
Airborne HySpex Hyperspectral Versus Multitemporal Sentinel-2 Images for Mountain Plant Communities Mapping




Abstract
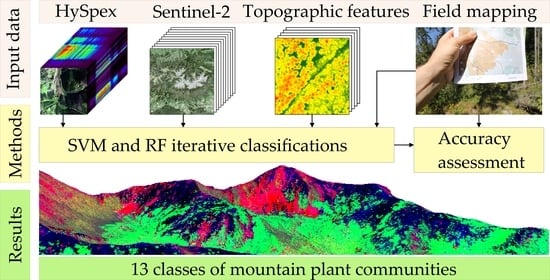
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Research Schema
2.3. Airborne and Satellite Input Data
2.4. Reference Data, Classification, and Accuracy Assessment
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 50 | 100 | 200 | 300 | 500 | 700 | 1000 | ||
|---|---|---|---|---|---|---|---|---|
| HySpex + TF | RF | 79.6 | 83.6 | 86.4 | 87.6 | 88.9 | 89.6 | 89.9 |
| SVM | 80.0 | 83.8 | 86.0 | 87.0 | 88.1 | 88.6 | 89.3 | |
| HySpex | RF | 66.1 | 69.4 | 72.1 | 73.1 | 74.4 | 75.3 | 75.9 |
| SVM | 69.1 | 72.8 | 75.4 | 76.7 | 77.7 | 78.5 | 79.3 | |
| 30 MNF bands + TF | RF | 83.5 | 85.7 | 87.4 | 87.8 | 88.7 | 88.8 | 89.0 |
| SVM | 74.1 | 77.2 | 79.2 | 80.2 | 81.0 | 81.0 | 81.9 | |
| 30 MNFbands | RF | 66.2 | 69.5 | 72.0 | 73.2 | 74.7 | 75.6 | 76.4 |
| SVM | 58.4 | 62.5 | 65.7 | 67.2 | 68.8 | 69.7 | 70.5 | |
| Sentinel-2 + TF | RF | 86.9 | 87.9 | 88.6 | 88.7 | 88.8 | 88.9 | 88.9 |
| SVM | 86.6 | 87.6 | 88.1 | 88.3 | 88.3 | 88.5 | 88.4 | |
| Sentinel-2 | RF | 81.8 | 82.8 | 83.5 | 83.4 | 83.5 | 83.5 | 83.5 |
| SVM | 83.1 | 84.8 | 85.5 | 85.8 | 86.1 | 86.0 | 86.1 |
References
- Malanson, G.P.; Resler, L.M.; Butler, D.R.; Fagre, D.B. Mountain plant communities: Uncertain sentinels? Prog. Phys. Geogr. Earth Environ. 2019, 43, 521–543. [Google Scholar] [CrossRef]
- Zhang, J.; Yao, Y.; Suo, N. Automatic classification of fine-scale mountain vegetation based on mountain altitudinal belt. PLoS ONE 2020, 15, e0238165. [Google Scholar] [CrossRef] [PubMed]
- Canedoli, C.; Ferrè, C.; Abu El Khair, D.; Comolli, R.; Liga, C.; Mazzucchelli, F.; Proietto, A.; Rota, N.; Colombo, G.; Bassano, B.; et al. Evaluation of ecosystem services in a protected mountain area: Soil organic carbon stock and biodiversity in alpine forests and grasslands. Ecosyst. Serv. 2020, 44, 101135. [Google Scholar] [CrossRef]
- Kycko, M.; Zagajewski, B.; Lavender, S.; Romanowska, E.; Zwijacz-Kozica, M. The impact of tourist traffic on the condition and cell structures of alpine swards. Remote Sens. 2018, 10, 220. [Google Scholar] [CrossRef] [Green Version]
- Migas-Mazur, R.; Kycko, M.; Zwijacz-Kozica, T.; Zagajewski, B. Assessment of sentinel-2 images, support vector machines and change detection algorithms for bark beetle outbreaks mapping in the tatra mountains. Remote Sens. 2021, 13, 3314. [Google Scholar] [CrossRef]
- Allan, E.; Weisser, W.; Weigelt, A.; Roscher, C.; Fischer, M.; Hillebrand, H. More diverse plant communities have higher functioning over time due to turnover in complementary dominant species. Proc. Natl. Acad. Sci. USA 2011, 108, 17034–17039. [Google Scholar] [CrossRef] [Green Version]
- Burdon, J.J.; Zhan, J. Climate change and disease in plant communities. PLoS Biol. 2020, 18, e3000949. [Google Scholar] [CrossRef]
- Cierniewski, J.; Kazmierowski, C.; Krolewicz, S.; Piekarczyk, J.; Wrobel, M.; Zagajewski, B. Effects of different illumination and observation techniques of cultivated soils on their hyperspectral bidirectional measurements under field and laboratory conditions. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 2525–2530. [Google Scholar] [CrossRef]
- Kycko, M.; Zagajewski, B.; Lavender, S.; Dabija, A. In situ hyperspectral remote sensing for monitoring of alpine trampled and recultivated species. Remote Sens. 2019, 11, 1296. [Google Scholar] [CrossRef] [Green Version]
- Raczko, E.; Zagajewski, B. Tree species classification of the UNESCO man and the biosphere karkonoski national park (Poland) using artificial neural networks and APEX hyperspectral images. Remote Sens. 2018, 10, 1111. [Google Scholar] [CrossRef] [Green Version]
- Zagajewski, B.; Kluczek, M.; Raczko, E.; Njegovec, A.; Dabija, A.; Kycko, M. Comparison of random forest, support vector machines, and neural networks for post-disaster forest species mapping of the Krkonoše/Karkonosze transboundary biosphere reserve. Remote Sens. 2021, 13, 2581. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Yu, Q.; Zimmerman, M.J.; Flint, S.; Waldron, M.C. Differentiating aquatic plant communities in a eutrophic river using hyperspectral and multispectral remote sensing. Freshw. Biol. 2010, 55, 1658–1673. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Gill, L.; Regan, S.; Naughton, O.; Johnston, P.; Waldren, S.; Ghosh, B. Mapping vegetation communities inside wetlands using sentinel-2 imagery in Ireland. Int. J. Appl. Earth Obs. Geoinf. 2020, 88, 102083. [Google Scholar] [CrossRef]
- Rapinel, S.; Rossignol, N.; Hubert-Moy, L.; Bouzillé, J.-B.; Bonis, A. Mapping grassland plant communities using a fuzzy approach to address floristic and spectral uncertainty. Appl. Veg. Sci. 2018, 21, 678–693. [Google Scholar] [CrossRef]
- Zagajewski, B.; Tømmervik, H.; Bjerke, J.; Raczko, E.; Bochenek, Z.; Kłos, A.; Jarocińska, A.; Lavender, S.; Ziółkowski, D. Intraspecific differences in spectral reflectance curves as indicators of reduced vitality in high-arctic plants. Remote Sens. 2017, 9, 1289. [Google Scholar] [CrossRef] [Green Version]
- Marcinkowska-Ochtyra, A.; Zagajewski, B.; Raczko, E.; Ochtyra, A.; Jarocińska, A. Classification of high-mountain vegetation communities within a diverse giant mountains ecosystem using airborne apex hyperspectral imagery. Remote Sens. 2018, 10, 570. [Google Scholar] [CrossRef] [Green Version]
- De Giglio, M.; Greggio, N.; Goffo, F.; Merloni, N.; Dubbini, M.; Barbarella, M. Comparison of pixel- and object-based classification methods of unmanned aerial vehicle data applied to coastal dune vegetation communities: Casal borsetti case study. Remote Sens. 2019, 11, 1416. [Google Scholar] [CrossRef] [Green Version]
- Laporte-Fauret, Q.; Lubac, B.; Castelle, B.; Michalet, R.; Marieu, V.; Bombrun, L.; Launeau, P.; Giraud, M.; Normandin, C.; Rosebery, D. Classification of atlantic coastal sand dune vegetation using in situ, UAV, and airborne hyperspectral data. Remote Sens. 2020, 12, 2222. [Google Scholar] [CrossRef]
- Kollert, A.; Bremer, M.; Löw, M.; Rutzinger, M. Exploring the potential of land surface phenology and seasonal cloud free composites of one year of Sentinel-2 imagery for tree species mapping in a mountainous region. Int. J. Appl. Earth Obs. Geoinf. 2021, 94, 102208. [Google Scholar] [CrossRef]
- Harrison, S. Plant community diversity will decline more than increase under climatic warming. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190106. [Google Scholar] [CrossRef] [Green Version]
- Raczko, E.; Zagajewski, B. Comparison of support vector machine, random forest and neural network classifiers for tree species classification on airborne hyperspectral APEX images. Eur. J. Remote Sens. 2017, 50, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Brodrick, P.G.; Davies, A.B.; Asner, G.P. Uncovering ecological patterns with convolutional neural networks. Trends Ecol. Evol. 2019, 34, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, A.; Clarke, K.; Lewis, M. Hyperspectral classification of plants: A review of waveband selection generalisability. Remote Sens. 2020, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Kycko, M.; Zagajewski, B.; Zwijacz-Kozica, M.; Cierniewski, J.; Romanowska, E.; Orłowska, K.; Ochtyra, A.; Jarocińska, A. Assessment of hyperspectral remote sensing for analyzing the impact of human trampling on alpine swards. Mt. Res. Dev. 2017, 37, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Zagajewski, B.; Kycko, M.; Tømmervik, H.; Bochenek, Z.; Wojtuń, B.; Bjerke, J.W.; Kłos, A. Feasibility of hyperspectral vegetation indices for the detection of chlorophyll concentration in three high Arctic plants: Salix polaris, Bistorta vivipara, and Dryas octopetala. Acta Soc. Bot. Pol. 2018, 87, 3604. [Google Scholar] [CrossRef]
- Jia, J.; Wang, Y.; Chen, J.; Guo, R.; Shu, R.; Wang, J. Status and application of advanced airborne hyperspectral imaging technology: A review. Infrared Phys. Technol. 2020, 104, 103115. [Google Scholar] [CrossRef]
- Marcinkowska-Ochtyra, A.; Zagajewski, B.; Ochtyra, A.; Jarocińska, A.; Wojtuń, B.; Rogass, C.; Mielke, C.; Lavender, S. Subalpine and alpine vegetation classification based on hyperspectral APEX and simulated EnMAP images. Int. J. Remote Sens. 2017, 38, 1839–1864. [Google Scholar] [CrossRef] [Green Version]
- Sabat-Tomala, A.; Raczko, E.; Zagajewski, B. Comparison of support vector machine and random forest algorithms for invasive and expansive species classification using airborne hyperspectral data. Remote Sens. 2020, 12, 516. [Google Scholar] [CrossRef] [Green Version]
- Mäyrä, J.; Keski-Saari, S.; Kivinen, S.; Tanhuanpää, T.; Hurskainen, P.; Kullberg, P.; Poikolainen, L.; Viinikka, A.; Tuominen, S.; Kumpula, T.; et al. Tree species classification from airborne hyperspectral and LiDAR data using 3D convolutional neural networks. Remote Sens. Environ. 2021, 256, 112322. [Google Scholar] [CrossRef]
- Yao, Y.; Suonan, D.; Zhang, J. Compilation of 1:50,000 vegetation type map with remote sensing images based on mountain altitudinal belts of Taibai Mountain in the North-South transitional zone of China. J. Geogr. Sci. 2020, 30, 267–280. [Google Scholar] [CrossRef]
- Mellor, A.; Haywood, A.; Stone, C.; Jones, S. The Performance of random forests in an operational setting for large area sclerophyll forest classification. Remote Sens. 2013, 5, 2838–2856. [Google Scholar] [CrossRef] [Green Version]
- Fei, H.; Fan, Z.; Wang, C.; Zhang, N.; Wang, T.; Chen, R.; Bai, T. Cotton classification method at the county scale based onmulti-features and random forest feature selection algorithm and classifier. Remote Sens. 2022, 14, 829. [Google Scholar] [CrossRef]
- Ochtyra, A. Forest disturbances in polish Tatra mountains for 1985–2016 in relation to topography, stand features, and protection zone. Forests 2020, 11, 579. [Google Scholar] [CrossRef]
- Kattenborn, T.; Eichel, J.; Wiser, S.; Burrows, L.; Fassnacht, F.E.; Schmidtlein, S. Convolutional Neural Networks accurately predict cover fractions of plant species and communities in Unmanned Aerial Vehicle imagery. Remote Sens. Ecol. 2020, 6, 472–486. [Google Scholar] [CrossRef] [Green Version]
- Hamylton, S.M.; Morris, R.H.; Carvalho, R.C.; Roder, N.; Barlow, P.; Mills, K.; Wang, L. Evaluating techniques for mapping island vegetation from unmanned aerial vehicle (UAV) images: Pixel classification, visual interpretation and machine learning approaches. Int. J. Appl. Earth Obs. Geoinf. 2020, 89, 102085. [Google Scholar] [CrossRef]
- Kattenborn, T.; Eichel, J.; Fassnacht, F.E. Convolutional Neural Networks enable efficient, accurate and fine-grained segmentation of plant species and communities from high-resolution UAV imagery. Sci. Rep. 2019, 9, 17656. [Google Scholar] [CrossRef]
- Xiao, H.; Su, F.; Fu, D.; Lyne, V.; Liu, G.; Pan, T.; Teng, J. Optimal and robust vegetation mapping in complex environments using multiple satellite imagery: Application to mangroves in Southeast Asia. Int. J. Appl. Earth Obs. Geoinf. 2021, 99, 102320. [Google Scholar] [CrossRef]
- Sothe, C.; Almeida, C.M.D.; Liesenberg, V.; Schimalski, M.B. Evaluating Sentinel-2 and Landsat-8 Data to Map Sucessional Forest Stages in a Subtropical Forest in Southern Brazil. Remote Sens. 2017, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Toulemont, A.; Fernandez, V.; Mavrocordatos, C.; Mandorlo, G.; Laberinti, P. Sentinel-2C instrument new features and first instrument performance characterization. In Proceedings of the Sensors, Systems, and Next-Generation Satellites XXIII, Strasbourg, France, 9–12 September 2019; Neeck, S.P., Kimura, T., Martimort, P., Eds.; SPIE: Bellingham, DC, USA, 2019; p. 25. [Google Scholar] [CrossRef]
- Toulemont, A.; Olivier, M.; Clerc, S.; Bellouard, R.; Reina, F.; Gascon, F.; Luce, J.-F.; Mavrocordatos, C.; Boccia, V. Copernicus Sentinel-2C/D Multi Spectral Instrument full field of view spectral characterization. In Proceedings of the Sensors, Systems, and Next-Generation Satellites XXV, Online, 13–18 September 2021; Neeck, S.P., Kimura, T., Babu, S.R., Hélière, A., Eds.; SPIE: Bellingham, DC, USA, 2021; p. 26. [Google Scholar] [CrossRef]
- Chrysafis, I.; Korakis, G.; Kyriazopoulos, A.P.; Mallinis, G. Predicting Tree Species Diversity Using Geodiversity and Sentinel-2 Multi-Seasonal Spectral Information. Sustainability 2020, 12, 9250. [Google Scholar] [CrossRef]
- Shukla, G.; Garg, R.D.; Kumar Garg, P.; Srivastava, H.S.; Kumar, P.; Mohanty, B. Exploring the capabilities of sentinel-2 data in vegetation health/stress mapping. In Proceedings of the IGARSS 2019—2019 IEEE International Geoscience and Remote Sensing Symposium, Yokohama, Japan, 28 July–2 August 2019; pp. 6652–6655. [Google Scholar] [CrossRef]
- Tarantino, C.; Forte, L.; Blonda, P.; Vicario, S.; Tomaselli, V.; Beierkuhnlein, C.; Adamo, M. Intra-annual sentinel-2 time-series supporting grassland habitat discrimination. Remote Sens. 2021, 13, 277. [Google Scholar] [CrossRef]
- Kwan, C.; Gribben, D.; Ayhan, B.; Li, J.; Bernabe, S.; Plaza, A. An accurate vegetation and non-vegetation differentiation approach based on land cover classification. Remote Sens. 2020, 12, 3880. [Google Scholar] [CrossRef]
- Kwon, H.; Nasrabadi, N.M. Kernel matched subspace detectors for hyperspectral target detection. IEEE Trans. Pattern Anal. Mach. Intell. 2006, 28, 178–194. [Google Scholar] [CrossRef]
- Khodadadzadeh, M.; Li, J.; Prasad, S.; Plaza, A. Fusion of hyperspectral and LiDAR remote sensing data using multiple feature learning. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 2971–2983. [Google Scholar] [CrossRef]
- Li, M.; Stein, A. Mapping land use from high resolution satellite images by exploiting the spatial arrangement of land cover objects. Remote Sens. 2020, 12, 4158. [Google Scholar] [CrossRef]
- Zagajewski, B. Assessment of neural networks and imaging spectroscopy for vegetation classification of the high Tatras. Teledetekcja Sr. 2010, 43, 113. [Google Scholar]
- Grabska, E.; Hostert, P.; Pflugmacher, D.; Ostapowicz, K. Forest stand species mapping using the sentinel-2 time series. Remote Sens. 2019, 11, 1197. [Google Scholar] [CrossRef] [Green Version]
- Wakulińska, M.; Marcinkowska-Ochtyra, A. Multi-temporal sentinel-2 data in classification of mountain vegetation. Remote Sens. 2020, 12, 2696. [Google Scholar] [CrossRef]
- Cierniewski, J.; Ceglarek, J.; Karnieli, A.; Królewicz, S.; Kaźmierowski, C.; Zagajewski, B. Predicting the diurnal blue-sky albedo of soils using their laboratory reflectance spectra and roughness indices. J. Quant. Spectrosc. Radiat. Transf. 2017, 200, 25–31. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Knollová, I.; Janssen, J.A.M.; Rodwell, J.S.; Peterka, T.; Marcenò, C.; Landucci, F.; Danihelka, J.; et al. EUNIS Habitat Classification: Expert system, characteristic species combinations and distribution maps of European habitats. Appl. Veg. Sci. 2020, 23, 648–675. [Google Scholar] [CrossRef]
- Dabija, A.; Kluczek, M.; Zagajewski, B.; Raczko, E.; Kycko, M.; Al-Sulttani, A.H.; Tardà, A.; Pineda, L.; Corbera, J. Comparison of support vector machines and random forests for corine land cover mapping. Remote Sens. 2021, 13, 777. [Google Scholar] [CrossRef]
- Zięba, A.; Wróbel, S.; Kauzal, P.; Delimat, A.; Ociepa, A.M.; Kozak, M.; Kozłowska-Kozak, K.; Czortek, P.; Baran, J.; Bryniarski, G.; et al. Przyczynki do flory Tatrzańskiego Parku Narodowego. Fragm. Florist. Geobot. Pol. 2020, 379–394. [Google Scholar] [CrossRef]
- Bodziarczyk, J.; Szwagrzyk, J.; Zwijacz-Kozica, T.; Zięba, A.; Szewczyk, J.; Gazda, A. The structure of forest stands in the Tatra National Park: The results of 2016–2017 inventory. For. Res. Pap. 2019, 80, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Czortek, P.; Kapfer, J.; Delimat, A.; Eycott, A.E.; Grytnes, J.-A.; Orczewska, A.; Ratyńska, H.; Zięba, A.; Jaroszewicz, B. Plant species composition shifts in the Tatra Mts as a response to environmental change: A resurvey study after 90 years. Folia Geobot. 2018, 53, 333–348. [Google Scholar] [CrossRef]
- Łupikasza, E.; Szypuła, B. Vertical climatic belts in the Tatra Mountains in the light of current climate change. Theor. Appl. Climatol. 2019, 136, 249–264. [Google Scholar] [CrossRef]
- Górnik, M.; Holko, L.; Pociask-Karteczka, J.; Bičárová, S. Variability of precipitation and runoff in the entire high Tatra Mountains in the period 1961–2010. Pr. Geogr. 2017, 151, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Mezei, P.; Jakuš, R.; Pennerstorfer, J.; Havašová, M.; Škvarenina, J.; Ferenčík, J.; Slivinský, J.; Bičárová, S.; Bilčík, D.; Blaženec, M.; et al. Storms, temperature maxima and the Eurasian spruce bark beetle Ips typographus—An infernal trio in Norway spruce forests of the Central European High Tatra Mountains. Agric. For. Meteorol. 2017, 242, 85–95. [Google Scholar] [CrossRef]
- Sproull, G.J.; Bukowski, M.; Mcnutt, N.; Zwijacz-Kozica, T.; Szwagrzyk, J. Landscape-level spruce mortality patterns and topographic forecasters of bark beetle outbreaks in managed and unmanaged forests of the Tatra mountains. Pol. J. Ecol. 2017, 65, 24–37. [Google Scholar] [CrossRef]
- Delimat, A.; Kiełtyk, P. Impact of troublesome expansive weed Rumex alpinus on species diversity of mountain pastures in Tatra National Park, Poland. Biologia 2019, 74, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Kiełtyk, P.; Delimat, A. Impact of the alien plant Impatiens glandulifera on species diversity of invaded vegetation in the northern foothills of the Tatra Mountains, Central Europe. Plant Ecol. 2019, 220, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Richter, R.; Schläpfer, D. Atmospheric/Topographic Correction for Satellite Imagery; ATCOR-2/3 User Guide, Version 9.0.2 (DLR-IB565-01/15); DLR—German Aerospace Center: Wessling, Germany; ReSe Applications: Wil, Switzerland, 2016; p. 263. [Google Scholar]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.3-13. 2020. Available online: https://rdrr.io/cran/raster/ (accessed on 10 December 2021).
- Ranghetti, L.; Boschetti, M.; Nutini, F.; Busetto, L. “sen2r”: An R toolbox for automatically downloading and preprocessing Sentinel-2 satellite data. Comput. Geosci. 2020, 139, 104473. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 23 April 2021).
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library; R Package Version 1.5-12. 2020. Available online: https://rdrr.io/cran/rgdal/ (accessed on 25 April 2020).
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Vapnik, V.N. The Nature of Statistical Learning Theory; Springer: New York, NY, USA, 1995; p. 314. ISBN 978-1-4757-3264-1. [Google Scholar] [CrossRef]
- Vapnik, V.N. An overview of statistical learning theory. IEEE Trans. Neural Netw. 1999, 10, 988–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehman, S.V.; Foody, G.M. Key issues in rigorous accuracy assessment of land cover products. Remote Sens. Environ. 2019, 231, 111199. [Google Scholar] [CrossRef]
- Sabat-Tomala, A.; Raczko, E.; Zagajewski, B. Mapping invasive plant species with hyperspectral data based on iterative accuracy assessment techniques. Remote Sens. 2022, 14, 64. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Lillesand, T.; Kiefer, R.W.; Chipman, J. Remote Sensing and Image Interpretation, 7th ed.; John Wiley and Sons: Hoboken, NJ, USA, 2015; p. 736. [Google Scholar]
- Van Rijsbergen, C.J. Information Retrieval, 2nd ed.; Butterworth-Heinemann: Newton, MA, USA, 1979; p. 208. [Google Scholar]
- Sasaki, Y. The truth of the F-measure. Teach. Tutor. Mater. 2007, 1, 1–5. [Google Scholar]
- Shao, G.; Tang, L.; Liao, J. Overselling overall map accuracy misinforms about research reliability. Landsc. Ecol. 2019, 34, 2487–2492. [Google Scholar] [CrossRef] [Green Version]
- Stehman, S.V.; Wickham, J. A guide for evaluating and reporting map data quality: Affirming Shao et al. “Overselling overall map accuracy misinforms about research reliability”. Landsc. Ecol. 2020, 35, 1263–1267. [Google Scholar] [CrossRef]
- Foody, G.M. Explaining the unsuitability of the kappa coefficient in the assessment and comparison of the accuracy of thematic maps obtained by image classification. Remote Sens. Environ. 2020, 239, 111630. [Google Scholar] [CrossRef]
- Brell, M.; Segl, K.; Guanter, L.; Bookhagen, B. Hyperspectral and lidar intensity data fusion: A framework for the rigorous correction of illumination, anisotropic effects, and cross calibration. IEEE Trans. Geosci. Remote Sens. 2017, 55, 2799–2810. [Google Scholar] [CrossRef] [Green Version]
- Lucas, R.; Bunting, P.; Paterson, M.; Chisholm, L. Classification of Australian forest communities using aerial photography, CASI and HyMap data. Remote Sens. Environ. 2008, 112, 2088–2103. [Google Scholar] [CrossRef]
- McPartland, M.Y.; Falkowski, M.J.; Reinhardt, J.R.; Kane, E.S.; Kolka, R.; Turetsky, M.R.; Douglas, T.A.; Anderson, J.; Edwards, J.D.; Palik, B.; et al. Characterizing boreal peatland plant composition and species diversity with hyperspectral remote sensing. Remote Sens. 2019, 11, 1685. [Google Scholar] [CrossRef] [Green Version]
- Deepa, P.; Thilagavathi, K. Data reduction techniques of hyperspectral images: A comparative study. In Proceedings of the 2015 3rd International Conference on Signal Processing, Communication and Networking (ICSCN), Chennai, India, 26–28 March 2015; pp. 1–6. [Google Scholar] [CrossRef]
- Hasanlou, M.; Samadzadegan, F. Comparative study of intrinsic dimensionality estimation and dimension reduction techniques on hyperspectral images using K-NN classifier. IEEE Geosci. Remote Sens. Lett. 2012, 9, 1046–1050. [Google Scholar] [CrossRef]
- Kopeć, D.; Sabat-Tomala, A.; Michalska-Hejduk, D.; Jarocińska, A.; Niedzielko, J. Application of airborne hyperspectral data for mapping of invasive alien Spiraea tomentosa L.: A serious threat to peat bog plant communities. Wetl. Ecol. Manag. 2020, 28, 357–373. [Google Scholar] [CrossRef] [Green Version]
- Kampouri, M.; Kolokoussis, P.; Argialas, D.; Karathanassi, V. Mapping of forest tree distribution and estimation of forest biodiversity using Sentinel-2 imagery in the University Research Forest Taxiarchis in Chalkidiki, Greece. Geocarto Int. 2019, 34, 1273–1285. [Google Scholar] [CrossRef]
- Ma, X.; Mahecha, M.D.; Migliavacca, M.; van der Plas, F.; Benavides, R.; Ratcliffe, S.; Kattge, J.; Richter, R.; Musavi, T.; Baeten, L.; et al. Inferring plant functional diversity from space: The potential of Sentinel-2. Remote Sens. Environ. 2019, 233, 111368. [Google Scholar] [CrossRef]
- Clark, M.L. Comparison of multi-seasonal Landsat 8, Sentinel-2 and hyperspectral images for mapping forest alliances in Northern California. ISPRS J. Photogramm. Remote Sens. 2020, 159, 26–40. [Google Scholar] [CrossRef]
- Xiao, C.; Li, P.; Feng, Z.; Liu, Y.; Zhang, X. Sentinel-2 red-edge spectral indices (RESI) suitability for mapping rubber boom in Luang Namtha Province, northern Lao PDR. Int. J. Appl. Earth Obs. Geoinf. 2020, 93, 102176. [Google Scholar] [CrossRef]
- Agrillo, E.; Filipponi, F.; Pezzarossa, A.; Casella, L.; Smiraglia, D.; Orasi, A.; Attorre, F.; Taramelli, A. Earth observation and biodiversity big data for forest habitat types classification and mapping. Remote Sens. 2021, 13, 1231. [Google Scholar] [CrossRef]
- Hakkenberg, C.R.; Peet, R.K.; Urban, D.L.; Song, C. Modeling plant composition as community continua in a forest landscape with LiDAR and hyperspectral remote sensing. Ecol. Appl. 2018, 28, 177–190. [Google Scholar] [CrossRef]
- Hościło, A.; Lewandowska, A. Mapping Forest Type and Tree Species on a Regional Scale Using Multi-Temporal Sentinel-2 Data. Remote Sens. 2019, 11, 929. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gong, W.; Hu, X.; Gong, J. Forest Type Identification with Random Forest Using Sentinel-1A, Sentinel-2A, Multi-Temporal Landsat-8 and DEM Data. Remote Sens. 2018, 10, 946. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Skidmore, A.K.; Wang, T.; Holzwarth, S.; Heiden, U.; Pinnel, N.; Zhu, X.; Heurich, M. Tree species classification using plant functional traits from LiDAR and hyperspectral data. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 207–219. [Google Scholar] [CrossRef]
- Waśniewski, A.; Hościło, A.; Zagajewski, B.; Moukétou-Tarazewicz, D. Assessment of Sentinel-2 Satellite Images and Random Forest Classifier for Rainforest Mapping in Gabon. Forests 2020, 11, 941. [Google Scholar] [CrossRef]
- Clark, M.L.; Buck-Diaz, J.; Evens, J. Mapping of forest alliances with simulated multi-seasonal hyperspectral satellite imagery. Remote Sens. Environ. 2018, 210, 490–507. [Google Scholar] [CrossRef]
- Burai, P.; Deák, B.; Valkó, O.; Tomor, T. Classification of Herbaceous Vegetation Using Airborne Hyperspectral Imagery. Remote Sens. 2015, 7, 2046–2066. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.C. Genus-Physiognomy-Ecosystem (GPE) System for Satellite-Based Classification of Plant Communities. Ecologies 2021, 2, 203–213. [Google Scholar] [CrossRef]
- Macintyre, P.; van Niekerk, A.; Mucina, L. Efficacy of multi-season Sentinel-2 imagery for compositional vegetation classification. Int. J. Appl. Earth Obs. Geoinf. 2020, 85, 101980. [Google Scholar] [CrossRef]
- Mishra, A.P.; Rai, I.D.; Pangtey, D.; Padalia, H. Vegetation Characterization at Community Level Using Sentinel-2 Satellite Data and Random Forest Classifier in Western Himalayan Foothills, Uttarakhand. J. Indian Soc. Remote Sens. 2021, 49, 759–771. [Google Scholar] [CrossRef]
- Kupková, L.; Červená, L.; Suchá, R.; Jakešová, L.; Zagajewski, B.; Březina, S.; Albrechtová, J. Classification of tundra vegetation in the Krkonoše Mts. National park using APEX, AISA dual and sentinel-2A data. Eur. J. Remote Sens. 2017, 50, 29–46. [Google Scholar] [CrossRef]
- Suchá, R.; Jakešová, L.; Kupková, L.; Červená, L. Classification of vegetation above the tree line in the Krkonoše Mts. National Park using remote sensing multispectral data. AUC Geogr. 2016, 51, 113–129. [Google Scholar] [CrossRef] [Green Version]
- Dubeau, P.; King, D.J.; Unbushe, D.G.; Rebelo, L.-M. Mapping the Dabus Wetlands, Ethiopia, Using Random Forest Classification of Landsat, PALSAR and Topographic Data. Remote Sens. 2017, 9, 1056. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Mao, D.; Wang, Z.; Qiu, Z.; Yan, H.; Feng, K.; Zhang, Z. Mapping Wetland Plant Communities Using Unmanned Aerial Vehicle Hyperspectral Imagery by Comparing Object/Pixel-Based Classifications Combining Multiple Machine-Learning Algorithms. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 8249–8258. [Google Scholar] [CrossRef]
- Prakash Hati, J.; Samanta, S.; Rani Chaube, N.; Misra, A.; Giri, S.; Pramanick, N.; Gupta, K.; Datta Majumdar, S.; Chanda, A.; Mukhopadhyay, A.; et al. Mangrove classification using airborne hyperspectral AVIRIS-NG and comparing with other spaceborne hyperspectral and multispectral data. Egypt. J. Remote Sens. Space Sci. 2021, 24, 273–281. [Google Scholar] [CrossRef]
- Bradter, U.; O’Connell, J.; Kunin, W.E.; Boffey, C.W.H.; Ellis, R.J.; Benton, T.G. Classifying grass-dominated habitats from remotely sensed data: The influence of spectral resolution, acquisition time and the vegetation classification system on accuracy and thematic resolution. Sci. Total Environ. 2020, 711, 134584. [Google Scholar] [CrossRef]
- Holzman, M.E.; Rivas, R.E.; Bayala, M.I. Relationship between TIR and NIR-SWIR as Indicator of Vegetation Water Availability. Remote Sens. 2021, 13, 3371. [Google Scholar] [CrossRef]









| Sensor | HySpex VNIR-1800 | HySpex SWIR-384 |
|---|---|---|
| Spectral range | 416–995 nm | 954–2510 nm |
| Spatial pixels | 1800 | 384 |
| Number of spectral bands | 163 | 288 |
| Spatial resolution | 1.0 m | 2.0 m |
| Field of view (FOV) across the track | 17–34° | 16–32° |
| Instantaneous field of view (IFOV) | 0.01–0.04° | 0.04–0.08° |
| Full width at half maximum (FWHM) | 3.26 nm | 5.45 nm |
| Date | Sentinel-2 Satellite | Track | Cloud Coverage (%) |
|---|---|---|---|
| 15 September 2019 | 2A | 79 | 0.33% |
| 22 September 2019 | 2A | 36 | 0.06% |
| 17 October 2019 | 2B | 36 | 0.27% |
| 25 October 2019 | 2A | 79 | 0.01% |
| 22 August 2020 | 2B | 36 | 0.24% |
| 4 September 2020 | 2B | 79 | 1.10% |
| 9 September 2020 | 2A | 79 | 0.03% |
| 14 September 2020 | 2B | 79 | 0.72% |
| 21 September 2020 | 2B | 36 | 1.11% |
| 9 September 2021 | 2B | 79 | 0.05% |
| 9 October 2021 | 2B | 79 | 0.04% |
| Acronym | Class | Description |
|---|---|---|
| RS | Rocks and scree communities | Vegetation growing on a loose bedrock or bare rock (initial phases of cryptogamic plant communities, epilithic lichens, and scree communities) |
| LA | Luzuletum alpino-pilosae | Alpine grasslands |
| OD | Oreochloo distichae-Juncetum trifidi | Alpine grasslands |
| FV | Festuco versicoloris Agrostietum | Alpine grasslands |
| CA | Calamagrostietum | Alpine grasslands |
| DF | Deschampsia flexuosa community | Alpine grasslands |
| ON | Other non-forest vegetation | Vegetation of mountain pastures and communities in transition |
| LO | Low shrubs | Vaccinium myrtillus, Empetrum nigrum and Calluna vulgaris |
| PM | Pinetum mugo carpaticum | Subalpine dwarf pine shrubs |
| MS | Montane spruce forests | Coniferous forests composed of Picea abies and an admixture of Abies alba |
| SN | Snags | Damaged Norway spruce (Picea abies) |
| DE | Deciduous forest | Fagus sylvatica, Acer pseudoplatanus |
| WA | Water | Stream and mountain lake waters |
| Class | Total Polygons | Total Pixels | |
|---|---|---|---|
| HySpex/MNF | Sentinel-2 | ||
| Rocks and scree communities | 18 | 579 | 36 |
| Luzuletum alpino-pilosae | 19 | 844 | 44 |
| Oreochloo distichae-Juncetum trifidi | 59 | 4465 | 161 |
| Festuco versicoloris-Agrostietum | 4 | 235 | 11 |
| Calamagrostietum | 13 | 1004 | 34 |
| Deschampsia flexuosa community | 15 | 233 | 37 |
| Other non-forest vegetation | 16 | 10,285 | 373 |
| Low shrubs | 58 | 4529 | 192 |
| Pinetum mugo carpaticum | 52 | 15,627 | 561 |
| Montane spruce forests | 66 | 33,005 | 1200 |
| Snags | 38 | 7750 | 263 |
| Deciduous forest | 25 | 9757 | 339 |
| Water | 6 | 5587 | 206 |
| Total | 389 | 93,900 | 3457 |
| Spectral | Spectral + DTM | Spectral + nDSM | Spectral + Slope | Spectral + Aspect | Spectral + TF | ||
|---|---|---|---|---|---|---|---|
| RF | HySpex | 75.3 | 85.7 | 79.5 | 78.1 | 76.2 | 89.6 |
| MNF | 75.6 | 83.0 | 77.2 | 77.8 | 77.9 | 88.8 | |
| S-2 | 83.5 | 87.4 | 85.3 | 84.7 | 83.6 | 88.9 | |
| SVM | HySpex | 78.5 | 86.2 | 80.6 | 81.4 | 79.0 | 88.6 |
| MNF | 69.7 | 78.2 | 72.6 | 72.2 | 70.4 | 88.8 | |
| S-2 | 86.0 | 87.1 | 86.8 | 86.6 | 86.4 | 88.5 |
| Class | HySpex | Sentinel-2 | MNF |
|---|---|---|---|
| Rocks and scree communities | +0.9 (Slope) | +2.1 (DTM) | +1.2 (nDSM) |
| Luzuletum alpino-pilosae | +8.4 (Aspect) | +0.6 (Slope) | +19.3 (Aspect) |
| Oreochloo distichae-Juncetum trifidi | +16.9 (Aspect) | +4.9 (DTM) | +14.6 (DTM) |
| Festuco versicoloris-Agrostietum | +13.3 (DTM) | +6.8 (Slope) | +2.4 (Slope) |
| Calamagrostietum | +28.5 (DTM) | +3.9 (nDSM) | +28.6 (DTM) |
| Deschampsia flexuosa community | +33.2 (DTM) | +6.5 (DTM) | +26.9 (DTM) |
| Other non-forest vegetation | +6.1 (nDSM) | +6.8 (DTM) | +4.3 (Slope) |
| Low shrubs | +15.8 (DTM) | +6.3 (DTM) | +10.4 (DTM) |
| Pinetum mugo carpaticum | +9.7 (nDSM) | +9.7 (nDSM) | +6.9 (nDSM) |
| Montane spruce forests | +8.3 (DTM) | +5.3 (nDSM) | +5.7 (nDSM) |
| Snags | +2.4 (DTM) | +0.4 (nDSM) | +0.1 (Slope) |
| Deciduous forest | +8.9 (DTM) | +3.0 (DTM) | +6.6 (DTM) |
| Water | +0.4 (nDSM) | +1.0 (nDSM) | +7.4 (Slope) |
| CA | DE | DF | LO | FV | LA | OD | ON | PM | MS | RS | SN | WA | Σ | UA | F1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CA | 217 | 0 | 0 | 2 | 0 | 0 | 29 | 0 | 0 | 2 | 10 | 0 | 0 | 260 | 83.5 | 87.7 |
| DE | 0 | 6059 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 566 | 0 | 16 | 0 | 6642 | 91.2 | 91.9 |
| DF | 0 | 0 | 99 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 103 | 96.1 | 91.2 |
| LO | 16 | 0 | 3 | 1713 | 0 | 0 | 56 | 0 | 9 | 0 | 0 | 2 | 0 | 1799 | 95.2 | 96.5 |
| FV | 0 | 0 | 0 | 0 | 39 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 39 | 100 | 100 |
| LA | 0 | 0 | 1 | 2 | 0 | 505 | 58 | 0 | 0 | 0 | 0 | 0 | 0 | 566 | 89.2 | 88.0 |
| OD | 2 | 0 | 10 | 31 | 0 | 77 | 2178 | 0 | 0 | 0 | 0 | 1 | 0 | 2299 | 94.7 | 94.0 |
| ON | 0 | 9 | 1 | 0 | 0 | 0 | 4 | 6034 | 0 | 1 | 0 | 0 | 0 | 6049 | 99.8 | 99.9 |
| PM | 0 | 1 | 0 | 1 | 0 | 0 | 6 | 0 | 6942 | 32 | 0 | 0 | 0 | 6982 | 99.4 | 99.6 |
| MS | 0 | 410 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 19,287 | 0 | 196 | 0 | 19,905 | 96.9 | 96.6 |
| RS | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 470 | 10 | 0 | 485 | 96.9 | 97.1 |
| SN | 0 | 65 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 128 | 3 | 3005 | 0 | 3201 | 93.9 | 93.5 |
| WA | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1232 | 1232 | 100 | 100 |
| Σ | 235 | 6544 | 114 | 1753 | 39 | 582 | 2336 | 6034 | 6964 | 20,016 | 483 | 3230 | 1232 | |||
| PA | 92.3 | 92.6 | 86.8 | 97.7 | 100 | 86.8 | 93.2 | 100 | 99.7 | 96.4 | 97.3 | 93.0 | 100 |
| Author | Data Used | No. of Classes | Object of Classification | Classifier | OA(%) |
|---|---|---|---|---|---|
| Our results | HySpex HySpex + TF MNF MNF_HySpex + TF Sentinel-2 Sentinel-2 + TF | 13 | Mountain forest and non-forest plant communities | RF SVM RF SVM RF SVM RF SVM RF SVM RF SVM | 83.4 87.9 96.4 94.5 85.1 79.5 95.7 90.4 87.9 92.3 98.5 95.3 |
| [96] | AVIRIS multitemporal | 16 | Forest alliances | SVM | 75.9 |
| [27] | MNF_APEX | 23 | Mountain non-forest plant communities | SVM | 74.3 74.4 |
| [97] | AISA EAGLE II | 19 | Herbaceous vegetation | RF SVM | 79 82 |
| [88] | Sentinel-2 Landsat 8 | 12 | Forest alliances | SVM | 83.7 78.6 |
| [50] | Multitemporal Sentinel-2 | 9 | Mountain non-forest plant communities | SVM | 74.2 |
| [98] | Landsat 8 | 6 | Vegetation types | RF | 83.4 |
| [99] | Multitemporal Sentinel-2 | 24 | Vegetationclassification | RF SVM | 71 78 |
| [2] | Multitemporal GF-2, ZY-3, GF-1 | 11 | Mountain forest and non-forest vegetation | RF | 92.2 |
| [93] | S-1 (VV) + S-2 + L8 + DEM | 9 | Forest types | RF | 82.8 |
| [100] | Multitemporal Sentinel-2 | 17 | Mountain vegetation communities | RF | 87 |
| [11] | Sentinel-2 Landsat 8 OLI | 4 | Mountain forest | SVM RF ANN SVM RF ANN | 87 83 84 83 85 77 |
| [101] | APEX Sentinel-2 | 7 | Mountain vegetation communities | SVM | 84.3 77.7 |
| [102] | WV-2 Landsat 8 OLI | 8 | Mountain vegetation communities | MLC SVM | 68.4 78.31 |
| [49] | Sentinel-2 | 9 | Forest types | RF | 92.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kluczek, M.; Zagajewski, B.; Kycko, M. Airborne HySpex Hyperspectral Versus Multitemporal Sentinel-2 Images for Mountain Plant Communities Mapping. Remote Sens. 2022, 14, 1209. https://doi.org/10.3390/rs14051209
Kluczek M, Zagajewski B, Kycko M. Airborne HySpex Hyperspectral Versus Multitemporal Sentinel-2 Images for Mountain Plant Communities Mapping. Remote Sensing. 2022; 14(5):1209. https://doi.org/10.3390/rs14051209
Chicago/Turabian StyleKluczek, Marcin, Bogdan Zagajewski, and Marlena Kycko. 2022. "Airborne HySpex Hyperspectral Versus Multitemporal Sentinel-2 Images for Mountain Plant Communities Mapping" Remote Sensing 14, no. 5: 1209. https://doi.org/10.3390/rs14051209