
Spatial and Seasonal Variations of the Island Mass Effect at the Sub-Antarctic Prince Edward Islands Archipelago


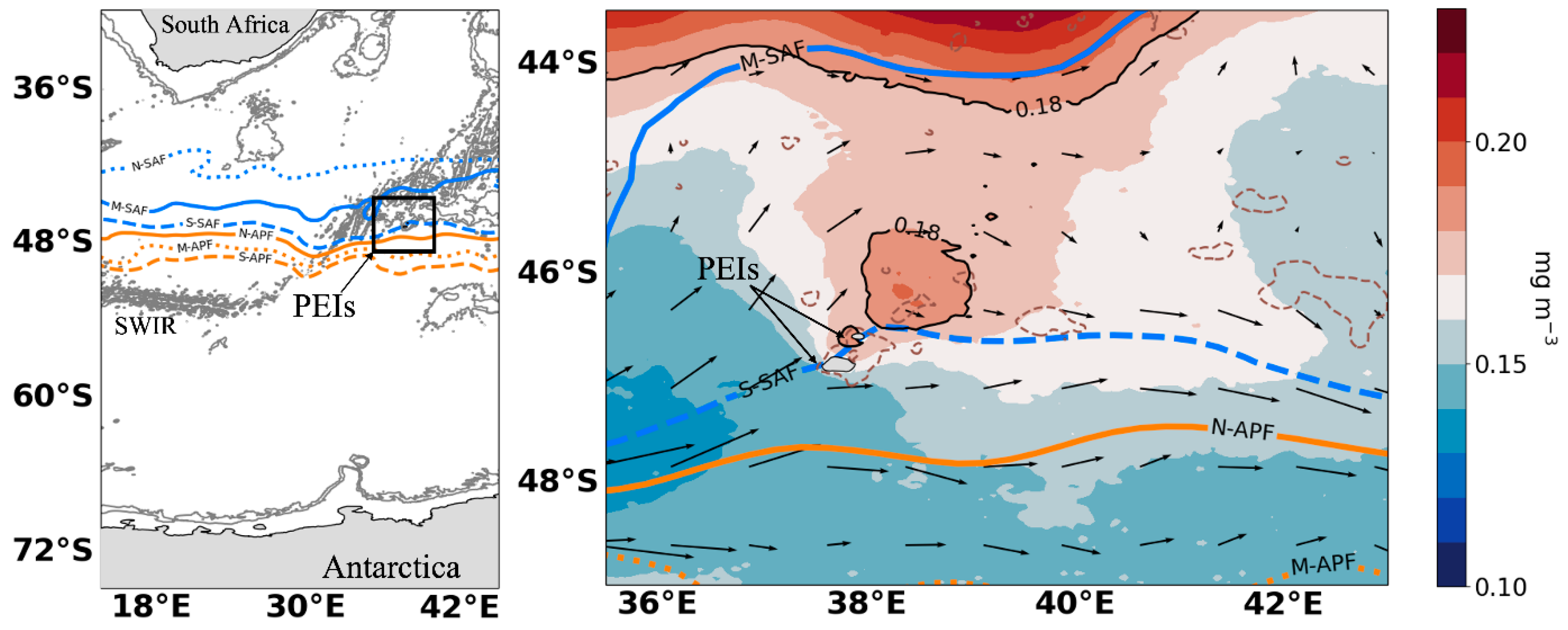{kind=link}
{kind=link}
{kind=link}
{kind=link}
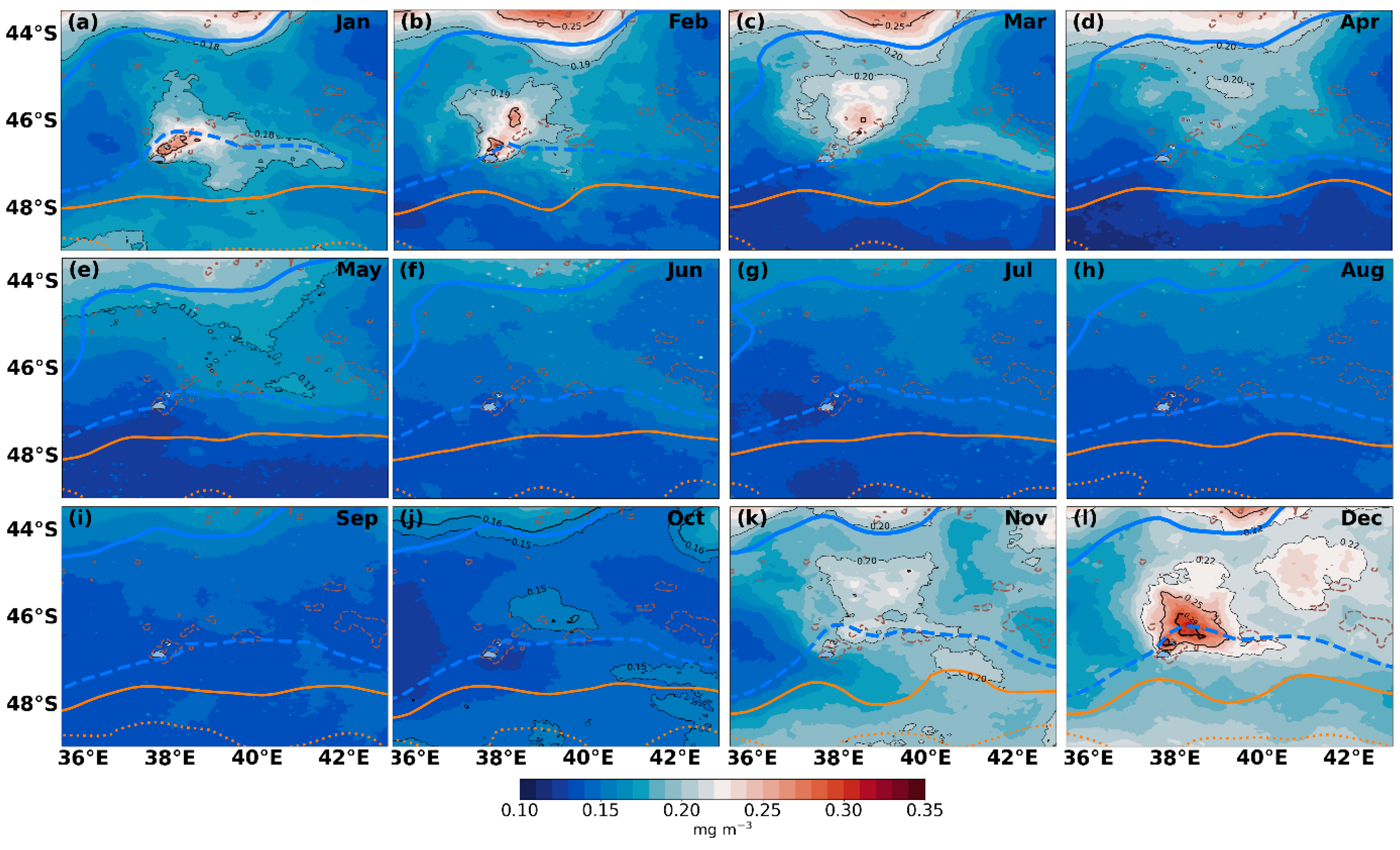
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Data and Methods
3. Results
3.1. Spatial and Seasonal Chlorophyll a Patterns
3.2. Factors Influencing Chlorophyll a Variability
3.2.1. Light, Temperature, and Nutrients
3.2.2. Mixed Layer Depths, Winds, and Currents
4. Discussion
4.1. Chlorophyll a Variability
4.2. Influence of Environmental Forcing on Chlorophyll a Variations
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deppeler, S.L.; Davidson, A.T. Southern Ocean phytoplankton in a changing climate. Front. Mar. Sci. 2017, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Pinkerton, M.H.; Boyd, P.W.; Deppeler, S.; Hayward, A.; Höfer, J.; Moreau, S. Evidence for the impact of climate change on primary producers in the Southern Ocean. Front. Ecol. Evol. 2021, 9, 592027. [Google Scholar] [CrossRef]
- Rogers, A.D.; Frinault, B.A.V.; Barnes, D.K.A.; Bindoff, N.L.; Downie, R.; Ducklow, H.W.; Friedlaender, A.S.; Hart, T.; Hill, S.L.; Hofmann, E.E.; et al. Antarctic Futures: An assessment of climate-driven changes in ecosystem structure, function, and service provisioning in the Southern Ocean. Ann. Rev. Mar. Sci. 2020, 12, 87–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento, J.L.; Gruber, N.; Brzezinski, M.A.; Dunne, J.P. High-latitude controls of thermocline nutrients and low latitude biological productivity. Nature 2004, 427, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, K.R.; van Dijken, G.L.; Bushinsky, S. Primary production in the Southern Ocean, 1997–2006. J. Geophys. Res. 2008, 113, C08004. [Google Scholar] [CrossRef]
- Bristow, L.A.; Mohr, W.; Ahmerkamp, S.; Kuypers, M.M.M. Nutrients that limit growth in the ocean. Curr. Biol. 2017, 27, R431–R510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venables, H.; Moore, C.M. Phytoplankton and light limitation in the Southern Ocean: Learning from high-nutrient, high-chlorophyll areas. J. Geophys. Res. 2010, 115, C02015. [Google Scholar] [CrossRef]
- Browning, T.J.; Achterberg, E.P.; Engel, A.; Mawji, E. Manganese co-limitation of phytoplankton growth and major nutrient drawdown in the Southern Ocean. Nat. Commun. 2021, 12, 884. [Google Scholar] [CrossRef]
- Graham, R.M.; De Boer, A.M.; van Sebille, E.; Kohfeld, K.E.; Schlosser, C. Inferring source regions and supply mechanisms of iron in the Southern Ocean from satellite chlorophyll data. Deep-Sea Res. I 2005, 104, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.K.; Abbott, M.R. Surface chlorophyll concentrations in relation to the Antarctic Polar Front: Seasonal and spatial patterns from satellite observations. J. Mar. Syst. 2002, 37, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, S.; Rintoul, S.R. On the relationship between fronts and the Antarctic Circumpolar Current and surface chlorophyll concentrations in the Southern Ocean. J. Geophys. Res. 2007, 112, C07030. [Google Scholar] [CrossRef]
- Boden, B.P. Observations of the island mass effect in the Prince Edward archipelago. Polar Biol. 1988, 9, 61–68. [Google Scholar] [CrossRef]
- Gove, J.M.; McManus, M.A.; Neuheimer, A.B.; Polovina, J.J.; Drazen, J.C.; Smith, C.R.; Merrifield, M.A.; Friedlander, A.M.; Ehses, J.S.; Young, C.W.; et al. Near-island biological hotspots in barren ocean basins. Nat. Commun. 2016, 7, 10581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gove, J.M.; Williams, G.J.; McManus, M.A.; Heron, S.F.; Sandin, S.A.; Vetter, O.J.; Foley, D.G. Quantifying climatological ranges and anomalies for Pacific coral reef ecosystems. PLoS ONE 2013, 8, e61974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.M.; Bakker, D.C.E.; Venables, H.J.; Watson, A.J. Dynamic seasonal cycling of inorganic carbon downstream of South Georgia, Southern Ocean. Deep-Sea Res. II 2012, 59–60, 25–35. [Google Scholar] [CrossRef]
- Quilty, P.G. Origin and evolution of the sub-Antarctic islands: The foundation. Pap. Proc. R. Soc. Tasmania 2007, 141, 35–58. [Google Scholar] [CrossRef] [Green Version]
- Lutjeharms, J.R.E.; Ansorge, I.J. Oceanographic setting of the Prince Edward Islands. In The Prince Edward Islands: Land-Sea Interactions in a Changing Ecosystem; Chown, S.L., Froneman, P.W., Eds.; Sun Press: Stellenbosch, South Africa, 2008; pp. 17–38. [Google Scholar]
- Whitehead, O.; von der Meden, C.; Skowno, A.L.; Sink, K.J.; van der Merwe, S.; Adams, R.; Holness, S. (Eds.) South African National Biodiversity Assessment 2018 Technical Report Volume 6: Sub-Antarctic Territory; South African National Biodiversity Institute: Pretoria, South Africa, 2019; Available online: http://hdl.handle.net/20.500.12143/6375 (accessed on 20 September 2021).
- Kirkman, S.P.; Yemane, D.G.; Lamont, T.; Meÿer, M.A.; Pistorius, P.A. Foraging behaviour of Subantarctic Fur Seals supports efficiency of a marine reserve’s design. PLoS ONE 2016, 11, e0152370. [Google Scholar] [CrossRef] [Green Version]
- Pakhomov, E.A.; Froneman, P.W. The Prince Edward Islands pelagic ecosystem: A review of achievements 1976–1990. J. Mar. Syst. 1999, 18, 297–310. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Froneman, P.W. Macroplankton/micronekton dynamics in the vicinity of the Prince Edward Islands (Southern Ocean). Mar. Biol. 1999, 134, 501–515. [Google Scholar] [CrossRef]
- Perissinotto, R.; McQuaid, C.D. Land-based predator impact on vertically migrating zooplankton and micronekton advected to a Southern Ocean archipelago. Mar. Ecol. Prog. Ser. 1992, 80, 15–27. [Google Scholar] [CrossRef]
- Ryan, P.G.; Bester, M.N. Pelagic predators. In The Prince Edward Islands: Land-Sea Interactions in a Changing Ecosystem; Chown, S.L., Froneman, P.W., Eds.; Sun Press: Stellenbosch, South Africa, 2008; pp. 121–164. [Google Scholar]
- von der Meden, C.E.O.; Atkinson, L.J.; Branch, G.M.; Asdar, S.; Ansorge, I.J.; van den Berg, M. Long-term change in epibenthic assemblages at the Prince Edward Islands: A comparison between 1988 and 2013. Polar Biol. 2017, 40, 2171–2185. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Chown, S.L. The Prince Edward Islands: Southern Ocean Oasis. Ocean Yearb. 2003, 17, 348–379. [Google Scholar] [CrossRef]
- McQuaid, C.D.; Froneman, P.W. Biology in the oceanographic environment. In The Prince Edward Islands: Land-Sea Interactions in a Changing Ecosystem; Chown, S.L., Froneman, P.W., Eds.; Sun Press: Stellenbosch, South Africa, 2008; pp. 97–120. [Google Scholar]
- Rae, C.M.D. Physical and chemical marine environment of the Prince Edward Islands (Southern Ocean) during April/May 1987. S. Afr. J. Mar. Sci. 1989, 8, 301–311. [Google Scholar] [CrossRef]
- Lamont, T.; van den Berg, M.A.; Tutt, G.C.O.; Ansorge, I.J. Impact of deep-ocean eddies and fronts on the shelf seas of a sub-Antarctic Archipelago: The Prince Edward Islands. Cont. Shelf Res. 2019, 177, 1–14. [Google Scholar] [CrossRef]
- Perissinotto, R.; Duncombe Rae, C.M. Occurrence of anti-cyclonic eddies on the Prince Edward Plateau (Southern Ocean): Effects on phytoplankton biomass and production. Deep-Sea Res. I 1990, 37, 777–793. [Google Scholar] [CrossRef]
- Perissinotto, R.; Duncombe Rae, C.M.; Boden, B.P.; Allanson, B.R. Vertical stability as a controlling factor of the marine phytoplankton production at the Prince Edward Archipelago (Southern Ocean). Mar. Ecol. Prog. Ser. 1990, 60, 205–209. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Froneman, P.W.; Ansorge, I.J.; Lutjeharms, J.R.E. Temporal variability in the physico-biological environment of the Prince Edward Islands (Southern Ocean). J. Mar. Syst. 2000, 26, 75–95. [Google Scholar] [CrossRef]
- Lamont, T.; Tutt, G.C.O.; Barlow, R.G. Phytoplankton biomass and photophysiology at the sub-Antarctic Prince Edward Islands ecosystem in the Southern Ocean. J. Mar. Syst. 2022, 226, 103669. [Google Scholar] [CrossRef]
- Bernard, A.T.F.; Ansorge, I.J.; Froneman, P.W.; Lutjeharms, J.R.E.; Bernard, K.S.; Swart, N.C. Entrainment of Antarctic euphausiids across the Antarctic Polar Front by a cold eddy. Deep-Sea Res. I 2007, 54, 1841–1851. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Ansorge, I.J.; Froneman, P.W. Variability in the inter-island environment of the Prince Edward Islands (Southern Ocean). Polar Biol. 2000, 23, 593–603. [Google Scholar] [CrossRef]
- Lamont, T.; van den Berg, M.A. Mesoscale eddies influencing the sub-Antarctic Prince Edward Islands Archipelago: Temporal variability and impact. Cont. Shelf Res. 2021, 212, 104309. [Google Scholar] [CrossRef]
- Perissinotto, R.; Lutjeharms, J.R.E.; van Ballegooyen, R.C. Biological-physical interactions and pelagic productivity at the Prince Edward Islands, Southern Ocean. J. Mar. Syst. 2000, 24, 327–341. [Google Scholar] [CrossRef]
- Ansorge, I.J.; Lutjeharms, J.R.E. Eddies originating at the South-West Indian Ridge. J. Mar. Syst. 2003, 39, 1–18. [Google Scholar] [CrossRef]
- Ansorge, I.J.; Lutjeharms, J.R.E. Direct observations of eddy turbulence at a ridge in the Southern Ocean. Geophys. Res. Lett. 2005, 32, L14603. [Google Scholar] [CrossRef]
- Durgadoo, J.V.; Ansorge, I.J.; de Cuevas, B.A.; Lutjeharms, J.R.E.; Coward, A.C. Decay of eddies at the South-West Indian Ridge. S. Afr. J. Sci. 2011, 107, 673. [Google Scholar] [CrossRef]
- Durgadoo, J.V.; Ansorge, I.J.; Lutjeharms, J.R.E. Oceanographic observations of eddies impacting the Prince Edward Islands, South Africa. Antarct. Sci. 2010, 22, 211–219. [Google Scholar] [CrossRef]
- Lamont, T.; van den Berg, M.A. Mesoscale eddies influencing the sub-Antarctic Prince Edward Islands Archipelago: Origin, Pathways, and Characteristics. Cont. Shelf Res. 2020, 210, 104257. [Google Scholar] [CrossRef]
- Ansorge, I.J.; Froneman, P.W.; Pakhomov, E.A.; Lutjeharms, J.R.E.; Perissinotto, R.; van Ballegooyen, R.C. Physical-biological coupling in the waters surrounding the Prince Edward Islands (Southern Ocean). Polar Biol. 1999, 21, 135–145. [Google Scholar] [CrossRef]
- Ansorge, I.J.; Durgadoo, J.V.; Pakhomov, E.A. Dynamics of physical and biological systems of the Prince Edward Islands in a changing climate. Pap. Proc. R. Soc. Tasmania 2009, 143, 15–18. [Google Scholar] [CrossRef]
- Ansorge, I.J.; Froneman, P.W.; Durgadoo, J.V. The marine ecosystem of the sub-Antarctic, Prince Edward Islands. In Marine Ecosystems; Cruzado, A., Ed.; InTech Press: Rijeka, Croatia, 2011; pp. 61–76. [Google Scholar] [CrossRef] [Green Version]
- Ansorge, I.J.; Lutjeharms, J.R.E. The hydrography and dynamics of the ocean environment of the Prince Edward Islands (Southern Ocean). J. Mar. Syst. 2002, 37, 107–127. [Google Scholar] [CrossRef]
- Stirnimann, L.; Bornman, T.G.; Verheye, H.M.; Bachèlery, M.-L.; van der Poel, J.; Fawcett, S.E. Plankton community composition and productivity near the Subantarctic Prince Edward Islands archipelago in autumn. Limnol. Oceanogr. 2021, 66, 4140–4158. [Google Scholar] [CrossRef]
- Venkatachalam, S.; Matcher, G.F.; Lamont, T.; van den Berg, M.; Ansorge, I.J.; Dorrington, R.A. Influence of oceanographic variability on near-shore microbial communities of the sub-Antarctic Prince Edward Islands. Limnol. Oceanogr. 2019, 64, 258–271. [Google Scholar] [CrossRef]
- Chown, S.L.; Froneman, P.W. (Eds.) The Prince Edward Islands: Land-Sea Interactions in a Changing Ecosystem; Sun Press: Stellenbosch, South Africa, 2008. [Google Scholar] [CrossRef] [Green Version]
- Toolsee, T.; Lamont, T.; Rouault, M.; Ansorge, I. Characterising the seasonal cycle of wind forcing, surface circulation and temperature around the sub-Antarctic Prince Edward Islands. Afr. J. Mar. Sci. 2021, 43, 61–76. [Google Scholar] [CrossRef]
- Ardyna, M.; Clautre, H.; Sallée, J.-B.; d’Ovidio, F.; Gentili, B.; van Dijken, G.; D’Ortenzio, F.; Arrigo, K.R. Delineating environmental control of phytoplankton biomass and phenology in the Southern Ocean. Geophys. Res. Lett. 2017, 44, 5016–5024. [Google Scholar] [CrossRef]
- Lamont, T.; Brewin, R.J.W.; Barlow, R.G. Seasonal variation in remotely-sensed phytoplankton size structure around southern Africa. Remote Sens. Environ. 2018, 204, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Mongin, M.; Molina, E.; Trull, T.W. Seasonality and scale of the Kergeulen plateau phytoplankton bloom: A remote sensing and modelling analysis of the influence of natural iron fertilization in the Southern Ocean. Deep-Sea Res. II 2008, 55, 880–892. [Google Scholar] [CrossRef]
- Thomalla, S.J.; Fauchereau, N.; Swart, S.; Monteiro, P.M.S. Regional scale characteristics of the seasonal cycle of chlorophyll in the Southern Ocean. Biogeosciences 2011, 8, 2849–2866. [Google Scholar] [CrossRef] [Green Version]
- Del Castillo, C.E.; Signorini, S.R.; Karaköylü, E.M.; Rivero-Calle, S. Is the Southern Ocean getting greener? Geophys. Res. Lett. 2019, 46, 6034–6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahru, M.; Mitchell, B.G. Blending of ocean colour algorithms applied to the Southern Ocean. Remote Sens. Lett. 2010, 1, 119–124. [Google Scholar] [CrossRef]
- Garnesson, P.; Mangin, A.; Fanton d’Andon, O.; Demaria, J.; Bretagnon, M. The CMEMS GlobColour chlorophyll a product based on satellite observation: Multi-sensor merging and flagging strategies. Ocean Sci. 2019, 15, 819–830. [Google Scholar] [CrossRef] [Green Version]
- Sathyendranath, S.; Brewin, R.J.W.; Brockmann, C.; Brotas, V.; Calton, B.; Chuprin, A.; Cipollini, P.; Couto, A.B.; Dingle, J.; Doerffer, R.; et al. An Ocean-Colour time series for use in climate studies: The experience of the Ocean-Colour Climate Change Initiative (OC-CCI). Sensors 2019, 19, 4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiozaki, T.; Kodama, T.; Furuya, K. Large-scale impact of the island mass effect through nitrogen fixation in the western South Pacific Ocean. Geophys. Res. Lett. 2014, 41, 2907–2913. [Google Scholar] [CrossRef]
- Jena, B. Satellite remote sensing of the island mass effect on the Sub-Antarctic Kerguelen Plateau, Southern Ocean. Front. Earth Sci. 2016, 10, 479–486. [Google Scholar] [CrossRef]
- Martinez, E.; Maamaatuaiahutapu, K. Island mass effect in the Marquesas Islands: Time variation. Geophys. Res. Lett. 2004, 31, L18307. [Google Scholar] [CrossRef] [Green Version]
- NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Sea-Viewing Wide Field-of-View Sensor (SeaWiFS) Photosynthetically Available Radiation Data; 2018 Reprocessing; NASA OB.DAAC: Greenbelt, MD, USA, 2018. Available online: https://oceancolor.gsfc.nasa.gov/data/10.5067/ORBVIEW-2/SEAWIFS/L3M/PAR/2018/ (accessed on 25 August 2021).
- NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Moderate-Resolution Imaging Spectroradiometer (MODIS) Aqua Photosynthetically Available Radiation Data; 2018 Reprocessing; NASA OB.DAAC: Greenbelt, MD, USA, 2018. [Google Scholar] [CrossRef]
- Garcia, H.E.; Weathers, K.W.; Paver, C.R.; Smolyar, I.; Boyer, T.P.; Locarnini, R.A.; Zweng, M.M.; Mishonov, A.V.; Baranova, O.K.; Seidov, D.; et al. World Ocean Atlas 2018. Vol. 4: Dissolved Inorganic Nutrients (Phosphate, Nitrate and Nitrate + Nitrite, Silicate); Mishonov, A., Ed.; NOAA Atlas NESDIS: Silver Spring, MD, USA, 2019; Volume 84, 35p. [Google Scholar]
- Taburet, G.; Sanchez-Roman, A.; Ballarotta, M.; Pujol, M.-I.; Legeais, J.-F.; Fournier, F.; Faugere, Y.; Dibarbourne, G. DUACS DT2018: 25 years of reprocessed sea level altimetry records. Ocean Sci. 2019, 15, 1207–1224. [Google Scholar] [CrossRef] [Green Version]
- Rio, M.-H.; Mulet, S.; Picot, N. Beyond GOCE for the ocean circulation estimate: Synergetic use of altimetry, gravimetry, and in situ data provides new insight into geostrophic and Ekman currents. Geophys. Res. Lett. 2014, 41, 8919–8925. [Google Scholar] [CrossRef]
- Sokolov, S.; Rintoul, S.R. Circumpolar structure and distribution of the Antarctic Circumpolar Current fronts: 1. Mean circumpolar paths. J. Geophys. Res. 2009, 114, C11018. [Google Scholar] [CrossRef]
- Sokolov, S.; Rintoul, S.R. Circumpolar structure and distribution of the Antarctic Circumpolar Current fronts: 2. Variability and relationship to sea surface height. J. Geophys. Res. 2009, 114, C11019. [Google Scholar] [CrossRef] [Green Version]
- Good, S.; Fiedler, E.; Mao, C.; Martin, M.J.; Maycock, A.; Reid, R.; Roberts-Jones, J.; Searle, T.; Waters, J.; While, J.; et al. The current configuration of the OSTIA system for operational production of foundation sea surface temperature and ice concentration analyses. Remote Sens. 2020, 12, 720. [Google Scholar] [CrossRef] [Green Version]
- Hersbach, H.; Bell, B.; Berrisford, P.; Biavati, G.; Horányi, A.; Muñoz Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Rozum, I.; et al. ERA5 hourly Data on Pressure Levels from 1979 to Present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). 2018. Available online: https://cds.climate.copernicus.eu/cdsapp#!/dataset/10.24381/cds.bd0915c6?tab=overview (accessed on 20 January 2021).
- Trenberth, K.E.; Large, W.G.; Olson, J.G. The mean annual cycle in global ocean wind stress. J. Phys. Oceanogr. 1990, 20, 1742–1760. [Google Scholar] [CrossRef] [Green Version]
- Risien, C.M.; Chelton, D.B. A global climatology of surface wind and wind stress fields from eight years of QuikSCAT scatterometer data. J. Phys. Oceanogr. 2008, 38, 2379–2413. [Google Scholar] [CrossRef]
- Holte, J.; Talley, L.D.; Gilson, J.; Roemmich, D. An Argo mixed layer climatology and database. Geophys. Res. Lett. 2017, 44, 5618–5626. [Google Scholar] [CrossRef] [Green Version]
- Zuo, H.; Balmaseda, M.A.; Tietsche, S.; Mogensen, K.; Mayer, M. The ECMWF operational ensemble reanalysis–analysis system for ocean and sea ice: A description of the system and assessment. Ocean Sci. 2019, 15, 779–808. [Google Scholar] [CrossRef] [Green Version]
- Lellouche, J.-M.; Le Galloudec, O.; Drévillon, M.; Régnier, C.; Greiner, E.; Garric, G.; Ferry, N.; Desportes, C.; Testut, C.-E.; Bricaud, C.; et al. Evaluation of global monitoring and forecasting systems at Mercator Océan. Ocean Sci. 2013, 9, 57–81. [Google Scholar] [CrossRef] [Green Version]
- Storto, A.; Masina, S.; Navarra, A. Evaluation of the CMCC eddy-permitting global ocean physical reanalysis system (C-GLORS, 1982-2012) and its assimilation components. Q. J. R. Meteorol. Soc. 2016, 142, 738–758. [Google Scholar] [CrossRef]
- MacLachlan, C.; Arribas, A.; Peterson, K.A.; Maidens, A.; Fereday, D.; Scaife, A.A.; Gordon, M.; Vellinga, M.; Williams, A.; Comer, R.E.; et al. Global seasonal forecast system version 5 (GloSea5): A high-resolution seasonal forecast system. Q. J. R. Meteorol. Soc. 2015, 141, 1072–1084. [Google Scholar] [CrossRef]
- Korb, R.E.; Whitehouse, M.J.; Atkinson, A.; Thorpe, S.E. Magnitude and maintenance of the phytoplankton bloom at South Georgia: A naturally iron-replete environment. Mar. Ecol. Prog. Ser. 2008, 368, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Venables, H.J.; Pollard, R.T.; Popova, E.E. Physical conditions controlling the development of a regular phytoplankton bloom north of the Crozet Plateau, Southern Ocean. Deep-Sea Res. II 2007, 54, 1949–1965. [Google Scholar] [CrossRef]
- Rouault, M.; Mélice, J.-L.; Reason, C.J.C.; Lutjeharms, J.R.E. Climate variability at Marion Island, Southern Ocean, since 1960. J. Geophys. Res. 2005, 110, C05007. [Google Scholar] [CrossRef] [Green Version]
- Rintoul, S.R. The global influence of localized dynamics in the Southern Ocean. Nature 2018, 558, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Whitehouse, M.J.; Priddle, J.; Cripps, G.C.; Ward, P.; Brandon, M.A. South Georgia, Antarctica: A productive, cold water, pelagic ecosystem. Mar. Ecol. Prog. Ser. 2001, 216, 279–308. [Google Scholar] [CrossRef] [Green Version]
- Korb, R.E.; Whitehouse, M.J.; Ward, P. SeaWiFS in the Southern Ocean: Spatial and temporal variability in phytoplankton biomass around South Georgia. Deep-Sea Res. II 2004, 51, 99–116. [Google Scholar] [CrossRef]
- Moore, J.K.; Abbott, M.R. Phytoplankton chlorophyll distributions and primary production in the Southern Ocean. J. Geophys. Res. 2000, 105, 28709–28722. [Google Scholar] [CrossRef]
- Hasegawa, D.; Yamazaki, H.; Ishimaru, T.; Nagashima, H.; Koike, Y. Apparent phytoplankton bloom due to island mass effect. J. Mar. Syst. 2008, 69, 238–246. [Google Scholar] [CrossRef]
- Hasegawa, D.; Lewis, M.R.; Gangopadhyay, A. How islands cause phytoplankton to bloom in their wakes. Geophys. Res. Lett. 2009, 36, L20605. [Google Scholar] [CrossRef]
- Messié, M.; Petrenko, A.; Doglioli, A.M.; Aldebert, C.; Martinez, E.; Koenig, G.; Bonnet, S.; Moutin, T. The delayed Island Mass Effect: How islands can remotely trigger blooms in the oligotrophic ocean. Geophys. Res. Lett. 2020, 47, e2019GL085282. [Google Scholar] [CrossRef] [Green Version]
- Blain, S.; Quéguiner, B.; Armand, L.; Belviso, S.; Bombled, B.; Bopp, L.; Bowie, A.; Brunet, C.; Brussard, C.; Carlottie, F.; et al. Effect of natural iron fertilization on carbon sequestration in the Southern Ocean. Nature 2007, 446, 7139. [Google Scholar] [CrossRef] [PubMed]
- Swart, S.; Thomalla, S.J.; Monteiro, P.M.S. The seasonal cycle of mixed layer dynamics and phytoplankton biomass in the Sub-Antarctic Zone: A high-resolution glider experiment. J. Mar. Syst. 2015, 147, 103–115. [Google Scholar] [CrossRef]
- Arteaga, L.A.; Boss, E.; Behrenfeld, M.J.; Westberry, T.K.; Sarmiento, J.L. Seasonal modulation of phytoplankton biomass in the Southern Ocean. Nat. Commun. 2020, 11, 5364. [Google Scholar] [CrossRef] [PubMed]
- Ratnarajah, L.; Blain, S.; Boyd, P.W.; Fourquez, M.; Obernosterer, I.; Tagliabue, A. Resource colimitation drives competition between phytoplankton and bacteria in the Southern Ocean. Geophys. Res. Lett. 2021, 48, e2020GL088369. [Google Scholar] [CrossRef] [PubMed]
- Behrenfeld, M.J.; O’Malley, R.T.; Siegel, D.A.; McClain, C.R.; Sarmiento, J.L.; Feldman, G.C.; Milligan, A.J.; Falkowski, P.G.; Letelier, R.M.; Boss, E.S. Climate-driven trends in contemporary ocean productivity. Nature 2006, 444, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Boyce, D.G.; Lewis, M.R.; Worm, B. Global phytoplankton decline over the past century. Nature 2010, 466, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Gregg, W.W.; Casey, N.W.; McClain, C.R. Recent trends in global ocean chlorophyll. Geophys. Res. Lett. 2005, 32, L03606. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Gao, L.; Xu, Y. Quantification of the impact of environmental factors on chlorophyll in the open ocean. J. Oceanol. Limnol. 2021, 39, 447–457. [Google Scholar] [CrossRef]
- Dunstan, P.K.; Foster, S.D.; King, E.; Risbey, J.; O’Kane, T.J.; Monselesan, D.; Hobday, A.J.; Hartog, J.R.; Thompson, P.A. Global patterns of change and variation in sea surface temperature and chlorophyll a. Sci. Rep. 2018, 8, 14624. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Durant, J.M.; Stige, L.C.; Hessen, D.O.; Hjermann, D.Ø.; Zhu, L.; Llope, M.; Stenseth, N.C. Contrasting correlation patterns between environmental factors and chlorophyll levels in the global ocean. Global Biogeochem. Cycles 2015, 29, 2095–2107. [Google Scholar] [CrossRef]
- Sarmiento, J.L.; Slater, R.; Barber, R.; Bopp, L.; Doney, S.C.; Hirst, A.C.; Kleypas, J.; Matear, R.; Mikolajewicz, U.; Monfray, P.; et al. Response of ocean ecosystems to climate warming. Global Biogeochem. Cycles 2004, 18, GB3003. [Google Scholar] [CrossRef] [Green Version]
- Boyd, P.W.; Dilllingham, P.W.; McGraw, C.M.; Armstrong, E.A.; Cornwall, C.E.; Feng, Y.-Y.; Hurd, C.L.; Gault-Ringold, M.; Roleda, M.Y.; Timmins-Schiffman, E.; et al. Physiological responses of a Southern Ocean diatom to complex future ocean conditions. Nat. Clim. Chang. 2016, 6, 207–213. [Google Scholar] [CrossRef]
- Montie, S.; Thomsen, M.S.; Rack, W.; Broady, P.A. Extreme summer marine heatwaves increase chlorophyll a in the Southern Ocean. Antarct. Sci. 2020, 32, 508–509. [Google Scholar] [CrossRef]
- Smith, V.R.; Froneman, P.W. 2008. Nutrient dynamics in the vicinity of the Prince Edward Islands. In The Prince Edward Islands: Land-Sea Interactions in a Changing Ecosystem; Chown, S.L., Froneman, P.W., Eds.; Sun Press: Stellenbosch, South Africa, 2008; pp. 165–179. [Google Scholar]
- Closset, I.; Lasbleiz, M.; Leblanc, K.; Quéguiner, B.; Cavanga, A.-J.; Elskens, M.; Navez, J.; Cardinal, D. Seasonal evolution of net and regenerated silica production around a natural Fe-fertilized area in the Southern Ocean estimated with Si isotopic approaches. Biogeosciences 2014, 11, 5827–5846. [Google Scholar] [CrossRef] [Green Version]
- Pollard, R.T.; Venables, H.J.; Read, J.F.; Allen, J.T. Large-scale circulation around the Crozet Plateau controls an annual phytoplankton bloom in the Crozet Basin. Deep-Sea Res. II 2007, 54, 1915–1929. [Google Scholar] [CrossRef]
- Puccinelli, E.; Smart, S.M.; Fawcett, S.E. Temporal variability in the trophic composition of benthic invertebrates in the Indian sub-Antarctic Ocean. Deep-Sea Res. I 2020, 163, 103340. [Google Scholar] [CrossRef]
- Fourquez, M.; Bressac, M.; Deppeler, S.L.; Ellwood, M.; Obernosterer, I.; Trull, T.W.; Boyd, P.W. Microbial competition in the subpolar Southern Ocean: An Fe-C co-limitation experiment. Front. Mar. Sci. 2020, 6, 776. [Google Scholar] [CrossRef] [Green Version]
- Planquette, H.; Statham, P.J.; Fones, G.R.; Charette, M.A.; Moore, C.M.; Salter, I.; Nédélec, F.H.; Taylor, S.L.; French, M.; Baker, A.R.; et al. Dissolved iron in the vicinity of the Crozet Islands, Southern Ocean. Deep-Sea Res. II 2007, 54, 1999–2019. [Google Scholar] [CrossRef] [Green Version]
- Carranza, M.M.; Gille, S.T. Southern Ocean wind-driven entrainment enhances satellite chlorophyll-a through the summer. J. Geophys. Res. Oceans 2015, 120, 304–323. [Google Scholar] [CrossRef] [Green Version]
- Kahru, M.; Gille, S.T.; Murtugudde, R.; Strutton, P.G.; Manzano-Sarabia, M.; Wang, H.; Mitchell, B.G. Global correlations between winds and ocean chlorophyll. J. Geophys. Res. 2010, 115, C12040. [Google Scholar] [CrossRef] [Green Version]
- Pellichero, V.; Boutin, J.; Claustre, H.; Merlivat, L.; Sallée, J.-B.; Blain, S. Relaxation of wind stress drives the abrupt onset of biological carbon uptake in the Kergeulen bloom: A multisensory approach. Geophys. Res. Lett. 2020, 47, e2019GL085992. [Google Scholar] [CrossRef]
- Irion, S.; Jardillier, L.; Sassenhagen, I.; Christaki, U. Marked spatiotemporal variations in small phytoplankton structure in contrasted waters of the Southern Ocean (Kerguelen area). Limnol. Oceanogr. 2020, 65, 2835–2852. [Google Scholar] [CrossRef]
- Nikurashin, M.; Ferrari, R. Overturning circulation driven by breaking internal waves in the deep ocean. Geophys. Res. Lett. 2013, 40, 3133–3137. [Google Scholar] [CrossRef]
- Dawson, H.R.S.; Strutton, P.G.; Gaube, P. The unusual surface chlorophyll signatures of Southern Ocean eddies. J. Geophys. Res. Oceans 2018, 123, 6053–6069. [Google Scholar] [CrossRef]
- Frenger, I.; Münnich, M.; Gruber, N. Imprint of Southern Ocean mesoscale eddies on chlorophyll. Biogeosciences 2018, 15, 4781–4798. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamont, T.; Toolsee, T. Spatial and Seasonal Variations of the Island Mass Effect at the Sub-Antarctic Prince Edward Islands Archipelago. Remote Sens. 2022, 14, 2140. https://doi.org/10.3390/rs14092140
Lamont T, Toolsee T. Spatial and Seasonal Variations of the Island Mass Effect at the Sub-Antarctic Prince Edward Islands Archipelago. Remote Sensing. 2022; 14(9):2140. https://doi.org/10.3390/rs14092140
Chicago/Turabian StyleLamont, Tarron, and Tesha Toolsee. 2022. "Spatial and Seasonal Variations of the Island Mass Effect at the Sub-Antarctic Prince Edward Islands Archipelago" Remote Sensing 14, no. 9: 2140. https://doi.org/10.3390/rs14092140