

Effects of Stand Density on Growth, Soil Water Content and Nutrients in Black Locust Plantations in the Semiarid Loess Hilly Region
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Characteristics
2.2. Plot Setting and Sampling Measurement
2.3. Calculation of Growth Indexes
2.4. Statistical Analysis
3. Results
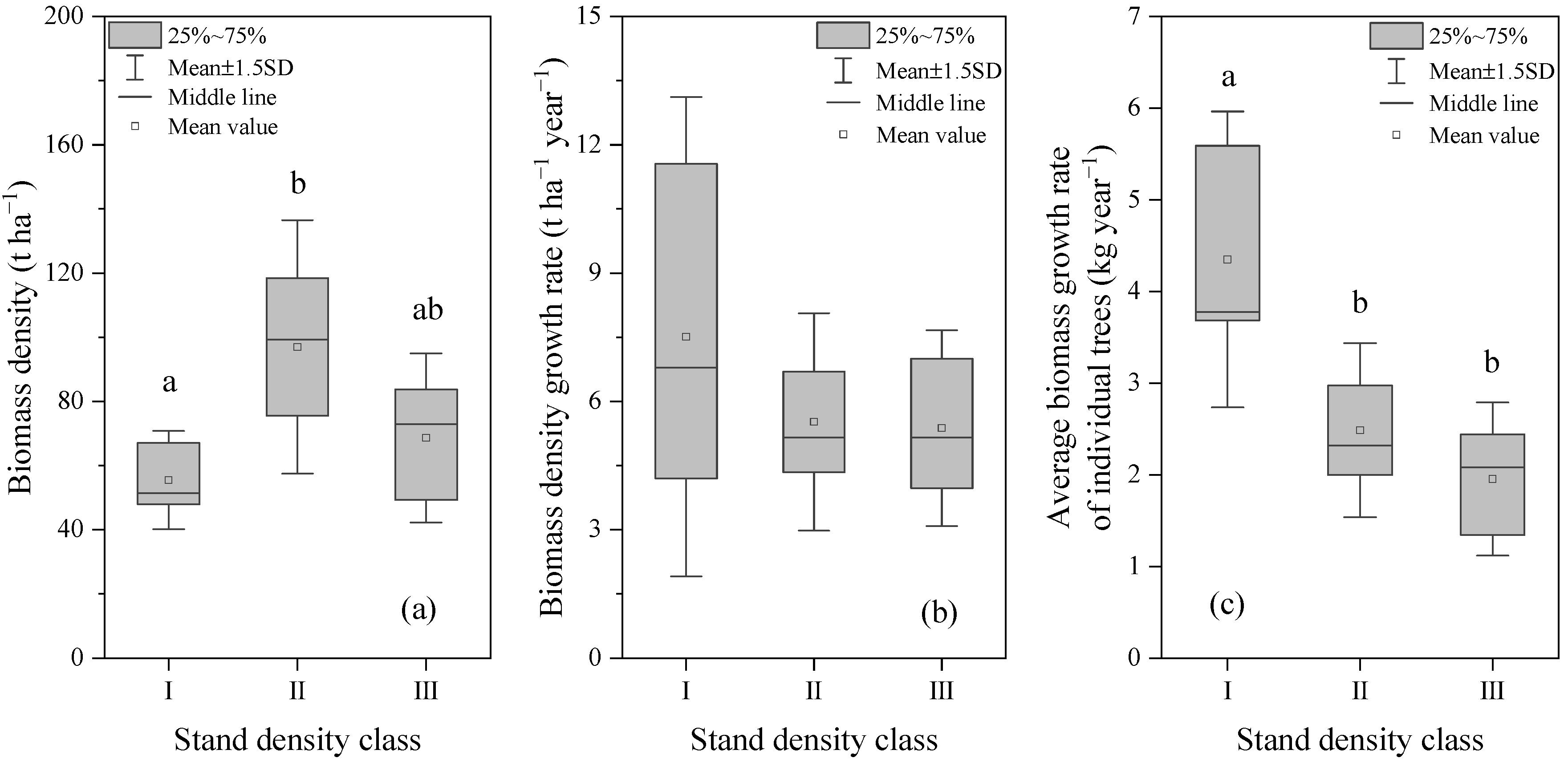
3.1. Variation in Stem Growth and Stand Biomass with Density Gradient
3.1.1. Variation in Stem Growth with Density Gradient
3.1.2. Variation in Stand Biomass with Density Gradient
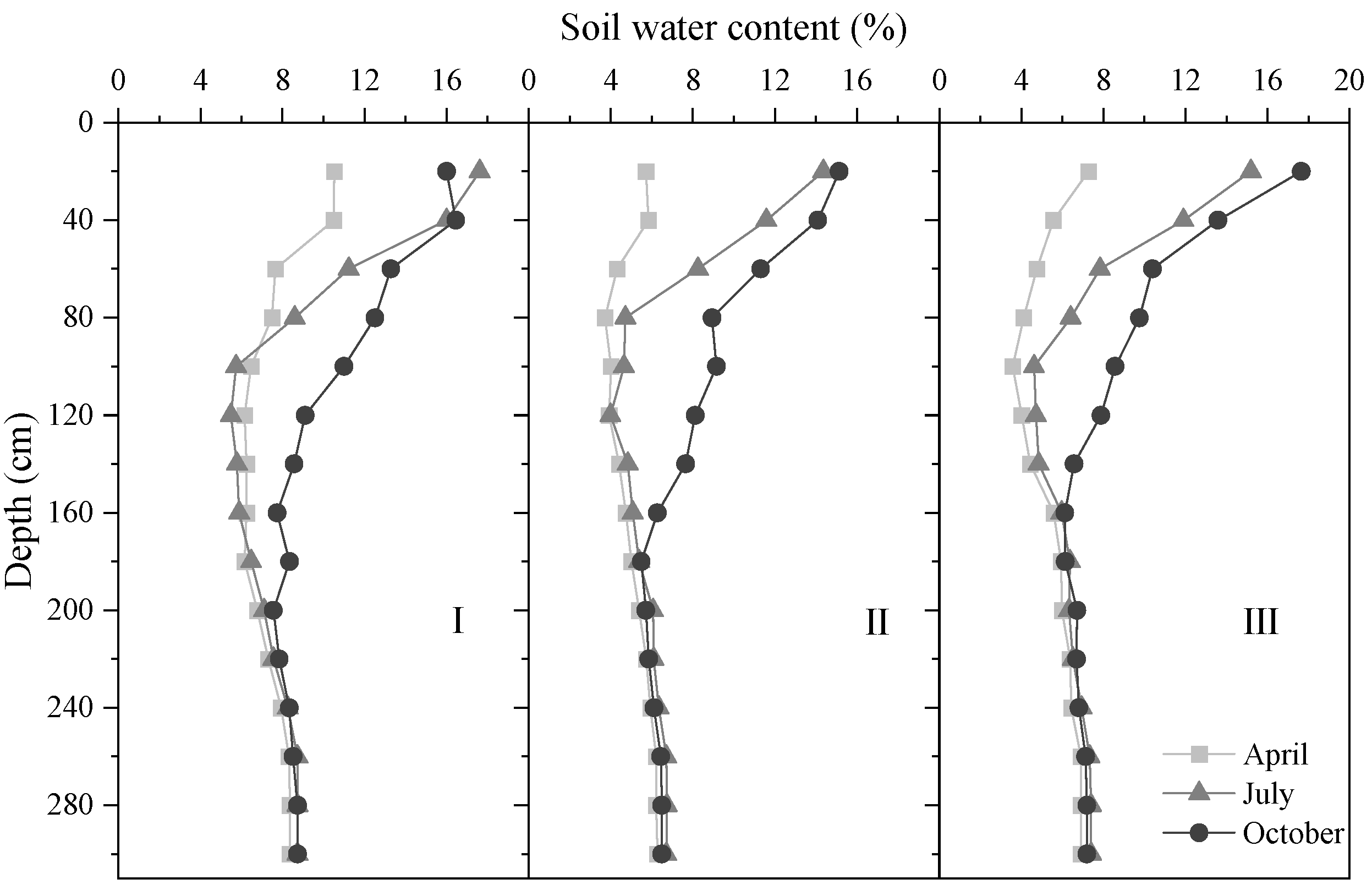
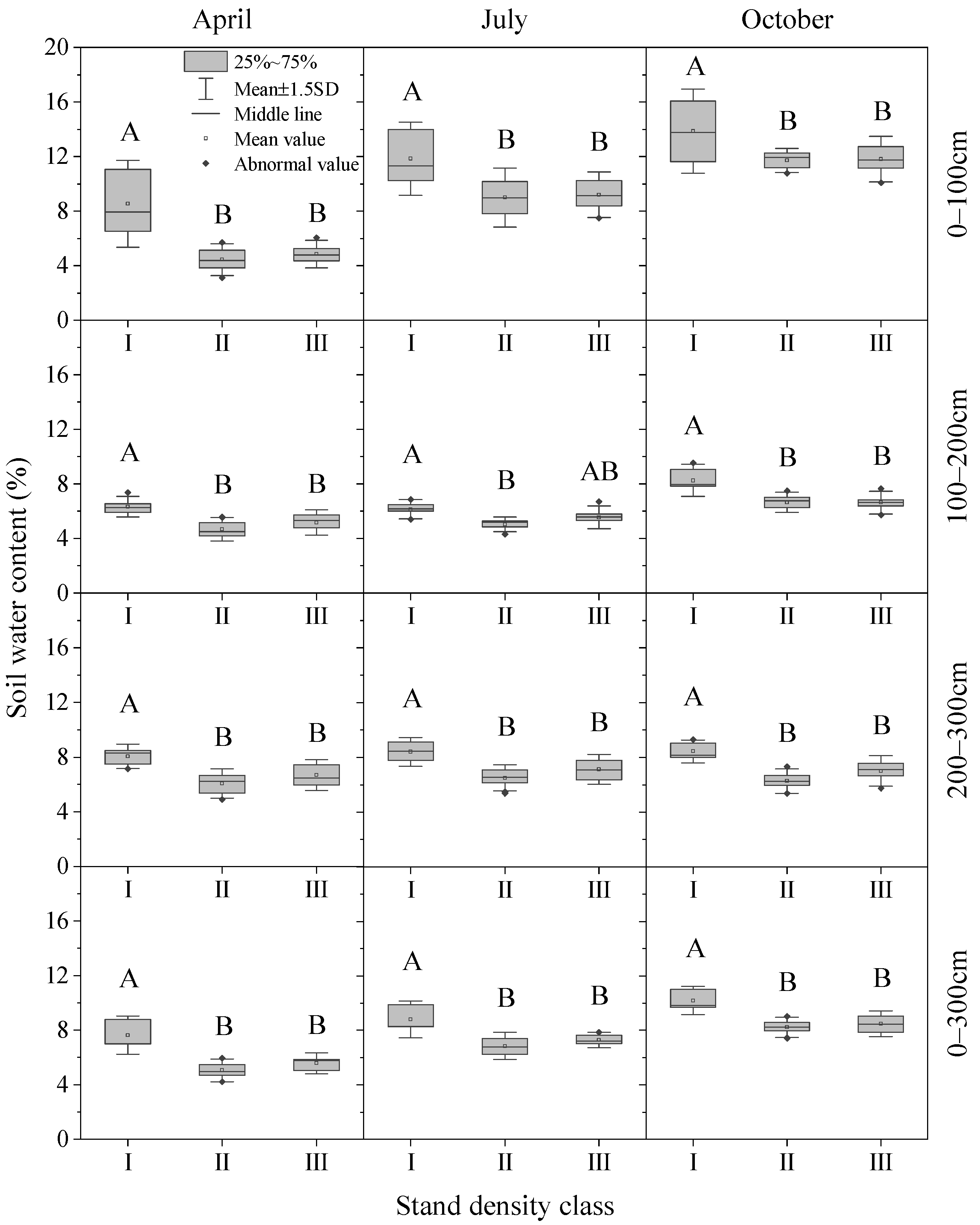
3.2. Effect of Stand Density on Spatio-Temporal Distribution of Soil Moisture
3.2.1. Variation in Vertical Distribution Dynamic of SWC with Density Gradient
3.2.2. Temporal Variations in SWC in Different Soil Layers and Density Classes
3.2.3. Monthly Variation Degree of SWC with Density Gradient
3.3. Effect of Stand Density on Soil Nutrient Condition
4. Discussion
4.1. Lower Density Is Beneficial to Stand Growth of the Plantations
4.2. Plantation with Lower Density Has Better Soil Moisture Conditions
4.3. Effect of Stand Density on Soil Nutrients in These Plantations Was Not Significant in the Present Growth Years
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.K.; Ni, J.; Yang, Q.K.; Li, R. Human impacts on soil erosion identified using land-use changes: A case study from the Loess Plateau, China. Phys. Geogr. 2006, 27, 109–126. [Google Scholar] [CrossRef]
- He, X.B.; Zhou, J.; Zhang, X.B.; Tang, K.L. Soil erosion response to climatic change and human activity during the Quaternary on the Loess Plateau, China. Reg. Environ. Chang. 2006, 6, 62–70. [Google Scholar] [CrossRef]
- Liu, J.G.; Li, S.X.; Ouyang, Z.Y.; Tam, C.; Chen, X.D. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proc. Natl. Acad. Sci. USA 2008, 105, 9477–9482. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.T.; Fu, B.J.; Liu, G.H.; Wang, Z. Hydrologic feasibility of artificial forestation in the semi-arid Loess Plateau of China. Hydrol. Earth Syst. Sci. 2011, 15, 2519–2530. [Google Scholar] [CrossRef]
- Deng, L.; Liu, G.B.; Shangguan, Z.P. Land-use conversion and changing soil carbon stocks in China’s ‘Grain-for-Green’ Program: A synthesis. Glob. Chang. Biol. 2014, 20, 3544–3556. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Q.; Xu, G.H.; Guo, K.; Du, S. Mapping the global potential geographical distribution of black locust (Robinia pseudoacacia L.) using herbarium data and a maximum entropy model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef]
- Zhang, Q.D.; Wei, W.; Chen, L.D.; Yang, L. The joint effects of precipitation gradient and afforestation on soil moisture across the Loess Plateau of China. Forests 2019, 10, 285. [Google Scholar] [CrossRef]
- Jiao, L.; An, W.M.; Li, Z.S.; Gao, G.Y.; Wang, C. Regional variation in soil water and vegetation characteristics in the Chinese Loess Plateau. Ecol. Indic. 2020, 115, 106399. [Google Scholar] [CrossRef]
- Liang, H.B.; Xue, Y.Y.; Li, Z.S.; Wang, S.; Wu, X.; Gao, G.Y.; Liu, G.H.; Fu, B.J. Soil moisture decline following the plantation of Robinia pseudoacacia forests: Evidence from the Loess Plateau. For. Ecol. Manag. 2018, 412, 62–69. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Fu, B.J.; Gao, G.Y.; Wang, S.; Jin, T.T.; Zhang, L.W.; Liu, J.B.; Zhang, D. Comparison of transpiration between different aged black locust (Robinia pseudoacacia) trees on the semi-arid Loess Plateau, China. J. Arid Land 2016, 8, 604–617. [Google Scholar] [CrossRef]
- Zhang, J.G.; Guan, J.H.; Shi, W.Y.; Yamanaka, N.; Du, S. Interannual variation in stand transpiration estimated by sap flow measurement in a semi-arid black locust plantation, Loess Plateau, China. Ecohydrology 2015, 8, 137–147. [Google Scholar] [CrossRef]
- Hou, G.R.; Bi, H.X.; Wei, X.; Wang, N.; Cui, Y.H.; Zhao, D.Y.; Ma, X.Z.; Wang, S.S. Optimal configuration of stand structures in a low-efficiency Robinia pseudoacacia forest based on a comprehensive index of soil and water conservation ecological benefits. Ecol. Indic. 2020, 114, 106308. [Google Scholar] [CrossRef]
- Zhao, X.K.; Li, Z.Y.; Zhu, D.H.; Zhu, Q.K.; Robeson, M.D.; Hu, J. Revegetation using the deep planting of container seedlings to overcome the limitations associated with topsoil desiccation on exposed steep earthy road slopes in the semiarid loess region of China. Land Degrad. Dev. 2018, 29, 2797–2807. [Google Scholar] [CrossRef]
- Wang, L.; Shao, M.A.; Hou, Q.C.; Yang, G.M. The analysis of dried soil layer of artificial Robinia pseudoacacia forestry land in the Yan’an experimental area. Acta Bot. Boreali-Occident. Sin. 2001, 21, 101–106. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yu, P.T.; Feger, K.H.; Wei, X.H.; Sun, G.; Bonell, M.; Xiong, W.; Zhang, S.L.; Xu, L.H. Annual runoff and evapotranspiration of forestlands and non-forestlands in selected basins of the Loess Plateau of China. Ecohydrology 2011, 4, 277–287. [Google Scholar] [CrossRef]
- Shao, M.A.; Wang, Y.Q.; Xia, Y.Q.; Jia, X.X. Soil drought and water carrying capacity for vegetation in the critical zone of the Loess Plateau: A review. Vadose Zone J. 2018, 17, 170077. [Google Scholar] [CrossRef]
- Jia, X.X.; Shao, M.A.; Zhu, Y.J.; Luo, Y. Soil moisture decline due to afforestation across the Loess Plateau, China. J. Hydrol. 2017, 546, 113–122. [Google Scholar] [CrossRef]
- Burkhart, H.E. Comparison of maximum size-density relationships based on alternate stand attributes for predicting tree numbers and stand growth. For. Ecol. Manag. 2013, 289, 404–408. [Google Scholar] [CrossRef]
- Moreno, G.; Cubera, E. Impact of stand density on water status and leaf gas exchange in Quercus ilex. For. Ecol. Manag. 2008, 254, 74–84. [Google Scholar] [CrossRef]
- Ji, L.B.; Shu, D.W. Influence of stand density of Cunninghamia lanceolate on stand growth. For. Inventory Plan. 2017, 42, 135–137. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Del Rio, M.; Heinrich, I.; Helle, G.; Canellas, I. Response of climate-growth relationships and water use efficiency to thinning in a Pinus nigra afforestation. For. Ecol. Manag. 2010, 259, 967–975. [Google Scholar] [CrossRef]
- Forrester, D.I.; Collopy, J.J.; Beadle, C.L.; Baker, T.G. Effect of thinning, pruning and nitrogen fertiliser application on light interception and light-use efficiency in a young Eucalyptus nitens plantation. For. Ecol. Manag. 2013, 288, 21–30. [Google Scholar] [CrossRef]
- Cai, H.Y.; Di, X.Y.; Chang, S.X.; Jin, G.Z. Stand density and species richness affect carbon storage and net primary productivity in early and late successional temperate forests differently. Ecol. Res. 2016, 31, 525–533. [Google Scholar] [CrossRef]
- Comita, L.S.; Queenborough, S.A.; Murphy, S.J.; Eck, J.L.; Xu, K.Y.; Krishnadas, M.; Beckman, N.; Zhu, Y. Testing predictions of the Janzen-Connell hypothesis: A meta-analysis of experimental evidence for distance- and density-dependent seed and seedling survival. J. Ecol. 2014, 102, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.H.; Shakoor, A.; Rashid, M.H.U.; Zhang, S.Y.; Wu, P.F.; Yan, W.D. Annual growth progression, nutrient transformation, and carbon storage in tissues of Cunninghamia lanceolata monoculture in relation to soil quality indicators influenced by intraspecific competition intensity. J. Soil Sci. Plant Nutr. 2021, 21, 3146–3158. [Google Scholar] [CrossRef]
- Lochhead, K.D.; Comeau, P.G. Relationships between forest structure, understorey light and regeneration in complex Douglas-fir dominated stands in south-eastern British Columbia. For. Ecol. Manag. 2012, 284, 12–22. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, J.J.; Zhou, H.; Zhao, Z. Photosynthetic carbon fixation capacity of black locust in rapid response to plantation thinning on the semiarid Loess Plateau in China. Pak. J. Bot. 2019, 51, 1365–1374. [Google Scholar] [CrossRef]
- Makinen, H.; Hein, S. Effect of wide spacing on increment and branch properties of young Norway spruce. Eur. J. For. Res. 2006, 125, 239–248. [Google Scholar] [CrossRef]
- Benomar, L.; DesRochers, A.; Larocque, G.R. The effects of spacing on growth, morphology and biomass production and allocation in two hybrid poplar clones growing in the boreal region of Canada. Trees-Struct. Funct. 2012, 26, 939–949. [Google Scholar] [CrossRef]
- Andrews, C.M.; D’Amato, A.W.; Fraver, S.; Palik, B.; Battaglia, M.A.; Bradford, J.B. Low stand density moderates growth declines during hot droughts in semi-arid forests. J. Appl. Ecol. 2020, 57, 1089–1102. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Q.; Gao, D.Q.; Zhang, B.B.; Zuo, H.J.; Jiang, J. Effects of thinning and understory removal on the soil water-holding capacity in Pinus massoniana plantations. Sci. Rep. 2021, 11, 13029. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Finley, K.A.; Johnson, N.G.; Ritchie, M.W. Lowering stand density enhances resiliency of ponderosa pine forests to disturbances and climate change. For. Sci. 2019, 65, 496–507. [Google Scholar] [CrossRef]
- Sanchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernandez-Cancio, A.; Vila-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Gavinet, J.; Ourcival, J.M.; Gauzere, J.; de Jalon, L.G.; Limousin, J.M. Drought mitigation by thinning: Benefits from the stem to the stand along 15 years of experimental rainfall exclusion in a holm oak coppice. For. Ecol. Manag. 2020, 473, 118266. [Google Scholar] [CrossRef]
- Tsamir, M.; Gottlieb, S.; Preisler, Y.; Rotenberg, E.; Tatarinov, F.; Yakir, D.; Tague, C.; Klein, T. Stand density effects on carbon and water fluxes in a semi-arid forest, from leaf to stand-scale. For. Ecol. Manag. 2019, 453, 117573. [Google Scholar] [CrossRef]
- Aldea, J.; Bravo, F.; Bravo-Oviedo, A.; Ruiz-Peinado, R.; Rodriguez, F.; del Rio, M. Thinning enhances the species-specific radial increment response to drought in Mediterranean pine-oak stands. Agric. For. Meteorol. 2017, 237, 371–383. [Google Scholar] [CrossRef]
- Simonin, K.; Kolb, T.E.; Montes-Helu, M.; Koch, G.W. The influence of thinning on components of stand water balance in a ponderosa pine forest stand during and after extreme drought. Agric. For. Meteorol. 2007, 143, 266–276. [Google Scholar] [CrossRef]
- Yang, X.; Li, T.C.; Shao, M. Factors controlling deep-profile soil organic carbon and water storage following afforestation of the Loess Plateau in China. For. Ecosyst. 2022, 9, 100079. [Google Scholar] [CrossRef]
- Wei, X.; Liang, W.J. Regulation of stand density alters forest structure and soil moisture during afforestation with Robinia pseudoacacia L. and Pinus tabulaeformis Carr. on the Loess Plateau. For. Ecol. Manag. 2021, 491, 119196. [Google Scholar] [CrossRef]
- Hu, Y.W.; Shi, Z.L.; Liu, C.; Xu, Q.T.; Zhang, J.J. Effects of stand densities on understory vegetation diversity and soil physicochemical properties of Robinia pseudoacacia forest in loess region of western Shanxi Province. Chin. J. Ecol. 2023, 42, 2072–2080. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, W.C.; Xu, M.P.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Lei, J.; Du, H.L.; Duan, A.G.; Zhang, J.G. Effect of stand density and soil layer on soil nutrients of a 37-year-old Cunninghamia lanceolata plantation in Naxi, Sichuan Province, China. Sustainability 2019, 11, 5410. [Google Scholar] [CrossRef]
- Jonsson, J.A.; Sigurdsson, B.D. Effects of early thinning and fertilization on soil temperature and soil respiration in a poplar plantation. Icel. Agric. Sci. 2010, 23, 97–109. [Google Scholar] [CrossRef]
- Ren, L.N.; Wang, H.Y.; Ding, G.D.; Gao, G.L.; Yang, X.J. Effects of Pinus tabulaeformis Carr. plantation density on soil organic carbon and nutrients characteristics in rocky mountain area of northern China. Arid Land Geogr. 2012, 35, 456–464. [Google Scholar] [CrossRef]
- He, Z.L.; Zhang, Y.X.; Guo, Y.D.; Yang, S.H.; Ren, D.; Ding, J.W.; Guo, J.P. Soil nutrient characteristics of natural Larix principis-rupprechtii in different stand density. Ecol. Environ. Sci. 2017, 26, 43–48. [Google Scholar] [CrossRef]
- Ma, C.K.; Luo, Y.; Shao, M.G. Comparative modeling of the effect of thinning on canopy interception loss in a semiarid black locust (Robinia pseudoacacia) plantation in northwest China. J. Hydrol. 2020, 590, 125234. [Google Scholar] [CrossRef]
- Tsunekawa, A.; Liu, G.B.; Yamanaka, N.; Du, S. Restoration and Development of the Degraded Loess Plateau, China; Springer: Tokyo, Japan, 2014; pp. 35–60. [Google Scholar]
- Cheng, R.R.; Chen, Q.W.; Zhang, J.G.; Shi, W.Y.; Li, G.Q.; Du, S. Soil moisture variations in response to precipitation in different vegetation types: A multi-year study in the loess hilly region in China. Ecohydrology 2020, 13, e2196. [Google Scholar] [CrossRef]
- Zheng, F.L. Effect of vegetation changes on soil erosion on the Loess Plateau. Pedosphere 2006, 16, 420–427. [Google Scholar] [CrossRef]
- State Forestry Administration. Regulations for Age-Class and Age-Group Division of Main Tree-Species. 2017, LY/T 2908-2017. Available online: http://lyj.baise.gov.cn/xxgk/zwgk/WJTZ/P020221114703370854900.pdf (accessed on 1 December 2023).
- Lyu, C.H.; Xu, Z.Y. Crop production changes and the impact of Grain for Green program in the Loess Plateau of China. J. Arid Land 2020, 12, 18–28. [Google Scholar] [CrossRef]
- Zhou, G.Y.; Yin, G.C.; Tang, X.L. Biomass Equation of Carbon Storage in China Forest Ecosystem; Science Press: Beijing, China, 2018. [Google Scholar]
- Paine, C.E.T.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: An update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Kerhoulas, L.P.; Kolb, T.E.; Koch, G.W. Tree size, stand density, and the source of water used across seasons by ponderosa pine in northern Arizona. For. Ecol. Manag. 2013, 289, 425–433. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rais, A. Wood quality in complex forests versus even-aged monocultures: Review and perspectives. Wood Sci. Technol. 2016, 50, 845–880. [Google Scholar] [CrossRef]
- Hakamada, R.E.; Hubbard, R.M.; Moreira, G.G.; Stape, J.L.; Campoe, O.; Ferraz, S.F.D. Influence of stand density on growth and water use efficiency in Eucalyptus clones. For. Ecol. Manag. 2020, 466, 118125. [Google Scholar] [CrossRef]
- Redei, K.; Meilby, H. Effect of thinning on the diameter increment in black locust (Robinia pseudoacacia L.) stands. Silva Gandav. 2000, 5, 63–74. [Google Scholar] [CrossRef]
- Wang, S.S. Directional Regulation of Robinia pseudoacacia Forest Density in the Loess Plateau in Western Shanxi, China; Beijing Forestry University: Beijing, China, 2021. [Google Scholar]
- Li, J.; Zhao, Z.; Yuan, Z.F.; Wang, D.H.; Hu, X.N. Dynamic model of soil moisture of Robinia pseudoacacia plantations in the Loess Plateau. Acta Bot. Boreali-Occident. Sin. 2014, 34, 1666–1675. [Google Scholar] [CrossRef]
- Ding, W.B.; Wang, F.; Jin, K. Effects of rainfall and plant characteristics on the spatiotemporal variation of soil moisture in a black locust plantation (Robinia pseudoacacia) on the Chinese Loess Plateau. Water 2023, 15, 1870. [Google Scholar] [CrossRef]
- Zhang, J.X.; Wang, X.; Wang, Y.K.; Jin, S.S. Regularities of rainfall infiltration and water migration in woodland drying soil in the loess hilly region. J. Soil Water Conserv. 2017, 31, 231–238. [Google Scholar] [CrossRef]
- Yang, L.; Wei, W.; Chen, L.D.; Chen, W.L.; Wang, J.L. Response of temporal variation of soil moisture to vegetation restoration in semi-arid Loess Plateau, China. CATENA 2014, 115, 123–133. [Google Scholar] [CrossRef]
- Bo, H.J.; Wen, C.Y.; Song, L.J.; Yue, Y.T.; Nie, L.S. Fine-root responses of populus tomentosa forests to stand density. Forests 2018, 9, 562. [Google Scholar] [CrossRef]
- Zheng, J.M.; Chen, X.Y.; Chen, L.G.; He, T.Y.; Rong, J.D.; Lin, Y.; Zheng, Y.S. Comprehensive evalution of soil quality at different stand densities of Dendrocalamus minor var. amoenus plantations. Appl. Ecol. Environ. Res. 2020, 18, 5985–5996. [Google Scholar] [CrossRef]
- Wei, X.; Bi, H.X.; Liang, W.J.; Hou, G.R.; Kong, L.X.; Zhou, Q.Z. Relationship between soil characteristics and stand structure of Robinia pseudoacacia L. and Pinus tabulaeformis Carr. mixed plantations in the Caijiachuan watershed: An application of structural equation modeling. Forests 2018, 9, 124. [Google Scholar] [CrossRef]
- Laganiere, J.; Angers, D.A.; Pare, D. Carbon accumulation in agricultural soils after afforestation: A meta-analysis. Glob. Chang. Biol. 2010, 16, 439–453. [Google Scholar] [CrossRef]
- Sun, M.M.; Zhai, B.C.; Chen, Q.W.; Li, G.Q.; Du, S. Response of density-related fine root production to soil and leaf traits in coniferous and broad-leaved plantations in the semiarid loess hilly region of China. J. For. Res. 2022, 33, 1071–1082. [Google Scholar] [CrossRef]
- Wang, Y.S.; Ma, B.M.; Gao, H.P.; Wang, B.T. Response of soil nutrients and their stoichiometric ratios to stand density in Pinus tabuliformis and Robinia pseudoacacia plantations in the loess region of western Shanxi Province, northern China. J. Beijing For. Univ. 2020, 42, 81–93. [Google Scholar] [CrossRef]
- Zhao, J.P. Effects of Mid-Aged Forest Density and Tending Years of Artificial Robinia pseudoacacia on Stand Composition Structure, Plant Functional Traits and Soil Properties; Northwest A&F University: Xianyang, China, 2022. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Density (Stems ha−1) | Stand Density Class | Altitude (m) | Age (Year) | Slope (°) | pH | Mean DBH (cm) | Mean Tree Height (m) | Soil Bulk Density (g cm−3) |
|---|---|---|---|---|---|---|---|---|
| 2130 | I | 1295 | 17 | 22 | 8.6 | 7.9 | 7.7 | 1.26 |
| 2315 | 1295 | 17 | 16 | 8.7 | 8.3 | 8.4 | 1.26 | |
| 2497 | 1290 | 17 | 14 | 8.7 | 7.3 | 6.5 | 1.26 | |
| 2745 | II | 1285 | 17 | 17 | 8.7 | 8.3 | 7.8 | 1.26 |
| 2825 | 1150 | 17 | 23 | 8.6 | 8.0 | 8.2 | 1.23 | |
| 3133 | 1150 | 17 | 21 | 8.7 | 9.1 | 8.5 | 1.23 | |
| 3478 | 1290 | 17 | 13 | 8.8 | 7.2 | 6.8 | 1.26 | |
| 3669 | III | 1150 | 17 | 21 | 8.6 | 6.2 | 6.2 | 1.23 |
| 3965 | 1285 | 17 | 23 | 8.7 | 7.1 | 7.3 | 1.26 | |
| 4026 | 1150 | 17 | 17 | 8.7 | 6.7 | 6.8 | 1.23 |
| Tree Organs | Biomass Equations |
|---|---|
| Roots | W = 0.0119(DBH2H)0.9501 |
| Leaves | W = 0.0060(DBH2H)0.8403 |
| Branches | W = 0.0040(DBH2H)1.0868 |
| Stem | W = 0.0302(DBH2H)0.9474 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, B.; Sun, M.; Shen, X.; Zhu, Y.; Li, G.; Du, S. Effects of Stand Density on Growth, Soil Water Content and Nutrients in Black Locust Plantations in the Semiarid Loess Hilly Region. Sustainability 2024, 16, 376. https://doi.org/10.3390/su16010376
Zhai B, Sun M, Shen X, Zhu Y, Li G, Du S. Effects of Stand Density on Growth, Soil Water Content and Nutrients in Black Locust Plantations in the Semiarid Loess Hilly Region. Sustainability. 2024; 16(1):376. https://doi.org/10.3390/su16010376
Chicago/Turabian StyleZhai, Bochao, Meimei Sun, Xiaojuan Shen, Yan Zhu, Guoqing Li, and Sheng Du. 2024. "Effects of Stand Density on Growth, Soil Water Content and Nutrients in Black Locust Plantations in the Semiarid Loess Hilly Region" Sustainability 16, no. 1: 376. https://doi.org/10.3390/su16010376