
Wearable Near-Infrared Spectroscopy as a Physiological Monitoring Tool for Seals under Anaesthesia

 , , , , , , , and
, , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Methods
2.2.1. Drugging Procedure
2.2.2. Instrumentation
2.2.3. Data Collection
2.3. Data Extraction
2.3.1. Extraction of Haemodynamic Variables ([ΔO2Hb] and [ΔHHb])
2.3.2. Extraction of Respiratory Data
2.3.3. Heart Rate Extraction
2.3.4. Arterial Oxygen Saturation (SpO2) Extraction
2.4. Statistical Modelling
3. Results
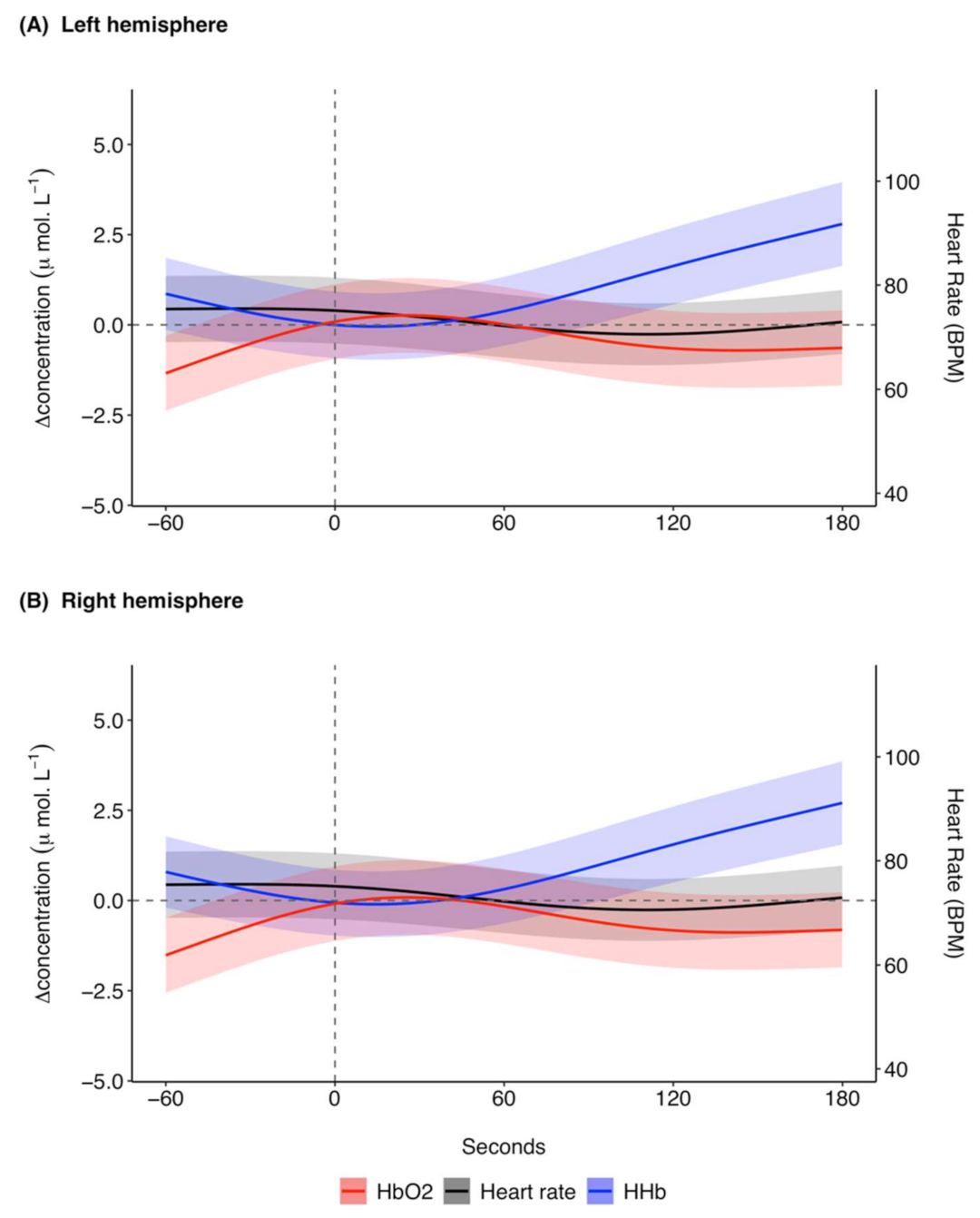
3.1. Ketamine
3.1.1. Raw Data
3.1.2. Model Results
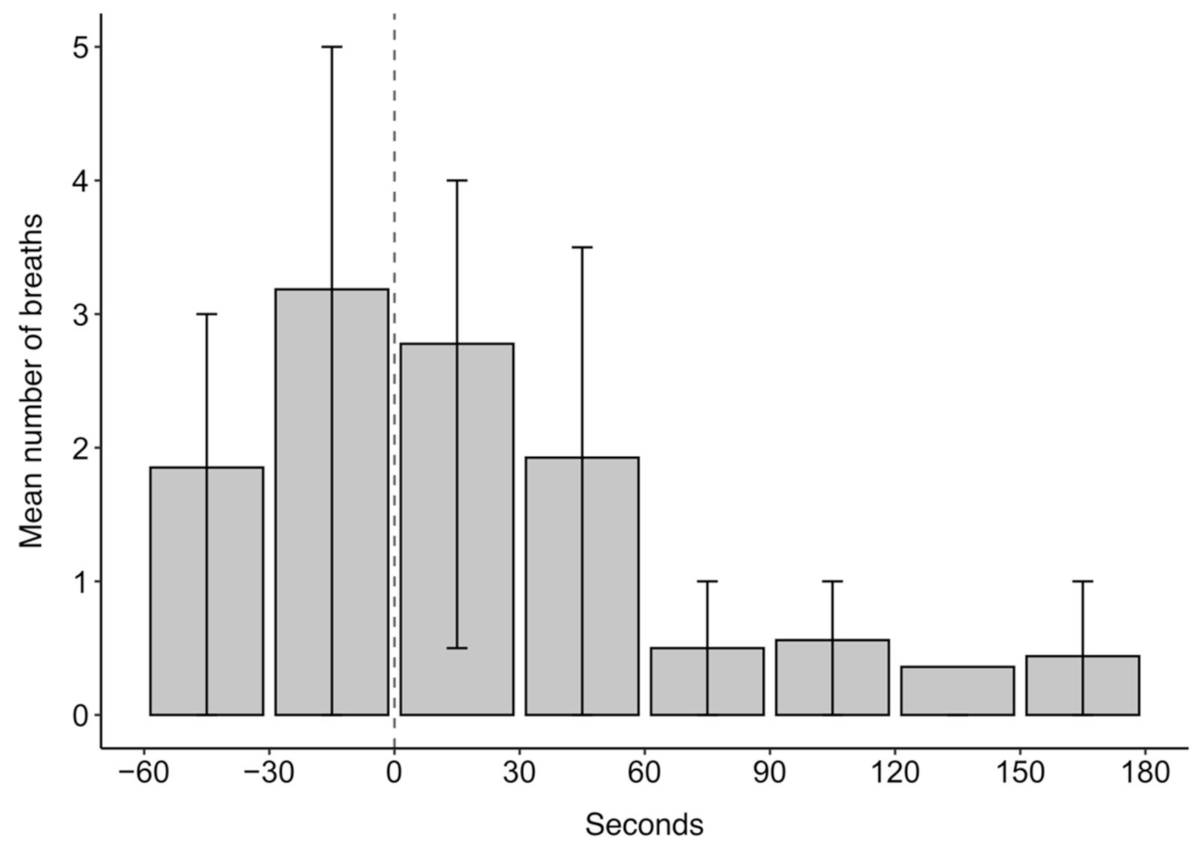
3.1.3. Breathing Rates
3.2. Midazolam
3.2.1. Raw Data
3.2.2. Model Results
3.2.3. Breathing Rates
4. Discussion
4.1. NIRS as a Monitoring Tool during Anaesthesia
4.2. Physiological Responses to Drugging
4.3. Additional Modelling Interpretations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McIntyre, T. Trends in tagging of marine mammals: A review of marine mammal biologging studies. Afr. J. Mar. Sci. 2014, 36, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Dierauf, L.A.; Gulland, F.M.D. (Eds.) CRC Handbook of Marine Mammal Medicine: Health, Disease, and Rehabilitation, 2nd ed.; CRC Press: Danvers, MA, USA, 2001. [Google Scholar]
- Gales, N.J. Chemical restraint and anesthesia of pinnipeds: A review. Mar. Mammal Sci. 1989, 5, 228–256. [Google Scholar] [CrossRef]
- Jones, D.R.; Fisher, H.D.; McTaggart, S.; West, N.H. Heart rate during breath-holding and diving in the unrestrained harbor seal (Phoca vitulina richardi). Can. J. Zool. 1973, 51, 671–680. [Google Scholar] [CrossRef]
- McKnight, J.C.; Bennett, K.A.; Bronkhorst, M.; Russell, D.J.F.; Balfour, S.; Milne, R.; Bivins, M.; Moss, S.E.W.; Colier, W.; Hall, A.J.; et al. Shining new light on mammalian diving physiology using wearable near-infrared spectroscopy. PLoS Biol. 2019, 17, e3000306. [Google Scholar] [CrossRef]
- Ridgway, S.H.; Harrison, R.J.; Joyce, P.L. Sleep and cardiac rhythm in the gray seal. Science 1975, 187, 553–555. [Google Scholar] [CrossRef]
- Gales, N.; Hindell, M.; Kirkwood, R. (Eds.) Marine Mammals: Fisheries, Tourism and Management Issues; CSIRO Publishing: Collingwood, VIC, Australia, 2003. [Google Scholar]
- Brunson, D.B. Comparative anesthesia and analgesia of aquatic mammals. In Veterinary Anaesthesia and Analgesia; Grimm, K.A., Lamont, L.A., Tranquili, W.J., Greene, S.A., Robertson, S.A., Eds.; John Wiley & Sons: Ames, IA, USA, 2015; pp. 777–783. [Google Scholar]
- Lynch, M.; Bodley, K. Phocid seals. In Immobilization and Anesthesia Zoo Animal & Wildlife; West, G., Heard, D., Caulkett, N., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 459–468. [Google Scholar]
- Lynch, M.J.; Thamindjis, M.A.; Gardner, H. Immobilisation of pinniped species. Aust. Vet. J. 1999, 77, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Baylis, A.M.M.; Page, B.; Staniland, I.; Arnould, J.P.Y.; McKenzie, J. Taking the sting out of darting: Risks, restraint drugs and procedures for the chemical restraint of Southern Hemisphere otariids. Mar. Mammal Sci. 2015, 31, 322–344. [Google Scholar] [CrossRef]
- Wolff, K.; Winstock, A.R. Ketamine: From medicine to misuse. CNS Drugs 2006, 20, 199–218. [Google Scholar] [CrossRef]
- Sinner, B.; Graf, B.M. Ketamine. In Handbook of Experimental Pharmacology: Modern Anesthetics; Schüttler, J., Schwilden, H., Eds.; Springer: Berlin/Heidelberg, Germay, 2008; Volume 182, pp. 313–335. [Google Scholar]
- Briggs, G.D.; Henrickson, R.V.; Burney, D.V.M.; Le Boeuf, J. Ketamine immobilization of northern elephant seals. J. Am. Vet. Med. Assoc. 1975, 167, 546–548. [Google Scholar]
- Olkkola, K.T.; Ahonen, J. Midazolam and other benzodiazepines. In Handbook of Experimental Pharmacology: Modern Anesthetics; Schüttler, J., Schwilden, H., Eds.; Springer: Heidelberg, Germany, 2008; Volume 182, pp. 335–360. [Google Scholar] [CrossRef]
- Saldman, L.J. Midazolam: Pharmacology and uses. Anesthesiology 1985, 62, 310–324. [Google Scholar]
- Parry, K.; Anderson, S.S.; Fedak, M.A. Chemical immobilization of gray seals. J. Wildl. Manag. 1981, 45, 986–990. [Google Scholar] [CrossRef]
- Dabin, W.; Beauplet, G.; Guinet, C. Response of wild subantarctic fur seal (Arctocephalus tropicalis) females to ketamine and tiletamine-zolazepam anesthesia. J. Wildl. Dis. 2002, 38, 846–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, J.; Reichmuth, C.; McDonald, B.I.; Kristensen, J.H.; Larson, J.; Johansson, F.; Sullivan, J.L.; Madsen, P.T. Drivers of the dive response in pinnipeds; apnea, submergence or temperature? J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntyre, T.; de Bruyn, P.J.N.; Ansorge, I.J.; Bester, M.N.; Bornemann, H.; Plötz, J.; Tosh, C.A. A lifetime at depth: Vertical distribution of southern elephant seals in the water column. Polar Biol. 2010, 33, 1037–1048. [Google Scholar] [CrossRef]
- Hindell, M.A.; Slip, D.J.; Burton, H.R. The diving behaviour of adult male and female southern elephant seals, Mirounga leonina (Pinnipedia, Phocidae). Aust. J. Zool. 1991, 39, 595–619. [Google Scholar] [CrossRef]
- Hindell, M.A.; Slip, D.J.; Burton, H.R.; Bryden, M.M. Physiological implications of continuous, prolonged, and deep dives of the southern elephant seal (Mirounga leonina). Can. J. Zool. 1992, 70, 370–379. [Google Scholar] [CrossRef]
- Schytte Blix, A. Adaptations to deep and prolonged diving in phocid seals. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Blix, A.S.; Elsner, R.; Kjekhus, J.K. Cardiac output and its distribution through capillaries and A-V shunts in diving seals. Acta Physiol. Scand. 1983, 118, 109–116. [Google Scholar] [CrossRef]
- Ridgway, S.H.; Carder, D.A.; Clark, W. Conditioned bradycardia in the sea lion Zalophus californianus. Nature 1975, 256, 37–38. [Google Scholar] [CrossRef]
- Elmegaard, S.L.; Johnson, M.; Madsen, P.T.; McDonald, B.I. Cognitive control of heart rate in diving harbor porpoises. Curr. Biol. 2016, 26, 1175–1176. [Google Scholar] [CrossRef] [Green Version]
- Fahlman, A.; Cozzi, B.; Manley, M.; Jabas, S.; Malik, M.; Blawas, A.; Janik, V.M. Conditioned variation in heart rate during static breath-holds in the bottlenose dolphin (Tursiops truncatus). Front. Physiol. 2020, 11, 1509. [Google Scholar] [CrossRef]
- Murkin, J.M.; Arango, M. Near-infrared spectroscopy as an index of brain and tissue oxygenation. Br. J. Anaesth. 2009, 103, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Fantini, S.; Ruesch, A.; Kainerstorfer, J.M. Noninvasive optical studies of the brain: Contributions from systemic physiology. In Neurophotonics and Biomedical Spectroscopy; Alfano, R.R., Shi, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 25–52. [Google Scholar]
- Suzuki, S.; Takasaki, S.; Ozaki, T.; Kobayashi, Y. A tissue oxygenation monitor using NIR spatially resolved spectroscopy. In SPIE, Optical Tomography and Spectroscopy of Tissue III; SPIE: Bellingham, WA, USA, 1999; Volume 3597, pp. 582–592. [Google Scholar] [CrossRef]
- McKnight, J.C.; Ruesch, A.; Bennett, K.; Bronkhorst, M.; Balfour, S.; Moss, S.E.W.; Milne, R.; Tyack, P.L.; Kainerstorfer, J.M.; Hastie, G.D. Shining new light on sensory brain activation and physiological measurement in seals using wearable optical technology. Philos. Trans. R. Soc. B 2021, 376, 20200224. [Google Scholar] [CrossRef]
- Oller, L.; Bennett, K.A.; McKnight, J.C.; Moss, E.W.; Milne, R.; Hall, A.J.; Rocha, J. Partial pressure of oxygen in adipose tissue and its relationship with fatness in a natural animal model of extreme fat deposition, the grey seal. Physiol. Rep. 2021, 9, e14972. [Google Scholar] [CrossRef]
- Reynolds, E.O.R.; Wyatt, J.S.; Azzopardi, D.; Delpy, D.T.; Cady, E.B.; Cope, M.; Wray, S. New non-invasive methods for assessing brain oxygenation and haemodynamics. Br. Med. Bull. 1988, 44, 1052–1075. [Google Scholar] [CrossRef] [Green Version]
- Delpy, D.T.; Cope, M.; van der Zee, P.; Arridge, S.; Wray, S.; Wyatt, J. Estimation of optical pathlength through tissue from direct time of flight measurement. Phys. Med. Biol. 1988, 33, 1433–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, S.M.; Roback, M.G.; Kennedy, R.M.; Krauss, B. Clinical practice guideline for emergency department ketamine dissociative sedation: 2011 update. Ann. Emerg. Med. 2011, 57, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Laude, D.; Goldman, M.; Escourrou, P.; Elghozi, J.-L. Effect of breathing pattern on blood pressure and heart rate oscillations in humans. Clin. Exp. Pharmacol. Physiol. 1993, 20, 619–626. [Google Scholar] [CrossRef]
- Reed, J.Z.; Chambers, C.; Fedak, M.A.; Butler, P.J. Gas exchange of captive freely diving grey seals (Halichoerus grypus). J. Exp. Biol. 1994, 191, 1–18. [Google Scholar] [CrossRef] [PubMed]
- McKnight, J.C.; Mulder, E.; Ruesch, A.; Kairnerstorfer, J.; Wu, J.; Hakimi, N.; Balfour, S.; Bronkhorst, M.; Horschig, J.M.; Pernett, F.; et al. When the human brain goes diving: Using near-infrared spectroscopy to measure cerebral and systemic cardiovascular responses to deep, breath-hold diving in elite freedivers. Philos. Trans. R. Soc. B 2021, 376, 20200349. [Google Scholar] [CrossRef]
- Hakimi, N.; Setarehdan, S.K. Stress assessment by means of heart rate derived from functional near- infrared spectroscopy. J. Biomed. Opt. 2018, 23, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Scholkmann, F.; Boss, J.; Wolf, M. An efficient algorithm for automatic peak detection in noisy periodic and quasi-periodic signals. Algorithms 2012, 5, 588–603. [Google Scholar] [CrossRef] [Green Version]
- Menssen, J.; Colier, W.; Hopman, J.; Liem, D.; de Korte, C. A method to calculate arterial and venous saturation from near infrared spectroscopy (NIRS). In Oxygen Transport to Tissue XXX. Advances in Experimental Medicine and Biology; Liss, P., Hansell, P., Bruley, D.F., Harrison, D.K., Eds.; Springer: Boston, MA, USA, 2009; Volume 645, pp. 135–140. [Google Scholar] [CrossRef]
- Wood, S.N. Package ‘mgcv’. 2021. Available online: https://cran.r-project.org/web/packages/mgcv/mgcv.pdf (accessed on 20 August 2021).
- Pang, D.S.J.; Rondenay, Y.; Measures, L.; Lair, S. The effects of two dosages of midazolam on short-duration anesthesia in the harp seal (Phoca groenlandica). J. Zoo Wildl. Med. 2006, 37, 27–32. [Google Scholar] [CrossRef]
- Mitchell, P.; Burton, H. Immobilisation of southern elephant seals and leopard seals with cyclohexamine anaesthetics and xylazine. Vet. Rec. 1991, 129, 332–336. [Google Scholar] [CrossRef]
- Woods, R.; Hindell, M.; Slip, D.J. Effects of physiological state on duration of sedation in southern elephant seals. J. Wildl. Dis. 1989, 25, 586–590. [Google Scholar] [CrossRef] [Green Version]
- Geraci, J.R. An appraisal of ketamine as an immobilizing agent in wild and captive pinnipeds. J. Am. Vet. Med. Assoc. 1973, 163, 574–577. [Google Scholar] [PubMed]
- Ferrari, M.; Quaresima, V. A brief review on the history of human functional near-infrared spectroscopy (fNIRS) development and fields of application. Neuroimage 2012, 63, 921–935. [Google Scholar] [CrossRef]
- Valencia, L.; Rodríguez-Pérez, A.; Kühlmorgen, B.; Santana, R.Y. Does sevoflurane preserve regional cerebral oxygen saturation measured by near-infrared spectroscopy better than propofol? Ann. Fr. Anesth. Reanim. 2014, 33, 59–65. [Google Scholar] [CrossRef]
- Iwasaki, K.; Nomoto, Y.; Ishiwata, M.; Yokota, T.; Ogawa, R. Vital capacity induction with 8% sevoflurane and N2O causes cerebral hyperemia. J. Anesth. 2003, 17, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Lovell, A.T.; Marshall, A.C.; Elwell, C.E.; Smith, M.; Goldstone, J.C. Changes in cerebral blood volume with changes in position in awake and anesthetized subjects. Anesth. Analg. 2000, 90, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kwon, J.Y.; Cho, A.R.; Kim, H.K.; Kim, T.K. The effects of sevoflurane and propofol anesthesia on cerebral oxygenation in gynecological laparoscopic surgery. Korean J. Anesthesiol. 2011, 61, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, T.; Okuda, Y.; Yamaguchi, S.; Takanishi, T.; Kumagai, M.; Ido, K. Response of cerebral oxygen metabolism in the head-up position during laparoscopic cholecystectomy. Surg. Laparosc. Endosc. 1998, 8, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Saager, R.B.; Berger, A.J. Measurement of layer-like hemodynamic trends in scalp and cortex: Implications for physiological baseline suppression in functional near-infrared spectroscopy. J. Biomed. Opt. 2008, 13, 034017. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.L.; Hendrickson, D.A. Surgical procedures in pinniped and cetacean species. J. Zoo Wildl. Med. 2013, 44, 817–836. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.J.E.; Tzeng, Y.C.; Galvin, S.D.; Thomas, K.N.; Ogoh, S.; Ainslie, P.N. Influence of changes in blood pressure on cerebral perfusion and oxygenation. Hypertension 2010, 55, 698–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, M.; Subudhi, A.W.; Roach, R.C. Effect of ventilation on cerebral oxygenation during exercise: Insights from canonical correlation. Respir. Physiol. Neurobiol. 2009, 166, 125–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuypers, J.; Matakas, F.; Potolicchio, S.J. Effect of central venous pressure on brain tissue pressure and brain volume. J. Neurosurg. 1976, 45, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Brazy, J.E. Effects of crying on cerebral blood volume and cytochrome aa3. J. Pediatr. 1988, 112, 457–461. [Google Scholar] [CrossRef]
- Todd, M.M.; Weeks, J.B.; Warner, D.S. The influence of intravascular volume expansion on cerebral blood flow and blood volume in normal rats. Anesthesiology 1993, 78, 945–953. [Google Scholar] [CrossRef]
- Tsai, F.Y.; Kostanian, V.; Rivera, M.; Lee, K.W.; Chen, C.C.; Nguyen, T.H. Cerebral venous congestion as indication for thrombolytic treatment. Cardiovasc. Intervent. Radiol. 2007, 30, 675–687. [Google Scholar] [CrossRef]
- Hamilton, W.F.; Woodbury, R.A.; Harper, H.T. Physiologic relationships between intrathoracic, intraspinal and arterial pressures. J. Am. Med. Assoc. 1936, 107, 853–856. [Google Scholar] [CrossRef]
- Haykowsky, M.J.; Eves, N.D.; Warburton, D.E.R.; Findlay, M.J. Resistance exercise, the Valsalva Maneuver, and cerebrovascular transmural pressure. Med. Sci. Sport. Exerc. 2003, 35, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Phan, P.; Highton, D.; Lai, J.; Smith, M.; Elwell, C.; Tachtsidis, I. Multi-channel multi-distance broadband near-infrared spectroscopy system to measure the spatial response of cellular oxygen metabolism and tissue oxygenation. Biomed. Opt. Express 2016, 7, 4424–4440. [Google Scholar] [CrossRef] [PubMed]
- Dowdy, E.G.; Kaya, K. Studies of the mechanism of cardiovascular responses to CI-581. Anesthesiology 1968, 29, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Waterman, A.; Livingston, A. Some physiological effects of ketamine in sheep. Res. Vet. Sci. 1978, 25, 225–233. [Google Scholar] [CrossRef]
- Chang, P. The effects of ketamine on guinea pig heart. Br. J. Anaesth. 1973, 45, 929–930. [Google Scholar] [CrossRef]
- Morse, Z.; Sano, K.; Kanri, T. Effects of a midazolam-ketamine admixture in human volunteers. Anesth. Prog. 2004, 51, 76–79. [Google Scholar]
- Andrews, R.D.; Jones, D.R.; Williams, J.D.; Thorson, P.H.; Oliver, G.W.; Costa, D.P.; Le Boeuf, B.J. Heart rates of northern elephant seals diving at sea and resting on the beach. J. Exp. Biol. 1997, 200, 2083–2095. [Google Scholar] [CrossRef]
- Thompson, D.; Fedak, M.A. Cardiac responses of grey seals during diving at sea. J. Exp. Biol. 1993, 174, 139–164. [Google Scholar] [CrossRef]
- Field, I.C.; Bradshaw, C.J.A.; McMahon, C.R.; Harrington, J.; Burton, H.R. Effects of age, size and condition of elephant seals (Mirounga leonina) on their intravenous anaesthesia with tiletamine and zolazepam. Vet. Rec. 2002, 151, 235–240. [Google Scholar] [CrossRef]
- McMahon, C.R.; Burton, H.; McLean, S.; Slip, D.; Bester, M. Field immobilization of southern elephant seals with intravenous tiletamine and zolazepam. Vet. Rec. 2000, 146, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Mellish, J.-A.E.; Tuomi, P.A.; Hindle, A.G.; Horning, M. Chemical immobilization of Weddell seals (Leptonychotes weddellii) by ketamine/midazolam combination. Vet. Anaesth. Analg. 2010, 37, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Cohen, W.S.; Roberts, W.N.; Levenson, J.L. Benzodiazepines, other anxiolytic agents, and sedative hypnotics. In Systemic Lupus Erythematosus; Lahita, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 785–825. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Variable Type | Dependent Variable | Drug |
|---|---|---|---|
| Time series starting at 60 s pre-drugging to 180 s post-drugging | Smooth | [ΔO2Hb], [ΔHHb], heart rate, and SpO2 | Ketamine and midazolam |
| Time since initial drugging (s) | Smooth | [ΔO2Hb], [ΔHHb], heart rate, and SpO2 | Ketamine and midazolam |
| Cumulative bolus volume of ketamine and midazolam (mL) | Smooth | [ΔO2Hb], [ΔHHb], heart rate, and SpO2 | Ketamine and midazolam |
| Respiratory band (mV) | Smooth | [ΔO2Hb], [ΔHHb], and heart rate | Ketamine and midazolam |
| Ketamine bolus volume (mL) | Smooth | [ΔO2Hb], [ΔHHb], heart rate, and SpO2 | Ketamine |
| Midazolam bolus volume (mL) | Factor | [ΔO2Hb], [ΔHHb], heart rate, and SpO2 | Midazolam |
| Side (left or right hemisphere) | Factor | [ΔO2Hb] and [ΔHHb] | Ketamine and midazolam |
| Animal ID | Random effect | [ΔO2Hb], [ΔHHb], heart rate, and SpO2 | Ketamine and midazolam |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bønnelycke, E.-M.S.; Hastie, G.D.; Bennett, K.A.; Kainerstorfer, J.M.; Milne, R.; Moss, S.E.W.; Ruesch, A.; Wu, J.; McKnight, J.C. Wearable Near-Infrared Spectroscopy as a Physiological Monitoring Tool for Seals under Anaesthesia. Remote Sens. 2021, 13, 3553. https://doi.org/10.3390/rs13183553
Bønnelycke E-MS, Hastie GD, Bennett KA, Kainerstorfer JM, Milne R, Moss SEW, Ruesch A, Wu J, McKnight JC. Wearable Near-Infrared Spectroscopy as a Physiological Monitoring Tool for Seals under Anaesthesia. Remote Sensing. 2021; 13(18):3553. https://doi.org/10.3390/rs13183553
Chicago/Turabian StyleBønnelycke, Eva-Maria S., Gordon D. Hastie, Kimberley A. Bennett, Jana M. Kainerstorfer, Ryan Milne, Simon E. W. Moss, Alexander Ruesch, Jingyi Wu, and J. Chris McKnight. 2021. "Wearable Near-Infrared Spectroscopy as a Physiological Monitoring Tool for Seals under Anaesthesia" Remote Sensing 13, no. 18: 3553. https://doi.org/10.3390/rs13183553