
Improving the Selection of Vegetation Index Characteristic Wavelengths by Using the PROSPECT Model for Leaf Water Content Estimation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Datasets
2.1.1. Simulation Datasets
2.1.2. In Situ Datasets
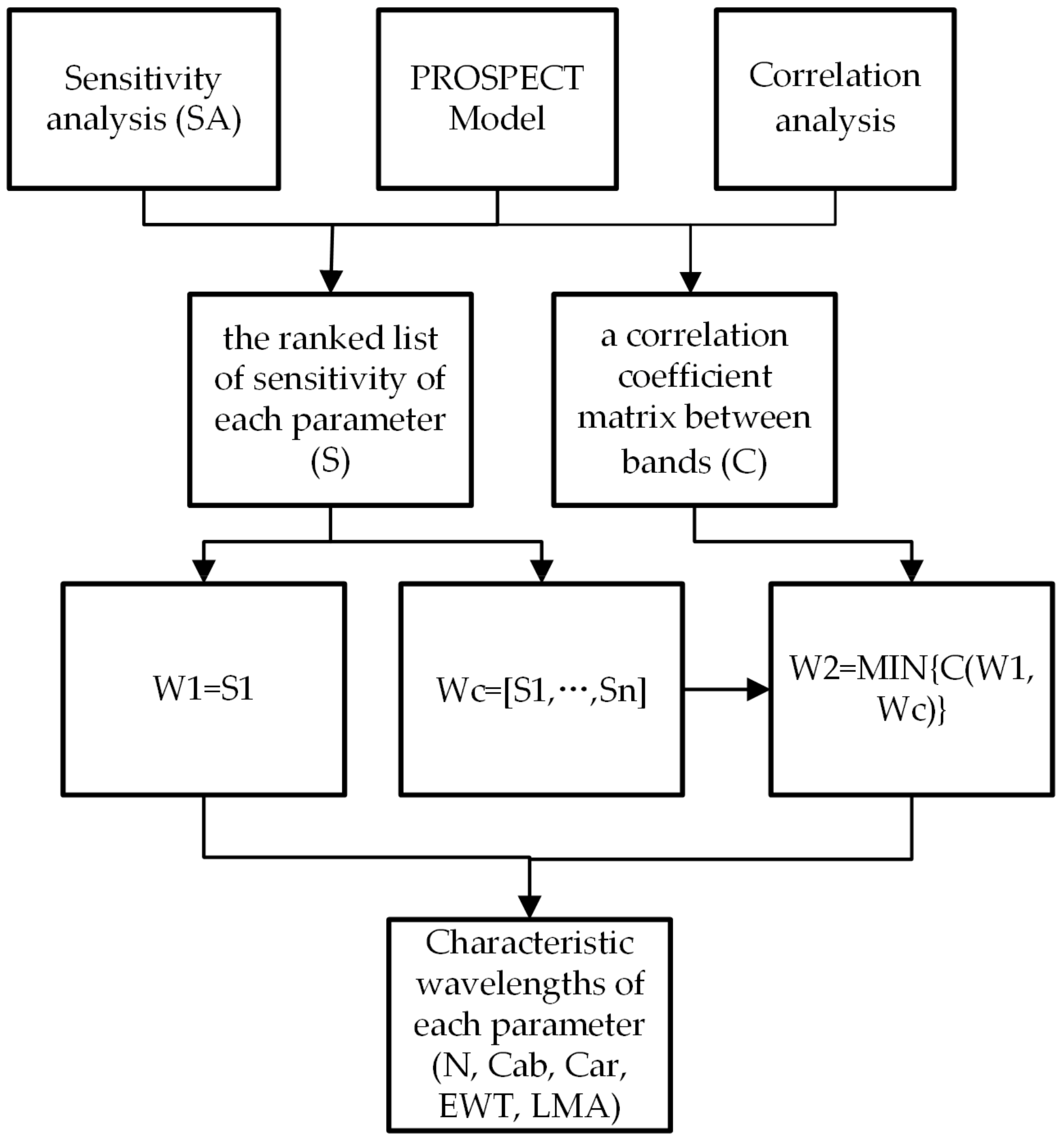
2.2. Characteristic Wavelength Selection Algorithm
2.3. VIs Correlation Analysis and Robustness Analysis
2.4. Gaussian Process Regression
2.5. Statistical Analysis
3. Results
3.1. Selection of Characteristic Wavelengths and nRVI Construction
3.2. Correlation Analysis Between the nRVI, Selected Published VIs, and EWT
3.3. Robustness Analysis
3.3.1. Robustness to the Change of N
3.3.2. Robustness to the Change of LMA
3.3.3. Robustness to the Change of Spectral Noise
3.4. Validation of the Performance of nRVI and Selected VIs for EWT Estimation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ceccato, P.; Flasse, S.; Gregoire, J.M. Designing a spectral index to estimate vegetation water content from remote sensing data—Part 2. Validation and applications. Remote Sens. Environ. 2002, 82, 198–207. [Google Scholar] [CrossRef]
- Ceccato, P.; Gobron, N.; Flasse, S.; Pinty, B.; Tarantola, S. Designing a spectral index to estimate vegetation water content from remote sensing data: Part 1: Theoretical approach. Remote Sens. Environ. 2002, 82, 188–197. [Google Scholar] [CrossRef]
- Stimson, H.C.; Breshears, D.D.; Ustin, S.L.; Kefauver, S.C. Spectral sensing of foliar water conditions in two co-occurring conifer species: Pinus edulis and Juniperus monosperma. Remote Sens. Environ. 2005, 96, 108–118. [Google Scholar] [CrossRef]
- Ullah, S.; Skidmore, A.K.; Groen, T.A.; Schlerf, M. Evaluation of three proposed indices for the retrieval of leaf water content from the mid-wave infrared (2–6 µm) spectra. Agric. For. Meteorol. 2013, 171, 65–71. [Google Scholar] [CrossRef]
- Fang, M.H.; Ju, W.M.; Zhan, W.F.; Cheng, T.; Qiu, F.; Wang, J. A new spectral similarity water index for the estimation of leaf water content from hyperspectral data of leaves. Remote Sens. Environ. 2017, 196, 13–27. [Google Scholar] [CrossRef]
- Cheng, T.; Rivard, B.; Sanchez-Azofeifa, A. Spectroscopic determination of leaf water content using continuous wavelet analysis. Remote Sens. Environ. 2011, 115, 659–670. [Google Scholar] [CrossRef]
- Cheng, T.; Rivard, B.; Sanchez-Azofeifa, A.G.; Feret, J.B.; Jacquemoud, S.; Ustin, S.L. Predicting leaf gravimetric water content from foliar reflectance across a range of plant species using continuous wavelet analysis. J. Plant Physiol. 2012, 169, 1134–1142. [Google Scholar] [CrossRef]
- Penuelas, J.; Pinol, J.; Ogaya, R.; Filella, I. Estimation of plant water concentration by the reflectance water index WI (R900/R970). Int. J. Remote Sens. 1997, 18, 2869–2875. [Google Scholar] [CrossRef]
- Penuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Save, R. The reflectance at the 950-970 Nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Cao, Z.; Wang, Q.; Zheng, C. Best hyperspectral indices for tracing leaf water status as determined from leaf dehydration experiments. Ecol. Indic. 2015, 54, 96–107. [Google Scholar] [CrossRef]
- Ceccato, P.; Flasse, S.; Tarantola, S.; Jacquemoud, S.; Gregoire, J.M. Detecting vegetation leaf water content using reflectance in the optical domain. Remote Sens. Environ. 2001, 77, 22–33. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and physiological basis for the reflectance of visible and near-infrared radiation from vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Thomas, J.R.; Namken, L.N.; Oerther, G.F.; Brown, R.G. Estimating leaf water content by reflectance measurements. Agron. J. 1971, 63, 845–847. [Google Scholar] [CrossRef]
- Tucker, C.J. Remote sensing of leaf water content in the near infrared. Remote Sens. Environ. 1979, 10, 23–32. [Google Scholar] [CrossRef]
- Maki, M.; Ishiahra, M.; Tamura, M. Estimation of leaf water status to monitor the risk of forest fires by using remotely sensed data. Remote Sens. Environ. 2004, 90, 441–450. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Ustin, S.L.; Verdebout, J.; Schmuck, G.; Andreoli, G.; Hosgood, B. Estimating leaf biochemistry using the PROSPECT leaf optical properties model. Remote Sens. Environ. 1996, 56, 194–202. [Google Scholar] [CrossRef]
- Riano, D.; Vaughan, P.; Chuvieco, E.; Zarco-Tejada, P.J.; Ustin, S.L. Estimation of fuel moisture content by inversion of radiative transfer models to simulate equivalent water thickness and dry matter content: Analysis at leaf and canopy level. IEEE Trans. Geosci. Remote 2005, 43, 819–826. [Google Scholar] [CrossRef]
- Colombo, R.; Merom, M.; Marchesi, A.; Busetto, L.; Rossini, M.; Giardino, C.; Panigada, C. Estimation of leaf and canopy water content in poplar plantations by means of hyperspectral indices and inverse modeling. Remote Sens. Environ. 2008, 112, 1820–1834. [Google Scholar] [CrossRef]
- Combal, B.; Baret, F.; Weiss, M.; Trubuil, A.; Mace, D.; Pragnere, A.; Myneni, R.; Knyazikhin, Y.; Wang, L. Retrieval of canopy biophysical variables from bidirectional reflectance—Using prior information to solve the ill-posed inverse problem. Remote Sens. Environ. 2003, 84, 1–15. [Google Scholar] [CrossRef]
- Jacquemoud, S. Inversion of the PROSPECT + SAIL canopy reflectance model from AVIRIS equivalent spectra: Theoretical-Study. Remote Sens. Environ. 1993, 44, 281–292. [Google Scholar] [CrossRef]
- Allen, W.A.; Gausman, H.W.; Richardson, A.J.; Thomas, J.R. Interaction of isotropic light with a compact plant leaf. JOSA 1969, 59, 1376–1379. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F. Prospect: A model of leaf optical-properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Feret, J.-B.; François, C.; Asner, G.P.; Gitelson, A.A.; Martin, R.E.; Bidel, L.P.R.; Ustin, S.L.; le Maire, G.; Jacquemoud, S. PROSPECT-4 and 5: Advances in the leaf optical properties model separating photosynthetic pigments. Remote Sens. Environ. 2008, 112, 3030–3043. [Google Scholar] [CrossRef]
- Feret, J.B.; Gitelson, A.A.; Noble, S.D.; Jacquemoud, S. PROSPECT-D: Towards modeling leaf optical properties through a complete lifecycle. Remote Sens. Environ. 2017, 193, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Comar, A.; Burger, P.; Bancal, P.; Weiss, M.; Baret, F. Estimation of leaf traits from reflectance measurements: Comparison between methods based on vegetation indices and several versions of the PROSPECT model. Plant Methods 2018, 14, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Cheng, T.; Jia, M.; Zhou, K.; Lu, N.; Yao, X.; Tian, Y.C.; Zhu, Y.; Cao, W.X. PROCWT: Coupling PROSPECT with continuous wavelet transform to improve the retrieval of foliar chemistry from leaf bidirectional reflectance spectra. Remote Sens. Environ. 2018, 206, 1–14. [Google Scholar] [CrossRef]
- Yang, J.; Yang, S.X.; Zhang, Y.Y.; Shi, S.; Du, L. Improving characteristic band selection in leaf biochemical property estimation considering interrelations among biochemical parameters based on the PROSPECT-D model. Opt. Express 2021, 29, 400–414. [Google Scholar] [CrossRef]
- Yilmaz, M.T.; Hunt, E.R.; Jackson, T.J. Remote sensing of vegetation water content from equivalent water thickness using satellite imagery. Remote Sens. Environ. 2008, 112, 2514–2522. [Google Scholar] [CrossRef]
- Datt, B. Remote sensing of water content in Eucalyptus leaves. Aust. J. Bot. 1999, 47, 909–923. [Google Scholar] [CrossRef]
- Eitel, J.U.H.; Gessler, P.E.; Smith, A.M.S.; Robberecht, R. Suitability of existing and novel spectral indices to remotely detect water stress in Populus spp. Forest Ecol. Manag. 2006, 229, 170–182. [Google Scholar] [CrossRef]
- Le Maire, G.; Francois, C.; Dufrene, E. Towards universal broad leaf chlorophyll indices using PROSPECT simulated database and hyperspectral reflectance measurements. Remote Sens. Environ. 2004, 89, 1–28. [Google Scholar] [CrossRef]
- Féret, J.-B.; François, C.; Gitelson, A.; Asner, G.P.; Barry, K.M.; Panigada, C.; Richardson, A.D.; Jacquemoud, S. Optimizing spectral indices and chemometric analysis of leaf chemical properties using radiative transfer modeling. Remote Sens. Environ. 2011, 115, 2742–2750. [Google Scholar] [CrossRef] [Green Version]
- Behmann, J.; Mahlein, A.K.; Rumpf, T.; Romer, C.; Plumer, L. A review of advanced machine learning methods for the detection of biotic stress in precision crop protection. Precis. Agric. 2015, 16, 239–260. [Google Scholar] [CrossRef]
- Cipollini, P.; Corsini, G.; Diani, M.; Grasso, R. Retrieval of sea water optically active parameters from hyperspectral data by means of generalized radial basis function neural networks. IEEE Trans. Geosci. Remote Sens. Lett. 2001, 39, 1508–1524. [Google Scholar] [CrossRef]
- Verrelst, J.; Munoz, J.; Alonso, L.; Delegido, J.; Rivera, J.P.; Camps-Valls, G.; Moreno, J. Machine learning regression algorithms for biophysical parameter retrieval: Opportunities for Sentinel-2 and -3. Remote Sens. Environ. 2012, 118, 127–139. [Google Scholar] [CrossRef]
- Camps-Valls, G.; Bruzzone, L.; Rojo-Alvarez, J.L.; Melgani, F. Robust support vector regression for biophysical variable estimation from remotely sensed images. IEEE Geosci. Remote Sens. Lett. 2006, 3, 339–343. [Google Scholar] [CrossRef]
- Verrelst, J.; Alonso, L.; Caicedo, J.P.R.; Moreno, J.; Camps-Valls, G. Gaussian process retrieval of chlorophyll content from imaging spectroscopy data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2013, 6, 867–874. [Google Scholar] [CrossRef]
- Demarez, V.; Mougin, E.; Marty, G.; Proisy, C.; Dufrene, E.; Le, D.V.; Gastellu-Etchegorry, J.P. Seasonal variation of leaf chlorophyll content of a temperate forest. Inversion of the PROSPECT model. Int. J. Remote Sens. 1999, 20, 879–894. [Google Scholar] [CrossRef]
- Sun, J.; Shi, S.; Yang, J.; Gong, W.; Qiu, F.; Wang, L.; Du, L.; Chen, B. Wavelength selection of the multispectral lidar system for estimating leaf chlorophyll and water contents through the PROSPECT model. Agric. Forest Meteorol. 2019, 266-267, 43–52. [Google Scholar] [CrossRef]
- Hosgood, B.; Jacquemoud, S.; Andreoli, G. Leaf Optical Properties EXperiment 93 (LOPEX93); Ispra Italy’European Commission, Joint Research Centre Institute of Remote Sensing Applications: Luxembourg, 1994. [Google Scholar]
- Martin, J.D.A.A.M.E. Leaf Chemistry, 1992–1993 (ACCP); Oak Ridge National Laboratory: Oak Ridge, TN, USA, 1999. [Google Scholar]
- Grossman, Y.L.; Ustin, S.L.; Jacquemoud, S.; Sanderson, E.W.; Schmuck, G.; Verdebout, J. Critique of stepwise multiple linear regression for the extraction of leaf biochemistry information from leaf reflectance data. Remote Sens. Environ. 1996, 56, 182–193. [Google Scholar] [CrossRef]
- Bowyer, P.; Danson, F.M. Sensitivity of spectral reflectance to variation in live fuel moisture content at leaf and canopy level. Remote Sens. Environ. 2004, 92, 297–308. [Google Scholar] [CrossRef]
- Cannavó, F. Sensitivity analysis for volcanic source modeling quality assessment and model selection. Comput. Geosci. 2012, 44, 52–59. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Rock, B.N. Detection of changes in leaf water content using Near- and Middle-Infrared reflectance. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar]
- Seelig, H.D.; Hoehn, A.; Stodieck, L.S.; Klaus, D.M.; Adams, W.W.; Emery, W.J. The assessment of leaf water content using leaf reflectance ratios in the visible, near-, and short-wave-infrared. Int. J. Remote Sens. 2008, 29, 3701–3713. [Google Scholar] [CrossRef]
- Yang, J.; Du, L.; Gong, W.; Shi, S.; Sun, J.; Chen, B. Potential of vegetation indices combined with laser-induced fluorescence parameters for monitoring leaf nitrogen content in paddy rice. PLoS ONE 2018, 13, e0191068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstraete, M.M.; Pinty, B. Designing optimal spectral indexes for remote sensing applications. IEEE Trans. Geosci. Remote Sens. 1996, 34, 1254–1265. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Verrelst, J.; Alonso, L.; Camps-Valls, G.; Delegido, J.; Moreno, J. Retrieval of vegetation biophysical parameters using gaussian process techniques. IEEE Trans. Geosci. Remote Sens. 2011, 50, 1832–1843. [Google Scholar] [CrossRef]
- Karimi, S.; Sadraddini, A.A.; Nazemi, A.H.; Xu, T.; Fard, A.F. Generalizability of gene expression programming and random forest methodologies in estimating cropland and grassland leaf area index. Comput. Electron. Agric. 2018, 144, 232–240. [Google Scholar] [CrossRef]
- Durbha, S.S.; King, R.L.; Younan, N.H. Support vector machines regression for retrieval of leaf area index from multiangle imaging spectroradiometer. Remote Sens. Environ. 2007, 107, 348–361. [Google Scholar] [CrossRef]
- Bacour, C.; Baret, F.; Béal, D.; Weiss, M.; Pavageau, K. Neural network estimation of LAI, fAPAR, fCover and LAI × C ab, from top of canopy MERIS reflectance data: Principles and validation. Remote Sens. Environ. 2006, 105, 313–325. [Google Scholar] [CrossRef]
- Verrelst, J.; Rivera, J.P.; Gitelson, A.; Delegido, J.; Moreno, J.; Camps-Valls, G. Spectral band selection for vegetation properties retrieval using Gaussian processes regression. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 554–567. [Google Scholar] [CrossRef]
- Lazaro-Gredilla, M.; Titsias, M.K.; Verrelst, J.; Camps-Valls, G. Retrieval of biophysical parameters with heteroscedastic gaussian processes. IEEE Geosci. Remote Sens. 2014, 11, 838–842. [Google Scholar] [CrossRef]
- Camps-Valls, G.; Gomez-Chova, L.; Muoz-Mari, J.; Vila-Frances, J.; Amoros, J.; Valle-Tascon, S.D.; Calpe-Maravilla, J. Biophysical parameter estimation with adaptive Gaussian Processes. In Proceedings of the Geoscience & Remote Sensing Symposium, Cape Town, South Africa, 12–17 July 2009. [Google Scholar]
- Verrelst, J.; Rivera, J.P.; Moreno, J.; Camps-Valls, G. Gaussian processes uncertainty estimates in experimental Sentinel-2 LAI and leaf chlorophyll content retrieval. ISPRS J. Photogramm. Remote Sens. 2013, 86, 157–167. [Google Scholar] [CrossRef]
- Huang, X.; Xu, Q.S.; Liang, Y.Z. PLS regression based on sure independence screening for multivariate calibration. Anal. Methods 2012, 4, 2815–2821. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Noland, T.L.; Mohammed, G.H.; Sampson, P.H. Scaling-up and model inversion methods with narrowband optical indices for chlorophyll content estimation in closed forest canopies with hyperspectral data. IEEE Trans. Geosci. Remote Sens. 2001, 39, 1491–1507. [Google Scholar] [CrossRef] [Green Version]
- Song, S.L.; Gong, W.; Zhu, B.; Huang, X. Wavelength selection and spectral discrimination for paddy rice, with laboratory measurements of hyperspectral leaf reflectance. ISPRS J. Photogramm. Remote Sens. 2011, 66, 672–682. [Google Scholar] [CrossRef]
- He, L.; Song, X.; Feng, W.; Guo, B.B.; Zhang, Y.S.; Wang, Y.H.; Wang, C.Y.; Guo, T.C. Improved remote sensing of leaf nitrogen concentration in winter wheat using multi-angular hyperspectral data. Remote Sens. Environ. 2016, 174, 122–133. [Google Scholar] [CrossRef]
- Hardisky, M.A.; Klemas, V.; Smart, R.M. The influence of soil-salinity, growth form, and leaf moisture on the spectral radiance of spartina-alterniflora canopies. Photogramm. Eng. Remote Sensing 1983, 49, 77–83. [Google Scholar]
- Asner, G.P.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Chen, J.M.; Miller, J.R.; Noland, T.L. Leaf chlorophyll content retrieval from airborne hyperspectral remote sensing imagery. Remote Sens. Environ. 2008, 112, 3234–3247. [Google Scholar] [CrossRef]
- Simic, A.; Chen, J.M.; Leblanc, S.G.; Dyk, A.; Croft, H.; Han, T. Testing the top-down model inversion method of estimating leaf reflectance used to retrieve vegetation biochemical content within empirical approaches. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 92–104. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Chlorophyll (Cab) (μg/cm2) | Carotenoid (Car) (μg/cm2) | Equivalent Water Thickness (EWT) (cm) | Dry Matter Content (LMA) (g/cm2) | |
|---|---|---|---|---|---|
| Mean value | 1.6 | 32.81 | 8.51 | 0.0129 | 0.0077 |
| Standard deviation | 0.3 | 18.87 | 3.92 | 0.0073 | 0.0035 |
| Minimum | 1 | 0.30 | 0.04 | 0.00005 | 0.002 |
| Maximum | 3.5 | 110 | 30 | 0.07 | 0.04 |
| Datasets | Synthetic Spectrum without Noise | Synthetic Spectrum with 2% Random Gaussian Noise | ANGERS | LOPEX | JR |
|---|---|---|---|---|---|
| Vegetation indices | WBI | MSI | SR | NDII | SR |
| Formula | (R900/R970) | (R1600/R820) | (R1300/R1450) | (R820-R1600)/(R820 + R1600) | (R1300/R1450) |
| R2 | 0.952 | 0.681 | 0.909 | 0.828 | 0.701 |
| References | [9] | [40] | [41] | [42] | [41] |
| N | Cab | Car | EWT | LMA | |
|---|---|---|---|---|---|
| Characteristic wavelengths | 1089 nm, 746 nm | 675 nm, 595 nm | 520 nm, 400 nm | 1906 nm, 1398 nm | 2286 nm, 2311 nm |
| Characteristic Wavelengths-EWT | Characteristic Wavelengths-EWT + N | Characteristic Wavelengths- EWT + Cab | Characteristic Wavelengths- EWT + Car | Characteristic Wavelengths-EWT + LMA | ||
|---|---|---|---|---|---|---|
| Synthetic spectrum without noise | R2 | 0.9686 | 0.9989 | 0.9719 | 0.9729 | 0.9629 |
| Root mean square error (RMSE) | 0.0012 | 0.0002 | 0.0011 | 0.0011 | 0.0011 | |
| Normalized RMSE (NRMSE) | 8.2270 | 1.4115 | 7.8654 | 7.7230 | 7.8063 | |
| Synthetic spectrum with 2% random Gaussian noise | R2 | 0.6728 | 0.8567 | 0.6842 | 0.6910 | 0.6766 |
| RMSE | 0.0038 | 0.0025 | 0.0037 | 0.0037 | 0.0038 | |
| NRMSE | 26.8854 | 17.8595 | 26.3449 | 26.1832 | 26.9376 | |
| ANGERS | R2 | 0.7536 | 0.9243 | 0.7821 | 0.7166 | 0.7892 |
| RMSE | 0.0024 | 0.0013 | 0.0023 | 0.0026 | 0.0022 | |
| NRMSE | 20.7514 | 11.4918 | 19.4984 | 22.4114 | 19.1916 | |
| LOPEX | R2 | 0.6975 | 0.9199 | 0.6912 | 0.6907 | 0.7090 |
| RMSE | 0.0038 | 0.0019 | 0.0039 | 0.0038 | 0.0037 | |
| NRMSE | 32.9737 | 16.8742 | 33.6663 | 33.5077 | 32.2661 | |
| JR | R2 | 0.3543 | 0.7871 | 0.4885 | 0.5829 | 0.3312 |
| RMSE | 0.0052 | 0.0030 | 0.0048 | 0.0042 | 0.0053 | |
| NRMSE | 32.3883 | 18.5350 | 30.1157 | 26.5135 | 33.1941 |
| Full-Wavelengths (400–2500 nm) | Full-Characteristic Wavelengths (10) | Characteristic Wavelengths-EWT + N (4) | Characteristic Wavelengths-EWT (2) | ||
|---|---|---|---|---|---|
| Synthetic spectrum without noise | R2 | 0.9979 | 0.9999 | 0.9989 | 0.9686 |
| RMSE | 0.0001 | 0.0003 | 0.0002 | 0.0012 | |
| NRMSE | 0.9712 | 0.2407 | 1.4115 | 8.2270 | |
| Synthetic spectrum with 2% random Gaussian noise | R2 | 0.1205 | 0.8951 | 0.8567 | 0.6728 |
| RMSE | 0.0156 | 0.0022 | 0.0025 | 0.0038 | |
| NRMSE | 110.4656 | 15.3237 | 17.8595 | 26.8854 | |
| ANGERS | R2 | 0.5170 | 0.9265 | 0.9243 | 0.7536 |
| RMSE | 0.0036 | 0.0013 | 0.0013 | 0.0024 | |
| NRMSE | 30.8190 | 11.3459 | 11.4918 | 20.7514 | |
| LOPEX | R2 | 0.8682 | 0.9012 | 0.9199 | 0.6975 |
| RMSE | 0.0025 | 0.0021 | 0.0019 | 0.0038 | |
| NRMSE | 21.7422 | 18.7645 | 16.8742 | 32.9737 | |
| JR | R2 | 0.4162 | 0.7695 | 0.7871 | 0.3543 |
| RMSE | 0.0065 | 0.0031 | 0.0030 | 0.0052 | |
| NRMSE | 40.3687 | 19.5815 | 18.5350 | 32.3883 |
| Datasets | Vegetation Indices | |||||
|---|---|---|---|---|---|---|
| nRVI | WBI | MSI | SR | NDII | ||
| Synthetic spectrum without noise | R2 | 0.9544 | 0.9518 | 0.9233 | 0.9423 | 0.9219 |
| RMSE | 0.0014 | 0.0015 | 0.0019 | 0.0016 | 0.0019 | |
| NRMSE | 10.0359 | 10.3498 | 13.0451 | 11.2950 | 13.1296 | |
| Synthetic spectrum with 2% random Gaussian noise | R2 | 0.8188 | 0.0320 | 0.6931 | 0.7554 | 0.6904 |
| RMSE | 0.0028 | 0.0066 | 0.0037 | 0.0033 | 0.0037 | |
| NRMSE | 20.2391 | 47.0444 | 26.2381 | 23.3033 | 26.5343 | |
| ANGERS | R2 | 0.9284 | 0.8280 | 0.8982 | 0.9097 | 0.8948 |
| RMSE | 0.0013 | 0.0020 | 0.0015 | 0.0015 | 0.0016 | |
| NRMSE | 11.1337 | 17.3393 | 13.3377 | 12.5704 | 13.5613 | |
| LOPEX | R2 | 0.8938 | 0.7363 | 0.9023 | 0.7304 | 0.9023 |
| RMSE | 0.0022 | 0.0035 | 0.0021 | 0.0036 | 0.0021 | |
| NRMSE | 19.5856 | 30.7482 | 18.7015 | 31.2757 | 18.7120 | |
| JR | R2 | 0.7766 | 0.5331 | 0.4950 | 0.6602 | 0.4991 |
| RMSE | 0.0030 | 0.0043 | 0.0044 | 0.0036 | 0.0044 | |
| NRMSE | 18.4784 | 27.0536 | 27.7635 | 22.7170 | 27.6439 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Zhang, Y.; Du, L.; Liu, X.; Shi, S.; Chen, B. Improving the Selection of Vegetation Index Characteristic Wavelengths by Using the PROSPECT Model for Leaf Water Content Estimation. Remote Sens. 2021, 13, 821. https://doi.org/10.3390/rs13040821
Yang J, Zhang Y, Du L, Liu X, Shi S, Chen B. Improving the Selection of Vegetation Index Characteristic Wavelengths by Using the PROSPECT Model for Leaf Water Content Estimation. Remote Sensing. 2021; 13(4):821. https://doi.org/10.3390/rs13040821
Chicago/Turabian StyleYang, Jian, Yangyang Zhang, Lin Du, Xiuguo Liu, Shuo Shi, and Biwu Chen. 2021. "Improving the Selection of Vegetation Index Characteristic Wavelengths by Using the PROSPECT Model for Leaf Water Content Estimation" Remote Sensing 13, no. 4: 821. https://doi.org/10.3390/rs13040821