Analyzing Ecological Vulnerability and Vegetation Phenology Response Using NDVI Time Series Data and the BFAST Algorithm

Abstract
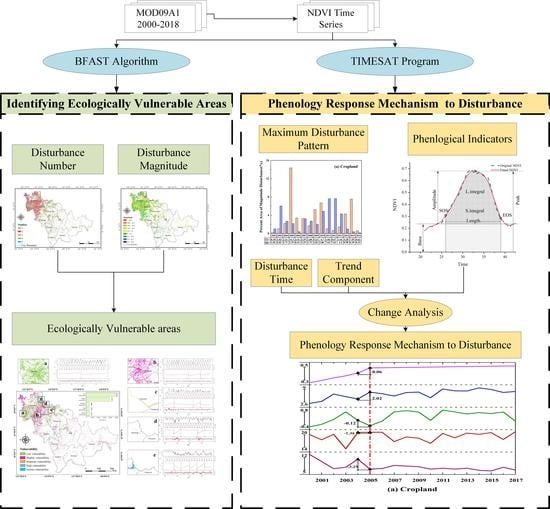
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Indexes Calculating and LULC Map
2.3. Ecologically Vulnerable Areas and Phenological Responses Methods
2.3.1. Identifying Ecologically Vulnerable Areas
2.3.2. BFAST Result Verification
2.3.3. Exploring the Phenological Response Mechanism to Negative Changes
- (1)
- The number of seasons and their approximate years were identified. For all vegetation types in the study area, one year was taken as a growing season. Every interval of 20 data points was considered a growth cycle, and 19 growth phases were counted between 2000 and 2018.
- (2)
- The best fitting model was selected according to the characteristics of the time-series trajectory. In TIMESAT, there are three fitting models: double logistic, asymmetric Gaussian, and SG filtering. The SG method was chosen, with which subtle and rapid changes in the simulation of local variations can be captured [52]. The expression for SG filtering is as follows:where is the reconstructed time-series NDVI data, Ci is the filtering coefficient, Yj is the original NDVI data, N is the number of data points in the sliding window (N = 2m + 1), and 2m + 1 is the filtering window width. Two parameters are required in SG filtering: the filter window width and the order of the polynomial for the smooth fitting process. The filter coefficients of the SG filter were determined by an unweighted linear least-squares regression and a second-order polynomial model. In this paper, the size of the filter window was set to 5, and the order of the fitting polynomial was taken as 2.
- (3)
- The time series trajectory characteristics of NDVI were extracted by BFAST and TIMESAT. The difference is that BFAST detects the inter-annual variation characteristics of the long-term time series, while TIMESAT detects the local variation characteristics of the NDVI time series (that is, the seasonal changes). The phenological indicators extracted by TIMESAT were divided into 3 categories according to their mean growth phase: start of season (SOS) and end of season (EOS). The maximum during the growth phase is taken as the peak and amplitude, and the integral during the growth phase includes the large integral (the L. integral) and small integral (the S. integral). The S. integral is equal to the L. integral minus the integral of the base line from SOS to EOS. In this study, the SOS and EOS were determined using a fixed threshold approach, with which the smoothed 8-day NDVI reached 25% of the mean amplitude for each growth phase for each pixel. The amplitude was calculated as the peak minus the minimum of the smoothed NDVI values. The minimum of the smoothed NDVI values was set to zero if the smoothed NDVI values were negative. The length of season (LOS) was calculated as the EOS minus SOS. The position of each indicator in the time-series trajectory is shown in Figure 4.
3. Results
3.1. Identification of Ecologically Vulnerable Areas by BFAST
3.2. BFAST Result Verification
3.3. Analysis of the Vulnerability of Different Vegetation Types
3.4. Vegetation Phenological Responses to Negative Changes
4. Discussion
4.1. Spatial Distribution of the Ecologically Vulnerable Area
4.2. The Response of Vegetation Phenology to Negative Changes
4.3. Limitations and Future Work
5. Conclusions
- (1)
- In this paper, an ecologically fragile zone identification framework based on the breakpoint detection of the BFAST NDVI time series was proposed. By identifying ecologically vulnerable areas to detect the number of negative changes and the magnitude of negative changes in vegetation growth over many years, we fully considered the long-term stability of vegetation growth and sensitivity to specific disturbances. This method can accurately reflect the long-term and short-term changes in vegetation growth and has better applicability in semi-arid regions.
- (2)
- During the past 19 years, northwest Jilin Province located in the semi-arid area was identified as ecologically vulnerable, primarily with low and slight vulnerability, where rainfall is scarce. Moreover, artificial irrigation cannot meet the needs of vegetation growth. Moisture is the main limiting factor leading to the vulnerability of the region’s ecological environment.
- (3)
- Compared to other vegetation types with dense coverage, 60% of the area had a number of negative changes greater than 1, a much larger area than that of other vegetation types (less than 50%). The magnitude of change in sparse vegetation was −0.429, which is the lowest among the vegetation types. This shows that sparse vegetation is more susceptible to drought. Therefore, increasing the vegetation coverage or changing to more stable vegetation types can reduce the fragility in ecologically vulnerable areas.
- (4)
- For vegetation types with dense coverage, the impact of negative changes on vegetation phenology shows long-term effects. The negative changes in the NDVI trends of various types of vegetation led to a fluctuation range of the integral value from −0.06 to −6.9 and a phenological period length from −0.3 to −5.4; the peak value varied from −0.11 to 0.12. Negative change had a significant effect on the cumulative values of the growth phase, such as the relative amount of vegetation biomass and the length of the growing period, but less of an effect on the instantaneous value of the peak. Detecting changes in the growth phase or the integral value could be used to predict whether the vegetation growth experiences a negative change.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feehan, J.; Harley, M.; Minnen, J. Climate change in Europe. Impact on terrestrial ecosystems and biodiversity. A review. Agron. Sustain. Dev. 2009, 29, 409–421. [Google Scholar] [CrossRef]
- Shriver, R.K.; Andrews, C.M.; Arkle, R.S.; Barnard, D.M.; Duniway, M.C.; Germino, M.J.; Pilliod, D.S.; Pyke, D.A.; Welty, J.L.; Bradford, J.B. Transient population dynamics impede restoration and may promote ecosystem transformation after disturbance. Ecol. Lett. 2019, 22, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- E Fernández-Giménez, M.; Allington, G.; Angerer, J.; Reid, R.S.; Jamsranjav, C.; Ulambayar, T.; Hondula, K.; Baival, B.; Batbuyan, B.; Altanzul, T.; et al. Using an integrated social-ecological analysis to detect effects of household herding practices on indicators of rangeland resilience in Mongolia. Environ. Res. Lett. 2018, 13, 075010. [Google Scholar] [CrossRef]
- Reid, R.S.; Fernández-Giménez, M.E.; Galvin, K.A. Dynamics and Resilience of Rangelands and Pastoral Peoples Around the Globe. Annu. Rev. Environ. Resour. 2014, 39, 217–242. [Google Scholar] [CrossRef]
- Turner, B.L.; Kasperson, R.E.; Matson, P.A.; McCarthy, J.J.; Corell, R.W.; Christensen, L.; Eckley, N.; Kasperson, J.X.; Luers, A.; Martello, M.L.; et al. A framework for vulnerability analysis in sustainability science. Proc. Natl. Acad Sci. USA 2003, 100, 8074–8079. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.; Jiang, R.; Yang, C.; Zhang, F.; Su, M.; Liao, Q. Establishing an ecological vulnerability assessment indicator system for spatial recognition and management of ecologically vulnerable areas in highly urbanized regions: A case study of Shenzhen, China. Ecol. Indic. 2016, 69, 540–547. [Google Scholar] [CrossRef]
- Zhumanova, M.; Mönnig, C.; Hergarten, C.; Darr, D.; Wrage-Mönnig, N. Assessment of vegetation degradation in mountainous pastures of the Western Tien-Shan, Kyrgyzstan, using eMODIS NDVI. Ecol. Indic. 2018, 95, 527–543. [Google Scholar] [CrossRef]
- Chen, X.; Wang, W.; Chen, J.; Zhu, X.; Shen, M.; Gan, L.; Cao, X. Does any phenological event defined by remote sensing deserve particular attention? An examination of spring phenology of winter wheat in Northern China. Ecol. Indic. 2020, 116, 106456. [Google Scholar] [CrossRef]
- Yuan, F. Land-cover change and environmental impact analysis in the Greater Mankato area of Minnesota using remote sensing and GIS modelling. Int. J. Remote. Sens. 2007, 29, 1169–1184. [Google Scholar] [CrossRef]
- Fraga, H.; Amraoui, M.; Malheiro, A.C.; Moutinho-Pereira, J.; Eiras-Dias, J.; Silvestre, J.; Santos, J.A. Examining the relationship between the Enhanced Vegetation Index and grapevine phenology. Eur. J. Remote. Sens. 2014, 47, 753–771. [Google Scholar] [CrossRef]
- Pei, H.; Fang, S.; Lin, L.; Qin, Z.; Wang, X. Methods and applications for ecological vulnerability evaluation in a hyper-arid oasis: A case study of the Turpan Oasis, China. Environ. Earth Sci. 2015, 74, 1449–1461. [Google Scholar] [CrossRef] [Green Version]
- Lewontin, R.C. The meaning of stability. In Brookhaven Symposia in Biology; Brookhaven National Laboratory: Upton, NY, USA, 1969; Volume 22, pp. 13–24. [Google Scholar]
- Romer, H.; Willroth, P.; Kaiser, G.; Vafeidis, A.T.; Ludwig, R.; Sterr, H.; Diez, J.R. Potential of remote sensing techniques for tsunami hazard and vulnerability analysis – a case study from Phang-Nga province, Thailand. Nat. Hazards Earth Syst. Sci. 2012, 12, 2103–2126. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.S.; Kolden, C.A.; Tinkham, W.T.; Talhelm, A.F.; Marshall, J.D.; Hudak, A.T.; Boschetti, L.; Falkowski, M.J.; Greenberg, J.A.; Anderson, J.W.; et al. Remote sensing the vulnerability of vegetation in natural terrestrial ecosystems. Remote. Sens. Environ. 2014, 154, 322–337. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Carpenter, S.R.; Ives, A.R.; Kucharik, C.J.; Ramiadantsoa, T.; Stegner, M.A.; Williams, J.W.; Zhang, J.; Turner, M.G. Abrupt Change in Ecological Systems: Inference and Diagnosis. Trends Ecol. Evol. 2018, 33, 513–526. [Google Scholar] [CrossRef]
- Bulletin of the People’s Republic of China Home Page. Available online: http://www.gov.cn/gongbao/content/2009/content_1250928.htm (accessed on 14 October 2020).
- Jiang, L.; Huang, X.; Wang, F.; Liu, Y.; An, P. Method for evaluating ecological vulnerability under climate change based on remote sensing: A case study. Ecol. Indic. 2018, 85, 479–486. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Sankey, J.B.; Wallace, C.S.; Ravi, S. Phenology-based, remote sensing of post-burn disturbance windows in rangelands. Ecol. Indic. 2013, 30, 35–44. [Google Scholar] [CrossRef]
- Ma, X.; Huete, A.; Moran, S.; Ponce-Campos, G.; Eamus, D. Abrupt shifts in phenology and vegetation productivity under climate extremes. J. Geophys. Res. Biogeosciences 2015, 120, 2036–2052. [Google Scholar] [CrossRef]
- Minin, A.A.; Trofimov, I.E.; Zakharov, V.M. Assessment of the Stability of Phenological Indices of the Silver Birch Betula pendula under Climate Change. Biol. Bull. 2020, 47, 149–152. [Google Scholar] [CrossRef]
- Verbesselt, J.; Zeileis, A.; Herold, M. Near real-time disturbance detection using satellite image time series. Remote. Sens. Environ. 2012, 123, 98–108. [Google Scholar] [CrossRef]
- Jones, M.O.; Kimball, J.S.; Jones, L.A. Satellite microwave detection of boreal forest recovery from the extreme 2004 wildfires in Alaska and Canada. Glob. Chang. Biol. 2013, 19, 3111–3122. [Google Scholar] [CrossRef] [PubMed]
- Bullock, E.L.; Woodcock, C.E.; Olofsson, P. Monitoring tropical forest degradation using spectral unmixing and Landsat time series analysis. Remote. Sens. Environ. 2020, 238, 110968. [Google Scholar] [CrossRef]
- Wen, L.; Saintilan, N. Climate phase drives canopy condition in a large semi-arid floodplain forest. J. Environ. Manag. 2015, 159, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Hao, X.; Qu, J.J. Monitoring Extreme Agricultural Drought over the Horn of Africa (HOA) Using Remote Sensing Measurements. Remote. Sens. 2019, 11, 902. [Google Scholar] [CrossRef] [Green Version]
- Price, B.; Waser, L.T.; Wang, Z.; Marty, M.; Ginzler, C.; Zellweger, F. Predicting biomass dynamics at the national extent from digital aerial photogrammetry. Int. J. Appl. Earth Obs. Geoinformation 2020, 90, 102116. [Google Scholar] [CrossRef]
- DeChant, B.; Ryu, Y.; Badgley, G.; Zeng, Y.; Berry, J.A.; Zhang, Y.; Goulas, Y.; Li, Z.; Zhang, Q.; Kang, M.; et al. Canopy structure explains the relationship between photosynthesis and sun-induced chlorophyll fluorescence in crops. Remote. Sens. Environ. 2020, 241, 111733. [Google Scholar] [CrossRef] [Green Version]
- Malenovský, Z.; Mishra, K.B.; Zemek, F.; Rascher, U.; Nedbal, L. Scientific and technical challenges in remote sensing of plant canopy reflectance and fluorescence. J. Exp. Bot. 2009, 60, 2987–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, J.; Masek, J.G. The vegetation greenness trend in Canada and US Alaska from 1984–2012 Landsat data. Remote. Sens. Environ. 2016, 176, 1–16. [Google Scholar] [CrossRef]
- Qian, Y.; Yang, Z.; Di, L.; Rahman, S.; Tan, Z.; Xue, L.; Gao, F.; Yu, E.; Zhang, X. Crop Growth Condition Assessment at County Scale Based on Heat-Aligned Growth Stages. Remote. Sens. 2019, 11, 2439. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Ding, Y.; Shi, H.; Cai, H.; Fu, Q.; Liu, S.; Li, T. Analysis and prediction of vegetation dynamic changes in China: Past, present and future. Ecol. Indic. 2020, 117, 106642. [Google Scholar] [CrossRef]
- Watts, L.M.; Laffan, S.W. Effectiveness of the BFAST algorithm for detecting vegetation response patterns in a semi-arid region. Remote. Sens. Environ. 2014, 154, 234–245. [Google Scholar] [CrossRef]
- Chen, H.; Liang, Q.; Liang, Z.; Liu, Y.; Xie, S. Remote-sensing disturbance detection index to identify spatio-temporal varying flood impact on crop production. Agric. For. Meteorol. 2019, 180–191. [Google Scholar] [CrossRef]
- White, P.S.; Pickett, S.T.A. Chapter 1—Natural Disturbance and Patch Dynamics: An Introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: San Diego, CA, USA, 1985; pp. 3–13. [Google Scholar] [CrossRef]
- Yang, Y.; Erskine, P.D.; Lechner, A.M.; Mulligan, D.; Zhang, S.; Wang, Z. Detecting the dynamics of vegetation disturbance and recovery in surface mining area via Landsat imagery and LandTrendr algorithm. J. Clean. Prod. 2018, 178, 353–362. [Google Scholar] [CrossRef]
- Morrison, J.; Higginbottom, T.P.; Symeonakis, E.; Jones, M.J.; Omengo, F.; Walker, S.L.; Cain, B. Detecting Vegetation Change in Response to Confining Elephants in Forests Using MODIS Time-Series and BFAST. Remote. Sens. 2018, 10, 1075. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Huang, C.; Zhu, Z.; Wen, W.; Xu, D.; Liu, A. Use of remote sensing coupled with a vegetation change tracker model to assess rates of forest change and fragmentation in Mississippi, USA. Int. J. Remote. Sens. 2009, 30, 6559–6574. [Google Scholar] [CrossRef]
- Shen, X.-J.; An, R.; Feng, L.; Ye, N.; Zhu, L.; Li, M. Vegetation changes in the Three-River Headwaters Region of the Tibetan Plateau of China. Ecol. Indic. 2018, 93, 804–812. [Google Scholar] [CrossRef]
- Verbesselt, J.; Hyndman, R.; Newnham, G.; Culvenor, D. Detecting trend and seasonal changes in satellite image time series. Remote. Sens. Environ. 2010, 114, 106–115. [Google Scholar] [CrossRef]
- Fang, X.; Zhu, Q.; Ren, L.; Chen, H.; Wang, K.; Peng, C. Large-scale detection of vegetation dynamics and their potential drivers using MODIS images and BFAST: A case study in Quebec, Canada. Remote. Sens. Environ. 2018, 206, 391–402. [Google Scholar] [CrossRef]
- Schultz, M.; Clevers, J.G.; Carter, S.; Verbesselt, J.; Avitabile, V.; Quang, H.V.; Herold, M. Performance of vegetation indices from Landsat time series in deforestation monitoring. Int. J. Appl. Earth Obs. Geoinformation 2016, 52, 318–327. [Google Scholar] [CrossRef]
- Roerink, G.J.; Menenti, M.; Verhoef, W. Reconstructing cloudfree NDVI composites using Fourier analysis of time series. Int. J. Remote. Sens. 2000, 21, 1911–1917. [Google Scholar] [CrossRef]
- Colditz, R.R.; Conrad, C.; Wehrmann, T.; Schmidt, M.; Dech, S. TiSeG: A Flexible Software Tool for Time-Series Generation of MODIS Data Utilizing the Quality Assessment Science Data Set. IEEE Trans. Geosci. Remote. Sens. 2008, 46, 3296–3308. [Google Scholar] [CrossRef]
- McKellip, R.; Prados, D.; Ryan, R.; Ross, K.; Spruce, J.; Gasser, G.; Greer, R. Remote-Sensing Time Series Analysis, a Vegetation Monitoring Tool. Front. Environ. Sci. 2008. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, P.; Eklundh, L. TIMESAT—a program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Stanimirova, R.; Cai, Z.; Melaas, E.; Gray, J.M.; Eklundh, L.; Jönsson, P.; Friedl, M.A. Gray An Empirical Assessment of the MODIS Land Cover Dynamics and TIMESAT Land Surface Phenology Algorithms. Remote. Sens. 2019, 11, 2201. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wu, J.; Li, X.; Zhou, H.; Yang, J.; Geng, G.; An, X.; Liu, L.; Tang, Z. A comprehensively quantitative method of evaluating the impact of drought on crop yield using daily multi-scale SPEI and crop growth process model. Int. J. Biometeorol. 2016, 61, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Hoyos, A.; Rembold, F.; Kerdiles, H.; Gallego, J. Comparison of Global Land Cover Datasets for Cropland Monitoring. Remote. Sens. 2017, 9, 1118. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote. Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Walker, J.J.; Soulard, C.E. Phenology Patterns Indicate Recovery Trajectories of Ponderosa Pine Forests After High-Severity Fires. Remote. Sens. 2019, 11, 2782. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Brandt, M.; Liu, Y.Y.; Verger, A.; Tagesson, T.; Diouf, A.A.; Rasmussen, K.; Mbow, C.; Wang, Y.; Fensholt, R. Remote sensing of vegetation dynamics in drylands: Evaluating vegetation optical depth (VOD) using AVHRR NDVI and in situ green biomass data over West African Sahel. Remote. Sens. Environ. 2016, 177, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, R.; Tong, Z.; Liu, X.; Zhang, J. Dynamic Evaluation and Regionalization of Maize Drought Vulnerability in the Midwest of Jilin Province. Sustainability 2019, 11, 4234. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Negative Changes | Vulnerability |
|---|---|
| 1 | Low Vulnerability |
| 2 | Slight Vulnerability |
| 3 | Moderate Vulnerability |
| 4 | High Vulnerability |
| 5 | Serious Vulnerability |
| Vegetation Types | Number of Changes | Year of Change |
|---|---|---|
| Cropland | 1 | 2011 |
| Herbaceous | 1 | 2012 |
| Mosaic Natural Vegetation | 1 | 2012 |
| Tree Cover | 1 | 2009 |
| Shrubland | 1 | 2014 |
| Grassland | 1 | 2010 |
| Sparse Vegetation | 1 | 2011 |
| Vegetation Types | Growth Phase | Maximum | Integral | |||
|---|---|---|---|---|---|---|
| SOS | EOS | Amplitude | Peak | L. integral | S. integral | |
| Cropland | −0.73 | 0.16 | 0.61 | 0.83 | 0.89 | 0.74 |
| Herbaceous | −0.43 | 0.54 | −0.15 | 0.18 | 0.75 | 0.68 |
| Mosaic Natural Vegetation | 0.68 | 0.65 | 0.76 | 0.86 | 0.95 | 0.83 |
| Tree Cover | −0.27 | 0.46 | 0.42 | 0.78 | 0.79 | 0.57 |
| Shrubland | −0.5 | 0.58 | 0.71 | 0.74 | 0.88 | 0.74 |
| Grassland | −0.28 | −0.27 | 0.69 | 0.92 | 0.81 | 0.70 |
| Sparse Vegetation | 0.29 | 0.71 | 0.15 | 0.65 | 0.67 | 0.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Zhang, C.; Guo, H.; Chen, W.; Yun, W.; Gao, L.; Wang, H. Analyzing Ecological Vulnerability and Vegetation Phenology Response Using NDVI Time Series Data and the BFAST Algorithm. Remote Sens. 2020, 12, 3371. https://doi.org/10.3390/rs12203371
Ma J, Zhang C, Guo H, Chen W, Yun W, Gao L, Wang H. Analyzing Ecological Vulnerability and Vegetation Phenology Response Using NDVI Time Series Data and the BFAST Algorithm. Remote Sensing. 2020; 12(20):3371. https://doi.org/10.3390/rs12203371
Chicago/Turabian StyleMa, Jiani, Chao Zhang, Hao Guo, Wanling Chen, Wenju Yun, Lulu Gao, and Huan Wang. 2020. "Analyzing Ecological Vulnerability and Vegetation Phenology Response Using NDVI Time Series Data and the BFAST Algorithm" Remote Sensing 12, no. 20: 3371. https://doi.org/10.3390/rs12203371