest and hardwood forest | The poplar forest should be located in growing forest patches and in mature forest patches that are well-connected to the river |
| Fallopia japonica | Presence and absence | Requires a wet environment and well-connected forest patches |
| Tilia cordata | Presence and absence | Colonizes and grows on the driest forest patches |
| Site and Plot Number | Reach | Proportion of Tilia sp. | Proportion of Shrub Species | Presence of Fraxinus excelsior |
|---|---|---|---|---|
| Mollon 1 | Shifting | 2% | 8% | Present |
| Mollon 2 | Shifting | 0% | 0% | Present |
| Martinaz 1 | Shifting | 10% | 10% | Present |
| Martinaz 2 | Shifting | 6% | 9% | Present |
| Cormoz 1 | Incised | 56% | 0% | Absent |
| Cormoz 2 | Incised | 0% | 85% | Absent |
| Bellegarde 1 | Incised | 0% | 0% | Present |
| Bellegarde 2 | Incised | 0% | 0% | Present |
| Vorgey 1 | Incised | 0% | 41% | Present |
| Vorgey 2 | Incised | 78% | 0% | Present |
| Spectral Index | Reference | Target | R2 vs. Mean Height (<50 y.o.) | R2 vs. Mean Height (>50 y.o.) | R2 vs. Mean Height (All Plots in the Shifting Reach) |
|---|---|---|---|---|---|
| ReNDVI | [48] | Greenness | 0.09 | 0.47 | 0.30 |
| VREI1 | [72] | Greenness | 0.11 | 0.52 | 0.33 |
| NDII | [73] | Canopy water content | 0.21 | 0.49 | 0.33 |
| NDMI | [74] | Canopy water content | 0.20 | 0.47 | 0.31 |
| MSI | [49] | Canopy water content | 0.21 | 0.49 | 0.33 |
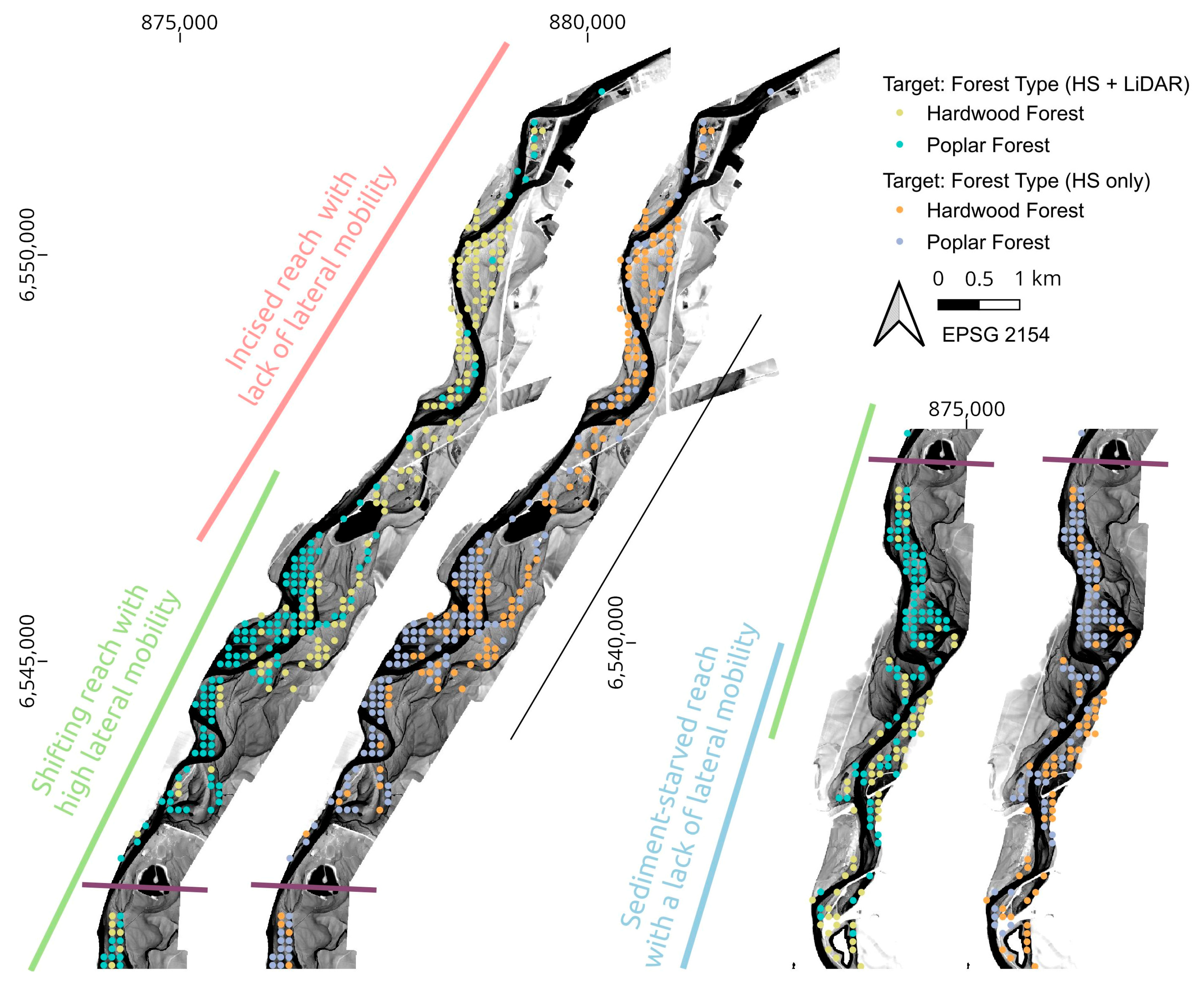
| Classification Target | Class | Class Error LiDAR + HS | Class Error LiDAR Only | Class Error HS Only |
|---|---|---|---|---|
| Age group | Growing (<50 y.o.) | 18% | 24% | 20% |
| Mature (>50 y.o.) | 16% | 16% | 28% | |
| Forest type | Poplar forest | 14% | 18% | 20% |
| Hardwood forest | 12% | 16% | 18% | |
| Presence of Fallopia japonica | Present | 20% | 20% | 26% |
| Absent | 16% | 14% | 28% | |
| Presence of Tilia cordata | Present | 20% | 36% | 26% |
| Absent | 18% | 28% | 24% |
| Classification Target | Class | Predicted A. | Predicted B. | Class Error | Mean Error |
|---|---|---|---|---|---|
| Age group | (A.) Growing (<50 y.o.) | 84 | 19 | 18.5% | 13.5% |
| (B.) Mature (>50 y.o.) | 28 | 281 | 9% | ||
| Forest type | (A.) Poplar forest | 120 | 17 | 12% | 11% |
| (B.) Hardwood forest | 8 | 72 | 10% | ||
| Presence of Fallopia japonica | (A.) Present | 76 | 6 | 7.5% | 12% |
| (B.) Absent | 53 | 277 | 16% | ||
| Presence of Tilia cordata | (A.) Present | 96 | 9 | 9.5% | 17% |
| (B.) Absent | 75 | 232 | 24.5% |
| Forest Type (ONF) | Age Group | Forest Type | Fallopia japonica | Tilia cordata | ||||
|---|---|---|---|---|---|---|---|---|
| Growing | Mature | Poplar | Hardwood | Present | Absent | Present | Absent | |
| Early pioneer forest | 17 | 0 | 16 | 1 | 15 | 2 | 0 | 18 |
| Rapid growth series | 32 | 35 | 62 | 5 | 47 | 20 | 6 | 61 |
| Slow growth series | 22 | 22 | 29 | 15 | 21 | 23 | 15 | 29 |
| Mature poplar forest with an understory | 2 | 122 | 56 | 68 | 17 | 107 | 81 | 43 |
| Post-pioneer hardwood forest | 6 | 88 | 13 | 81 | 15 | 79 | 59 | 35 |
| Others | 29 | 37 | 36 | 30 | 36 | 30 | 10 | 56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godfroy, J.; Lejot, J.; Demarchi, L.; Bizzi, S.; Michel, K.; Piégay, H. Combining Hyperspectral, LiDAR, and Forestry Data to Characterize Riparian Forests along Age and Hydrological Gradients. Remote Sens. 2023, 15, 17. https://doi.org/10.3390/rs15010017
Godfroy J, Lejot J, Demarchi L, Bizzi S, Michel K, Piégay H. Combining Hyperspectral, LiDAR, and Forestry Data to Characterize Riparian Forests along Age and Hydrological Gradients. Remote Sensing. 2023; 15(1):17. https://doi.org/10.3390/rs15010017
Chicago/Turabian StyleGodfroy, Julien, Jérôme Lejot, Luca Demarchi, Simone Bizzi, Kristell Michel, and Hervé Piégay. 2023. "Combining Hyperspectral, LiDAR, and Forestry Data to Characterize Riparian Forests along Age and Hydrological Gradients" Remote Sensing 15, no. 1: 17. https://doi.org/10.3390/rs15010017