
Combining Passive Acoustics and Environmental Data for Scaling Up Ecosystem Monitoring: A Test on Coral Reef Fishes

 , , and
, , and
Abstract
:1. Introduction
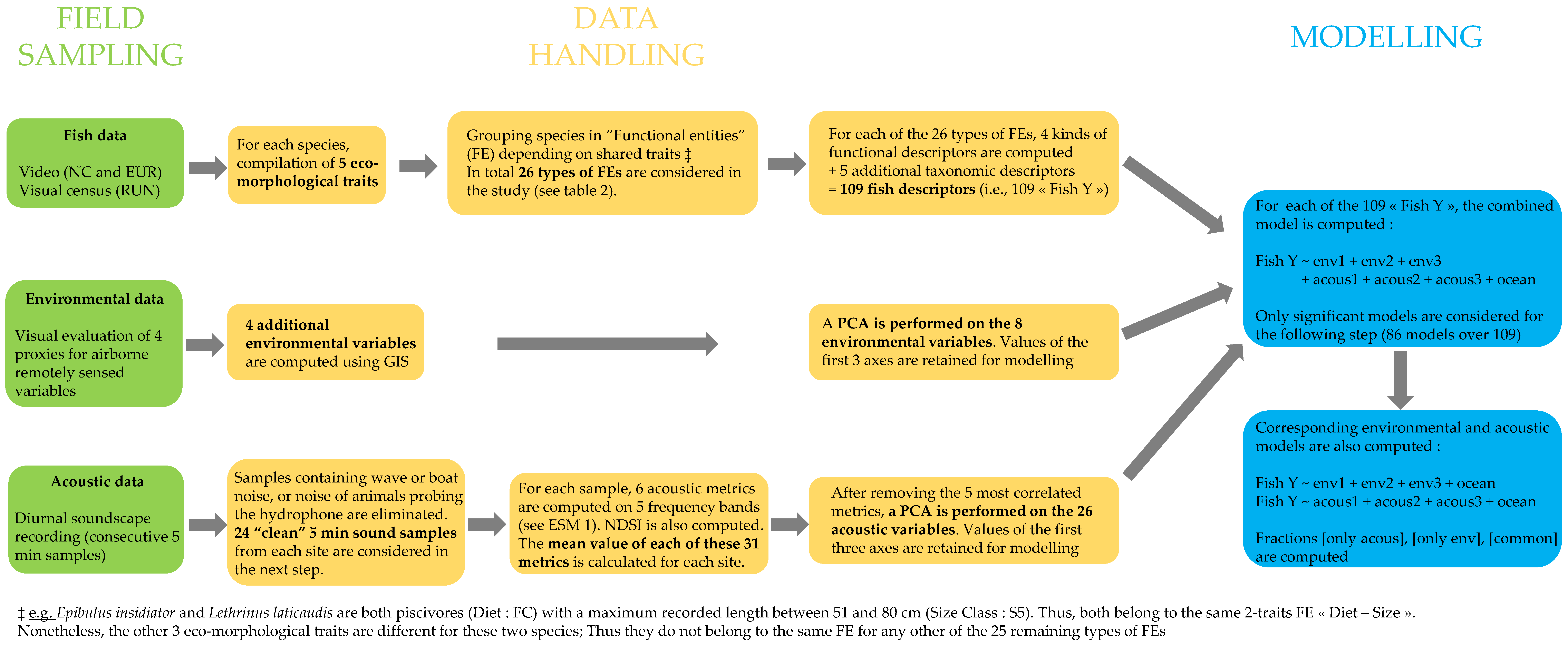
2. Materials and Methods
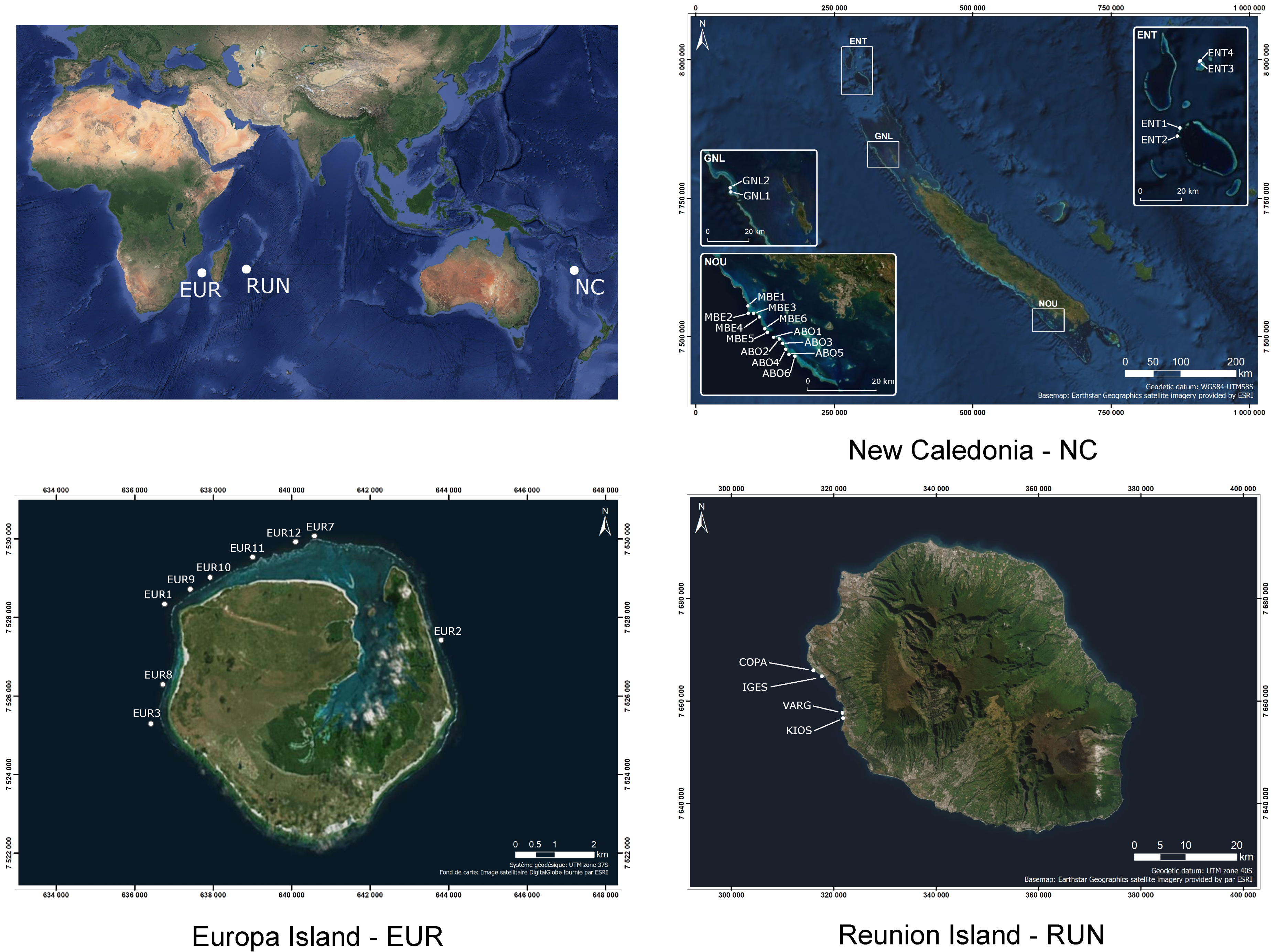
2.1. Study Sites
2.2. Evaluation of Reef Fish Assemblage Characteristics
2.3. Environmental Variables
2.4. Soundscape Recordings and Acoustic Metrics
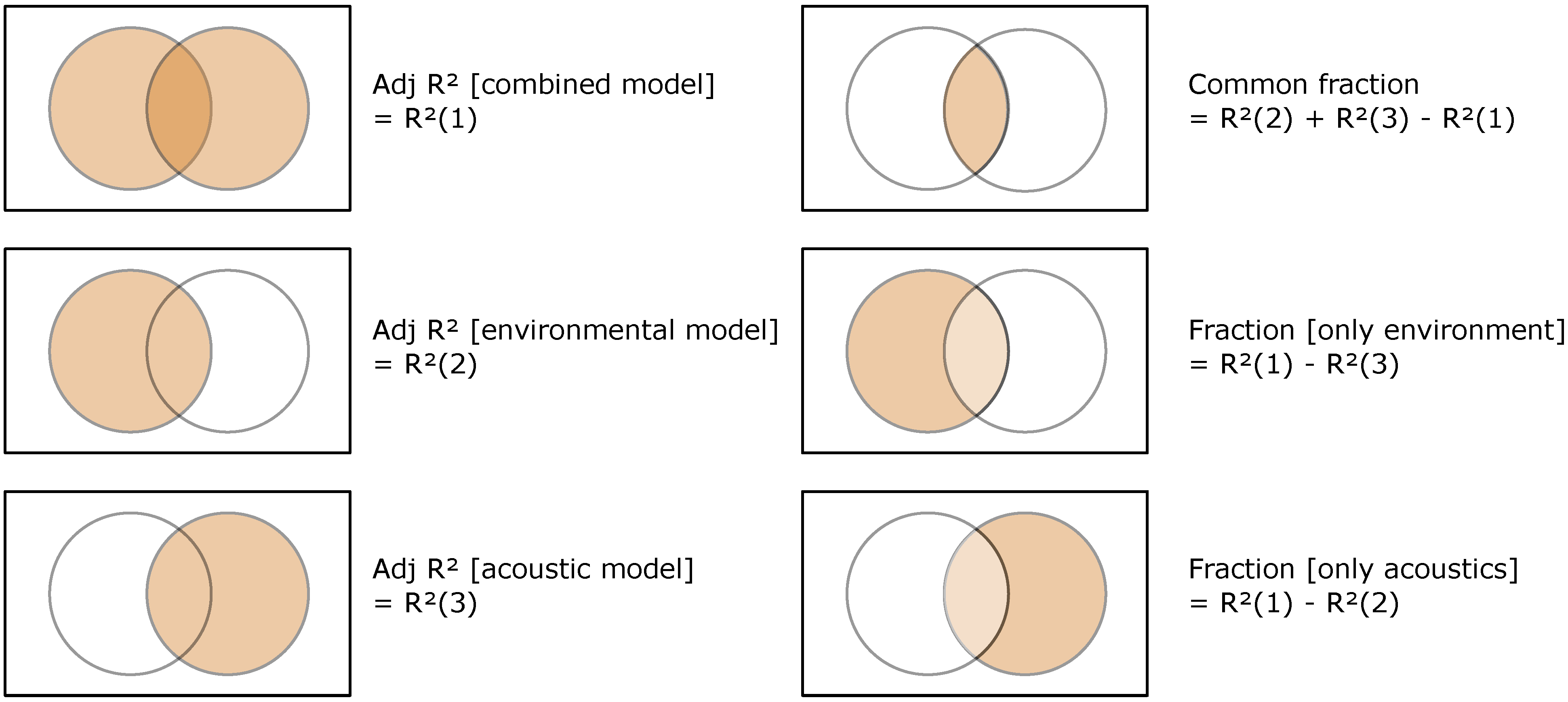
2.5. Statistical Analyses
2.5.1. Reduction of Environmental Data and Acoustic Data
2.5.2. Test of the Added Value of Acoustic data in Predicting Fish Assemblage Characteristics
2.5.3. Identification of the Fish Assemblage Characteristics Best Predicted by Acoustic Data
3. Results
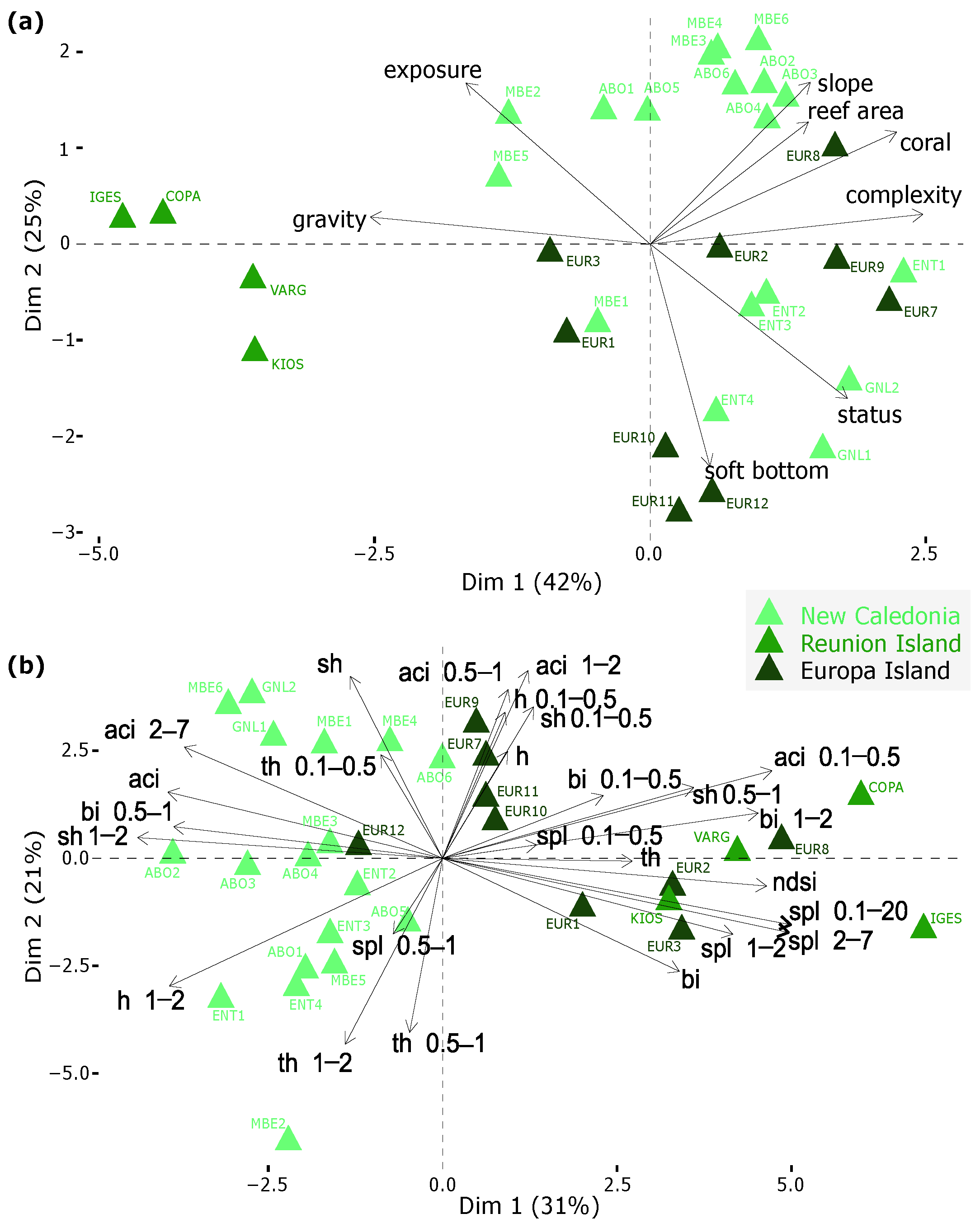
3.1. Reduction of Environmental Data and Acoustic Data
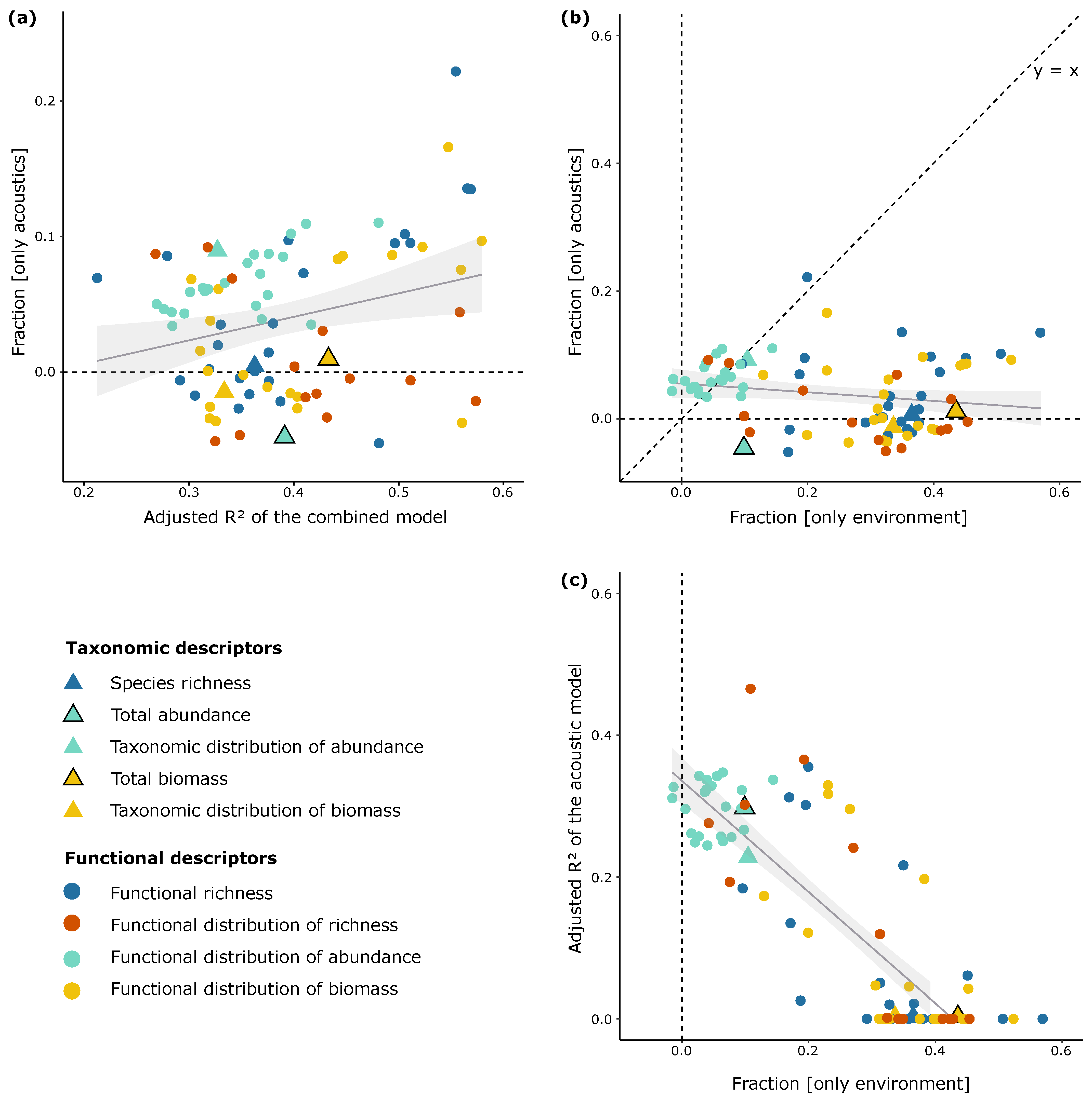
3.2. Test of the Added Value of Acoustic Data in Predicting Fish Assemblage Characteristics
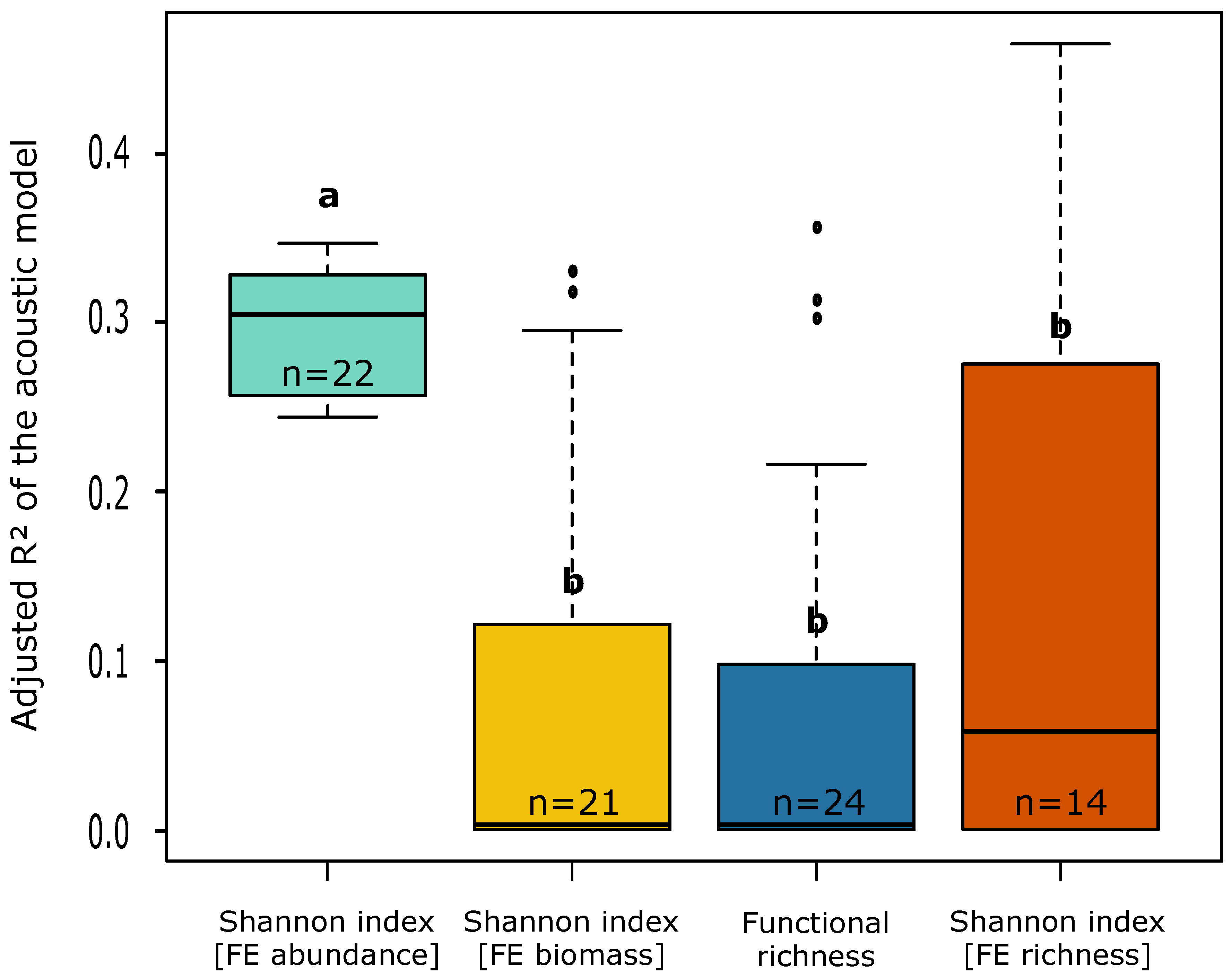
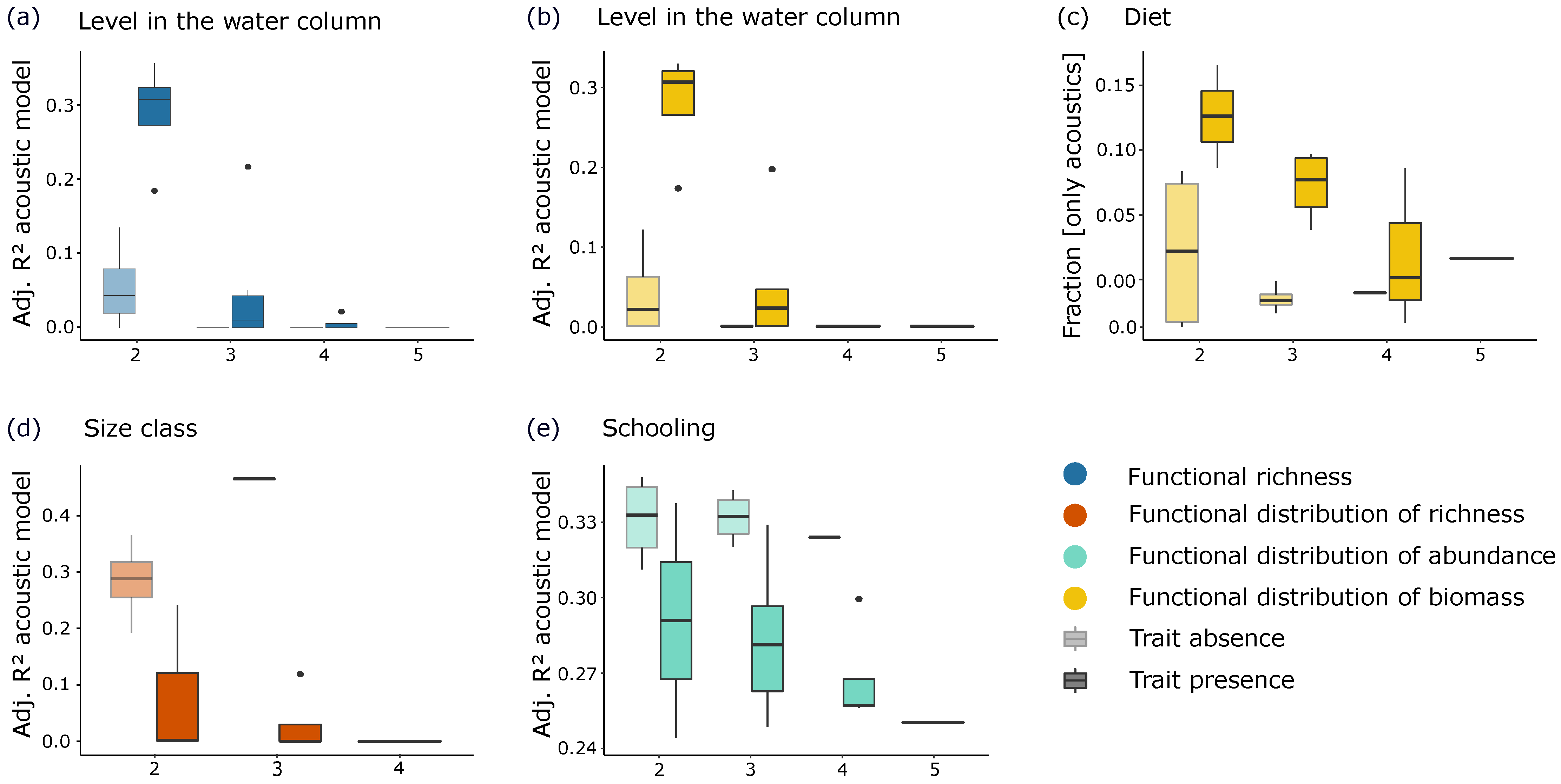
3.3. Identification of the Fish Assemblage Characteristics Best Predicted by the Acoustic Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harborne, A.; Rogers, A.; Bozec, Y.; Mumby, P.J. Multiple Stressors and the Functioning of Coral Reefs. Annu. Rev. Mar. Sci. 2016, 9, 445–468. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.; Kleypas, J.; Van De Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Kennedy, E.V.; Beyer, H.L.; McClennen, C.; Possingham, H.P. Securing a long-term future for coral reefs. Trends Ecol. Evol. 2018, 12, 936–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.J.; Graham, N.A.J. Rethinking coral reef functional futures. Funct. Ecol. 2019, 33, 942–947. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Pratchett, M.S.; Morrison, T.H.; Gurney, G.G.; Hughes, T.P.; Álvarez-Romero, J.G.; Day, J.C.; Grantham, R.; Grech, A.; Hoey, A.S.; et al. Coral reef conservation in the Anthropocene: Confronting spatial mismatches and prioritizing functions. Biol. Conserv. 2019, 236, 604–615. [Google Scholar] [CrossRef]
- Duarte, C.M.; Agusti, S.; Barbier, E.; Britten, G.L.; Castilla, J.C.; Gattuso, J.P.; Fulweiler, R.W.; Hughes, T.P.; Knowlton, N.; Lovelock, C.E.; et al. Rebuilding marine life. Nature 2020, 580, 39–51. [Google Scholar] [CrossRef]
- Obura, D.O.; Aeby, G.; Amornthammarong, N.; Appeltans, W.; Bax, N.J.; Bishop, J.; Brainard, R.E.; Chan, S.; Fletcher, P.; Gordon, T.A.; et al. Coral reef monitoring, reef assessment technologies, and ecosystem-based management. Front. Mar. Sci. 2019, 6, 580. [Google Scholar] [CrossRef] [Green Version]
- Roelfsema, C.M.; Kovacs, E.; Ortiz, J.C.; Wolff, N.H.; Callaghan, D.; Wettle, M.; Ronan, M.; Hamylton, S.M.; Mumby, P.J.; Phinn, S. Coral reef habitat mapping: A combination of object-based image analysis and ecological modelling. Remote Sens. Environ. 2018, 208, 27–41. [Google Scholar] [CrossRef]
- Roelfsema, C.M.; Kovacs, E.M.; Ortiz, J.C.; Callaghan, D.P.; Hock, K.; Mongin, M.; Johansen, K.; Mumby, P.J.; Wettle, M.; Ronan, M.; et al. Habitat maps to enhance monitoring and management of the Great Barrier Reef. Coral Reefs 2020, 39, 1039–1054. [Google Scholar] [CrossRef]
- Bajjouk, T.; Mouquet, P.; Ropert, M.; Quod, J.P.; Hoarau, L.; Bigot, L.; Le Dantec, N.; Delacourt, C.; Populus, J. Detection of changes in shallow coral reefs status: Towards a spatial approach using hyperspectral and multispectral data. Ecol. Indic. 2019, 96, 174–191. [Google Scholar] [CrossRef]
- González-Barrios, F.J.; Álvarez-Filip, L. A framework for measuring coral species-specific contribution to reef functioning in the Caribbean. Ecol. Indic. 2018, 95, 877–886. [Google Scholar] [CrossRef]
- Obura, D.O.; Gudka, M.; Rabi, F.A.; Gian, S.B.; Bijoux, J.; Freed, S.; Maharavo, J.; Mwaura, J.; Porter, S.; Sola, E.; et al. Coral Reef Status Report for the Western Indian Ocean (2017). In Nairobi Convention; Global Coral Reef Monitoring Network (GCRMN)/International Coral Reef Initiative (ICRI); Indian Ocean Commission: Port Louis, Mauritius, 2017. [Google Scholar]
- Mellin, C.; Parrott, L.; Andréfouët, S.; Bradshaw, C.J.A.; MacNeil, M.A.; Caley, M.J. Multi-scale marine biodiversity patterns inferred efficiently from habitat image processing. Ecol. Appl. 2012, 22, 792–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staaterman, E.; Ogburn, M.B.; Altieri, A.H.; Brandl, S.J.; Seemann, J.; Whippo, R.; Goodison, M.; Duffy, J.E. Bioacoustic measurements complement visual biodiversity surveys: Preliminary evidence from four shallow marine habitats. Mar. Ecol. Prog. Ser. 2017, 575, 207–215. [Google Scholar] [CrossRef] [Green Version]
- West, K.M.; Stat, M.; Harvey, E.S.; Skepper, C.L.; DiBattista, J.D.; Richards, Z.T.; Travers, M.J.; Newman, S.J.; Bunce, M. eDNA metabarcoding survey reveals fine-scale coral reef community variation across a remote, tropical island ecosystem. Mol. Ecol. 2020, 29, 1069–1086. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, W.; Getinet, W. Remote sensing for marine spatial planning and integrated coastal areas management: Achievements, challenges, opportunities and future prospects. Remote Sens. Appl. Soc. Environ. 2016, 4, 138–157. [Google Scholar] [CrossRef]
- Wedding, L.M.; Jorgensen, S.; Lepczyk, C.A.; Friedlander, A.M. Remote sensing of three-dimensional coral reef structure enhances predictive modeling of fish assemblages. Remote Sens. Ecol. Conserv. 2019, 5, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Rappaport, D.I.; Royle, J.A.; Morton, D.C. Acoustic space occupancy: Combining ecoacoustics and lidar to model biodiversity variation and detection bias across heterogeneous landscapes. Ecol. Indic. 2020, 113, 106172. [Google Scholar] [CrossRef]
- Cinner, J.E.; Maire, E.; Huchery, C.; MacNeil, M.A.; Graham, N.A.J.; Mora, C.; McClanahan, T.R.; Barnes, M.L.; Kittinger, J.N.; Hicks, C.C.; et al. Gravity of human impacts mediates coral reef conservation gains. Proc. Natl. Acad. Sci. USA 2018, 115, E6116–E6125. [Google Scholar] [CrossRef] [Green Version]
- Purkis, S.J.; Gleason, A.C.R.; Purkis, C.R.; Dempsey, A.C.; Renaud, P.G.; Faisal, M.; Saul, S.; Kerr, J.M. High-resolution habitat and bathymetry maps for 65,000 sq. km of Earth’s remotest coral reefs. Coral Reefs 2019, 38, 467–488. [Google Scholar] [CrossRef] [Green Version]
- Friedlander, A.M.; Brown, E.K.; Jokiel, P.L.; Smith, W.R.; Rodgers, K.S. Effects of habitat, wave exposure, and marine protected area status on coral reef fish assemblages in the Hawaiian archipelago. Coral Reefs 2003, 22, 291–305. [Google Scholar] [CrossRef]
- Darling, E.S.; Graham, N.A.J.; Januchowski-Hartley, F.A.; Nash, K.L.; Pratchett, M.S.; Wilson, S.K. Relationships between structural complexity, coral traits, and reef fish assemblages. Coral Reefs 2017, 36, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Barneche, D.R.; Rezende, E.L.; Parravicini, V.; Maire, E.; Edgar, G.J.; Stuart-Smith, R.D.; Arias-González, J.E.; Ferreira, C.E.; Friedlander, A.M.; Green, A.L.; et al. Body size, reef area and temperature predict global reef-fish species richness across spatial scales. Glob. Ecol. Biogeogr. 2019, 28, 315–327. [Google Scholar] [CrossRef]
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Rogers, A.; Blanchard, J.L.; Mumby, P.J. Fisheries productivity under progressive coral reef degradation. J. Appl. Ecol. 2018, 55, 1041–1049. [Google Scholar] [CrossRef]
- Pieretti, N.; Martire, M.L.; Farina, A.; Danovaro, R. Marine soundscape as an additional biodiversity monitoring tool: A case study from the Adriatic Sea (Mediterranean Sea). Ecol. Indic. 2017, 83, 13–20. [Google Scholar] [CrossRef]
- Carrasco, S.A.; Bravo, M.; Avilés, E.; Ruíz, P.; Yori, A.; Hinojosa, I.A. Exploring overlooked components of remote South-east Pacific oceanic islands: Larval and macrobenthic assemblages in reef habitats with distinct underwater soundscapes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 273–289. [Google Scholar] [CrossRef]
- Ferrier-Pagès, C.; Leal, M.C.; Calado, R.; Schmid, D.W.; Bertucci, F.; Lecchini, D.; Allemand, D. Noise pollution on coral reefs?—A yet underestimated threat to coral reef communities. Mar. Pollut. Bull. 2021, 165, 112129. [Google Scholar] [CrossRef]
- Parsons, M.; Lin, T.H.; Mooney, A.; Erbe, C.; Juanes, F.; Lammers, M.; Li, S.; Linke, S.; Looby, A.; Nedelec, S.L.; et al. Sounding the Call for a Global Library of Underwater Biological Sounds. Front. Ecol. Evol. 2022, 10, 810156. [Google Scholar] [CrossRef]
- Mooney, T.A.; Di Iorio, L.; Lammers, M.; Lin, T.H.; Nedelec, S.L.; Parsons, M.; Radford, C.; Urban, E.; Stanley, J. Listening forward: Approaching marine biodiversity assessments using acoustic methods. R. Soc. Open Sci. 2020, 7, 201287. [Google Scholar] [CrossRef]
- Kennedy, E.V.; Holderied, M.W.; Mair, J.M.; Guzman, H.M.; Simpson, S.D. Spatial patterns in reef-generated noise relate to habitats and communities: Evidence from a Panamanian case study. J. Exp. Mar. Bio. Ecol. 2010, 395, 85–92. [Google Scholar] [CrossRef]
- Bertucci, F.; Parmentier, E.; Lecellier, G.; Hawkins, A.D.; Lecchini, D. Acoustic indices provide information on the status of coral reefs: An example from Moorea Island in the South Pacific. Sci. Rep. 2016, 6, 33326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elise, S.; Urbina-Barreto, I.; Pinel, R.; Mahamadaly, V.; Bureau, S.; Penin, L.; Adjeroud, M.; Kulbicki, M.; Bruggemann, J.H. Assessing key ecosystem functions through soundscapes: A new perspective from coral reefs. Ecol. Indic. 2019, 107, 105623. [Google Scholar] [CrossRef]
- Myers, E.; Harvey, E.; Saunders, B.; Travers, M. Fine-scale patterns in the day, night and crepuscular composition of a temperate reef fish assemblage. Mar. Ecol. 2016, 37, 668–678. [Google Scholar] [CrossRef]
- FishBase. 2019. Available online: http://www.fishbase.org/ (accessed on 20 December 2019).
- Labrosse, P.; Kulbicki, M.; Ferraris, J. Underwater Visual Fish Census Surveys; Secretariat of the Pacific Community: Nouméa, Nouvelle-Calédonie, 2002; 60p. [Google Scholar]
- Mellin, C.; Mouillot, D.; Kulbicki, M.; McClanahan, T.R.; Vigliola, L.; Bradshaw, C.J.A.; Brainard, R.E.; Chabanet, P.; Edgar, G.J.; Fordham, D.A.; et al. Humans and seasonal climate variability threaten large bodied coral reef fish with small ranges. Nat. Commun. 2016, 7, 10491. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R.; Streit, R.P.; Brandl, S.J.; Tebbett, S.B. The meaning of the term “function” in ecology: A coral reef perspective. Funct. Ecol. 2019, 33, 948–961. [Google Scholar] [CrossRef] [Green Version]
- UNEP-WCMC; WorldFish Centre; WRI; TNC. Global Distribution of Warm-Water Coral Reefs, Compiled from Multiple Sources Including the Millennium Coral Reef Mapping Project; Version 1.3; Includes Contributions from IMaRS-USF and IRD (2005), IMaRS-USF (2005) and Spalding et al. (2001); UNEP World Conservation Monitoring Centre: Cambridge, UK, 2010; Available online: http://data.unep-wcmc.org/datasets/1 (accessed on 15 January 2020).
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2018. Available online: http://qgis.osgeo.org (accessed on 15 January 2020).
- WDPA. 2020. Available online: https://www.protectedplanet.net/en (accessed on 15 January 2020).
- Polunin, N.V.C.; Roberts, C.M. Greater biomass and value of target coral-reef fishes in two small Caribbean marine reserves. Mar. Ecol. Prog. Ser. 1993, 100, 167. [Google Scholar] [CrossRef]
- English, S.; Wilkinson, C.; Baker, V. Survey Manual for Tropical Marine Resources, 2nd ed.; AIMS: Townsville, Australia, 1997. [Google Scholar]
- Elise, S.; Bailly, A.; Urbina-Barreto, I.; Mou-Tham, G.; Chiroleu, F.; Vigliola, L.; Robbins, W.D.; Bruggemann, J.H. An optimised passive acoustic sampling scheme to discriminate among coral reefs’ ecological states. Ecol. Indic. 2019, 107, 105627. [Google Scholar] [CrossRef]
- Legendre, P. Studying Beta Diversity: Ecological Variation Partitioning by Multiple Regression and Canonical Analysis. Chin. J. Plant Ecol. 2007, 31, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Heenan, A.; Williams, G.J.; Williams, I.D. Natural variation in coral reef trophic structure across environmental gradients. Front. Ecol. Environ. 2020, 18, 69–75. [Google Scholar] [CrossRef]
- Beyer, H.L.; Kennedy, E.V.; Beger, M.; Chen, C.A.; Cinner, J.E.; Darling, E.S.; Eakin, C.M.; Gates, R.D.; Heron, S.F.; Knowlton, N.; et al. Risk-sensitive planning for conserving coral reefs under rapid climate change. Conserv. Lett. 2018, 11, e12587. [Google Scholar] [CrossRef]
- Mumby, P.J. Trends and frontiers for the science and management of the oceans. Curr. Biol. 2017, 27, R431–R434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madin, E.M.; Darling, E.S.; Hardt, M.J. Emerging technologies and coral reef conservation: Opportunities, challenges, and moving forward. Front. Mar. Sci. 2019, 6, 727. [Google Scholar] [CrossRef] [Green Version]
- Chérubin, L.M.; Dalgleish, F.; Ibrahim, A.K.; Schärer-Umpierre, M.; Nemeth, R.S.; Matthews, A.; Appeldoorn, R. Fish Spawning Aggregations Dynamics as Inferred From a Novel, Persistent Presence Robotic Approach. Front. Mar. Sci. 2020, 6, 779. [Google Scholar] [CrossRef]
- Hopkinson, B.M.; King, A.C.; Owen, D.P.; Johnson-Roberson, M.; Long, M.H.; Bhandarkar, S.M. Automated classification of three-dimensional reconstructions of coral reefs using convolutional neural networks. PLoS ONE 2020, 15, e0230671. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Nadaoka, K.; Nakamura, T. Towards Benthic Habitat 3D Mapping Using Machine Learning Algorithms and Structures from Motion Photogrammetry. Remote Sens. 2020, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.S.; Jones, N.S.; Fulcher, B.D.; Picinali, L.; Clink, D.J.; Klinck, H.; Orme, C.D.; Wrege, P.H.; Ewers, R.M. Characterizing soundscapes across diverse ecosystems using a universal acoustic feature set. Proc. Natl. Acad. Sci. USA 2020, 117, 17049–17055. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.; Chung, K.W.; Salloum, H.; Sedunov, A.; Sedunov, N.; Sutin, A.; Graber, H.; Mallas, P. Concurrent use of satellite imaging and passive acoustics for maritime domain awareness. In Proceedings of the 2010 International Waterside Security Conference, Carrara, Italy, 3–5 November 2010; IEEE: Manhattan, NY, USA, 2010; pp. 1–8. [Google Scholar]
- Sveegaard, S.; Galatius, A.; Dietz, R.; Kyhn, L.; Koblitz, J.C.; Amundin, M.; Nabe-Nielsen, J.; Sinding, M.H.; Andersen, L.W.; Teilmann, J. Defining management units for cetaceans by combining genetics, morphology, acoustics and satellite tracking. Glob. Ecol. Conserv. 2015, 3, 839–850. [Google Scholar] [CrossRef] [Green Version]
- McLaren, K.; McIntyre, K.; Prospere, K. Using the random forest algorithm to integrate hydroacoustic data with satellite images to improve the mapping of shallow nearshore benthic features in a marine protected area in Jamaica. GIScience Remote Sens. 2019, 56, 1065–1092. [Google Scholar] [CrossRef]
- Lobel, P.S.; Kaatz, I.M.; Rice, A.N. Acoustical Behavior of Coral Reef Fishes, Reproduction and Sexuality in Marine FishesPatterns and Processes; University of California Press: Berkeley, CA, USA, 2010. [Google Scholar] [CrossRef]
- Amorim, M.C.P. Communication in Fishes: Diversity of Sound Production in Fish. Commun. Fishes 2006, 1, 71–105. [Google Scholar]
- Holmes, T.H.; Wilson, S.K.; Travers, M.J.; Langlois, T.J.; Evans, R.D.; Moore, G.I.; Douglas, R.A.; Shedrawi, G.; Harvey, E.S.; Hickey, K. A comparison of visual-and stereo-video based fish community assessment methods in tropical and temperate marine waters of Western Australia. Limnol. Oceanogr. Methods 2013, 11, 337–350. [Google Scholar] [CrossRef]
- Mallet, D.; Wantiez, L.; Lemouellic, S.; Vigliola, L.; Pelletier, D. Complementarity of rotating video and underwater visual census for assessing species richness, frequency and density of reef fish on coral reef slopes. PLoS ONE 2014, 9, e84344. [Google Scholar] [CrossRef] [PubMed]
- Schramm, K.D.; Harvey, E.S.; Goetze, J.S.; Travers, M.J.; Warnock, B.; Saunders, B.J. A comparison of stereo-BRUV, diver operated and remote stereo-video transects for assessing reef fish assemblages. J. Exp. Mar. Biol. Ecol. 2020, 524, 151273. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Holmes, T.H.; MacNeil, M.A.; Ryan, N.M. Visual versus video methods for estimating reef fish biomass. Ecol. Indic. 2018, 85, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Zarco-Perello, S.; Enríquez, S. Remote underwater video reveals higher fish diversity and abundance in seagrass meadows, and habitat differences in trophic interactions. Sci. Rep. 2019, 9, 6596. [Google Scholar] [CrossRef]
- Villanueva-Rivera, L.J.; Pijanowski, B.C. Soundecology: Soundscape Ecology. R Package. 2016. Available online: https://CRAN.R-project.org/package=soundecology (accessed on 4 March 2022).
- Sueur, J. Sound Analysis and Synthesis with R; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar] [CrossRef]
- Boelman, N.T.; Asner, G.P.; Hart, P.J.; Martin, R.E. Multi-trophic invasion resistance in Hawaii: Bioacoustics, field surveys, and airborne remote sensing. Ecol. Appl. 2007, 17, 2137–2144. [Google Scholar] [CrossRef]
- Sueur, J.; Aubin, T.; Simonis, C. Seewave: A free modular tool for sound analysis and synthesis. Bioacoustics 2008, 18, 213–226. [Google Scholar] [CrossRef]
- Sueur, J.; Pavoine, S.; Hamerlynck, O.; Duvail, S. Rapid acoustic survey for biodiversity appraisal. PLoS ONE 2008, 3, e4065. [Google Scholar] [CrossRef] [Green Version]
- Pieretti, N.; Farina, A.; Morri, D. A new methodology to infer the singing activity of an avian community: The Acoustic Complexity Index (ACI). Ecol. Ind. 2011, 11, 868–873. [Google Scholar] [CrossRef]
- Harris, S.A.; Shears, N.T.; Radford, C.A. Ecoacoustic indices as proxies for biodiversity on temperate reefs. Methods Ecol. Evol. 2016, 7, 713–724. [Google Scholar] [CrossRef]
- Ligges, U.; Krey, S.; Mersmann, O.; Schnackenberg, S. tuneR: Analysis of Music and Speech. R Package. 2018. Available online: https://CRAN.R-project.org/package=tuneR (accessed on 4 March 2022).
- Bolgan, M.; Amorim, M.C.P.; Fonseca, P.J.; Di Iorio, L.; Parmentier, E. Acoustic Complexity of vocal fish communities: A field and controlled validation. Sci. Rep. 2018, 8, 10559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnenstiehl, D.R.; Lyon, R.P.; Caretti, O.N.; Ricci, S.W.; Eggleston, D.B. Investigating the utility of ecoacoustic metrics in marine soundscapes. J. Ecoacoustics 2018, 2, R1156L. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Species Size Class | Schooling | Mobility | Level in the Water Column |
|---|---|---|---|---|
| H: herbivores | S1: <7 cm | Sol: solitary species | Sed: sedentary species | Bottom: species staying on the bottom |
| OM: omnivores | S2: 7–15 cm | Pairs: species living in pairs | Mob: species staying within the same reef for several days | Demersal: species hovering just above the bottom |
| SI: sessile invertebrate feeders | S3: 16–30 cm | SmallG: 3–20 fish on average in a group | VMob: constantly moving around and usually changing reefs within a day | Pelagic: species hovering high above the reef |
| MI: mobile invertebrate feeders | S4: 31–50 cm | MedG: 20–50 fish on average in a group | ||
| PK: plankton feeders | S5: 51–80 cm | LargeG: >50 fish on average in a group | ||
| FC: piscivores | S6: >80 cm |
| 2-Trait FEs | 3-Trait FEs | 4-Trait FEs | 5-Trait FEs |
|---|---|---|---|
| diet-size | diet-size-school | diet-size-school-mobility | diet-size-mobility-school-lwater |
| diet-schooling | diet-size-mobility | diet-size-mobility-lwater | |
| diet-mobility | diet-size-lwater | diet-size-school-lwater | |
| diet-lwater | diet-school-mobility | diet-mobility-school-lwater | |
| size-school | diet-mobility-lwater | size-mobility-school-lwater | |
| size- mobility | diet-school-lwater | ||
| size-lwater | size-mobility-school | ||
| mobility-school | size-mobility-lwater | ||
| mobility-lwater | size-school-lwater | ||
| school-lwater | mobility-school-lwater |
| Df | Sum Sq | Mean Sq | F Value | Pr(>F) | |
|---|---|---|---|---|---|
| Type of functional descriptor | 3 | 0.7586 | 0.2529 | 26.2160 | 1.293 × 10−11 |
| Nb of traits | 1 | 0.2102 | 0.2102 | 21.7897 | 1.349 × 10−5 |
| Type of descriptor x Nb of traits | 3 | 0.0374 | 0.0125 | 1.2921 | 0.2836 |
| Residuals | 73 | 0.7041 | 0.0096 |
| Estimate | Std. Error | t Value | Pr(>|t|) | |
|---|---|---|---|---|
| (Intercept) | 0.48706 | 0.04572 | 10.652 | <2 × 10−16 |
| Shannon index [FE biomass] | −0.23275 | 0.03019 | −7.709 | 3.97 × 10−11 |
| Functional Richness | −0.23399 | 0.02918 | −8.019 | 1.01 × 10−11 |
| Shannon index [FE richness] | −0.18337 | 0.03419 | −5.363 | 8.52 × 10−7 |
| Nb of traits | −0.06185 | 0.01333 | −4.641 | 1.42 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elise, S.; Guilhaumon, F.; Mou-Tham, G.; Urbina-Barreto, I.; Vigliola, L.; Kulbicki, M.; Bruggemann, J.H. Combining Passive Acoustics and Environmental Data for Scaling Up Ecosystem Monitoring: A Test on Coral Reef Fishes. Remote Sens. 2022, 14, 2394. https://doi.org/10.3390/rs14102394
Elise S, Guilhaumon F, Mou-Tham G, Urbina-Barreto I, Vigliola L, Kulbicki M, Bruggemann JH. Combining Passive Acoustics and Environmental Data for Scaling Up Ecosystem Monitoring: A Test on Coral Reef Fishes. Remote Sensing. 2022; 14(10):2394. https://doi.org/10.3390/rs14102394
Chicago/Turabian StyleElise, Simon, François Guilhaumon, Gérard Mou-Tham, Isabel Urbina-Barreto, Laurent Vigliola, Michel Kulbicki, and J. Henrich Bruggemann. 2022. "Combining Passive Acoustics and Environmental Data for Scaling Up Ecosystem Monitoring: A Test on Coral Reef Fishes" Remote Sensing 14, no. 10: 2394. https://doi.org/10.3390/rs14102394