
Thermal Preference May Facilitate Spatial Coexistence of Two Invasive Fish Species in Lake Bosten, China
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
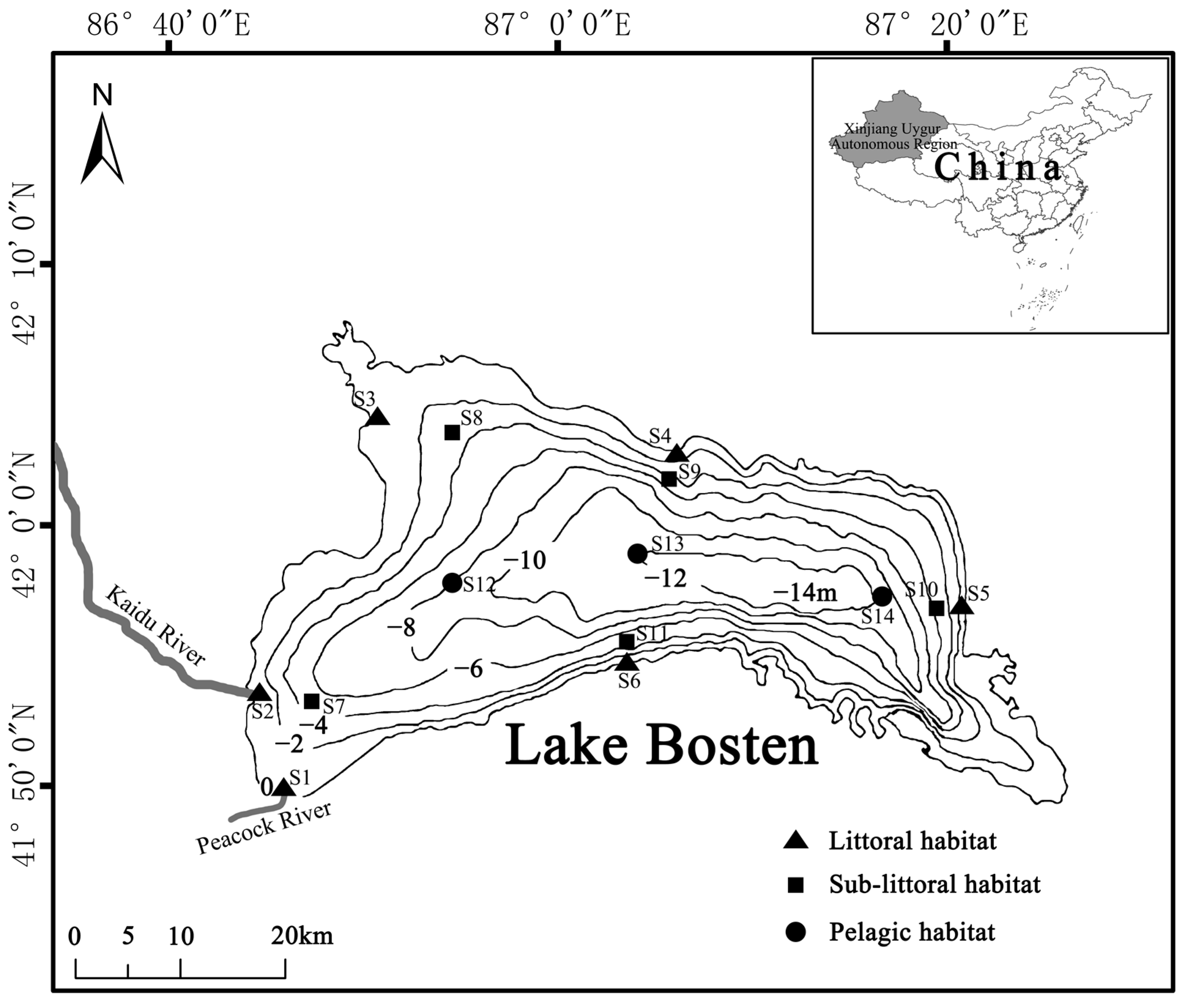
2.1. Study Area
2.2. Sampling Protocol
2.3. Statistical Analysis
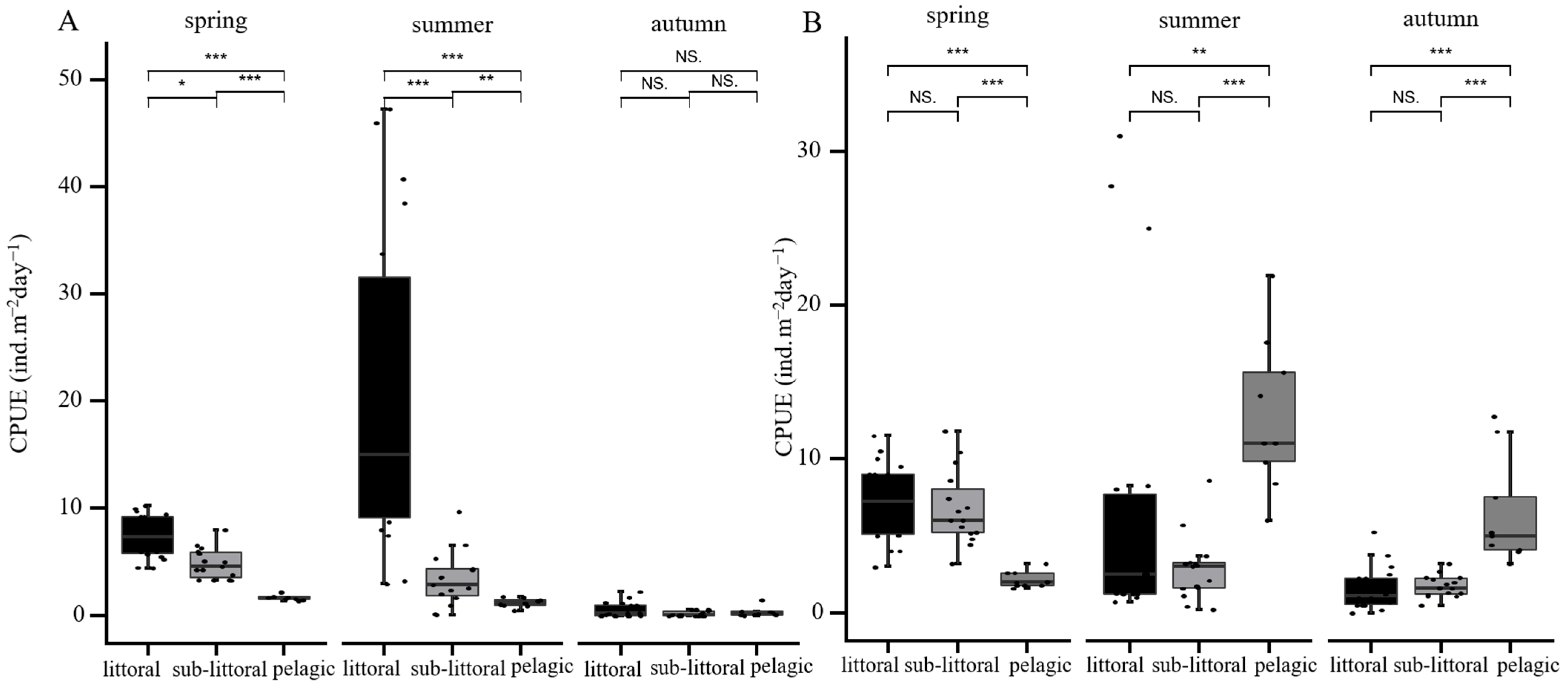
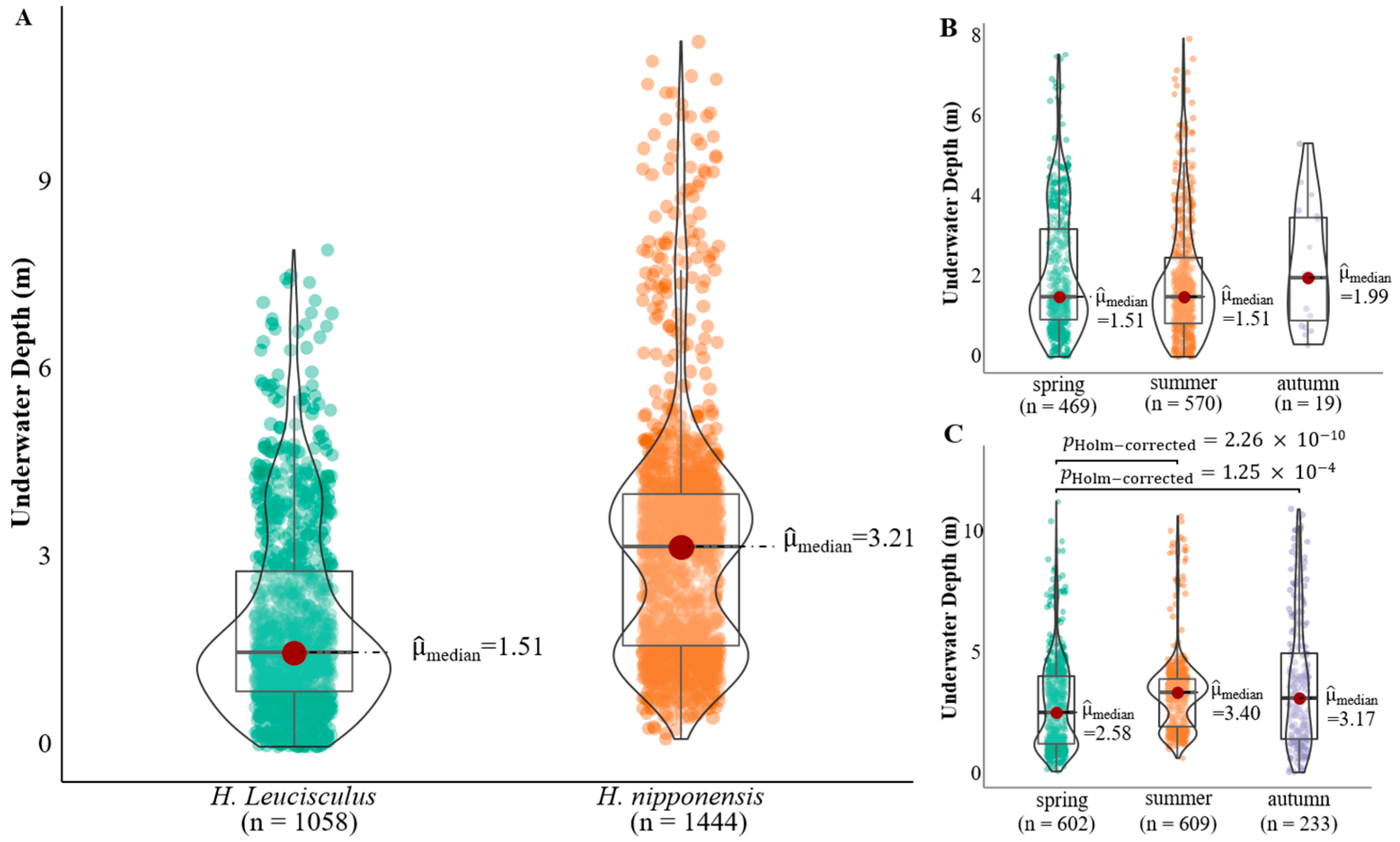
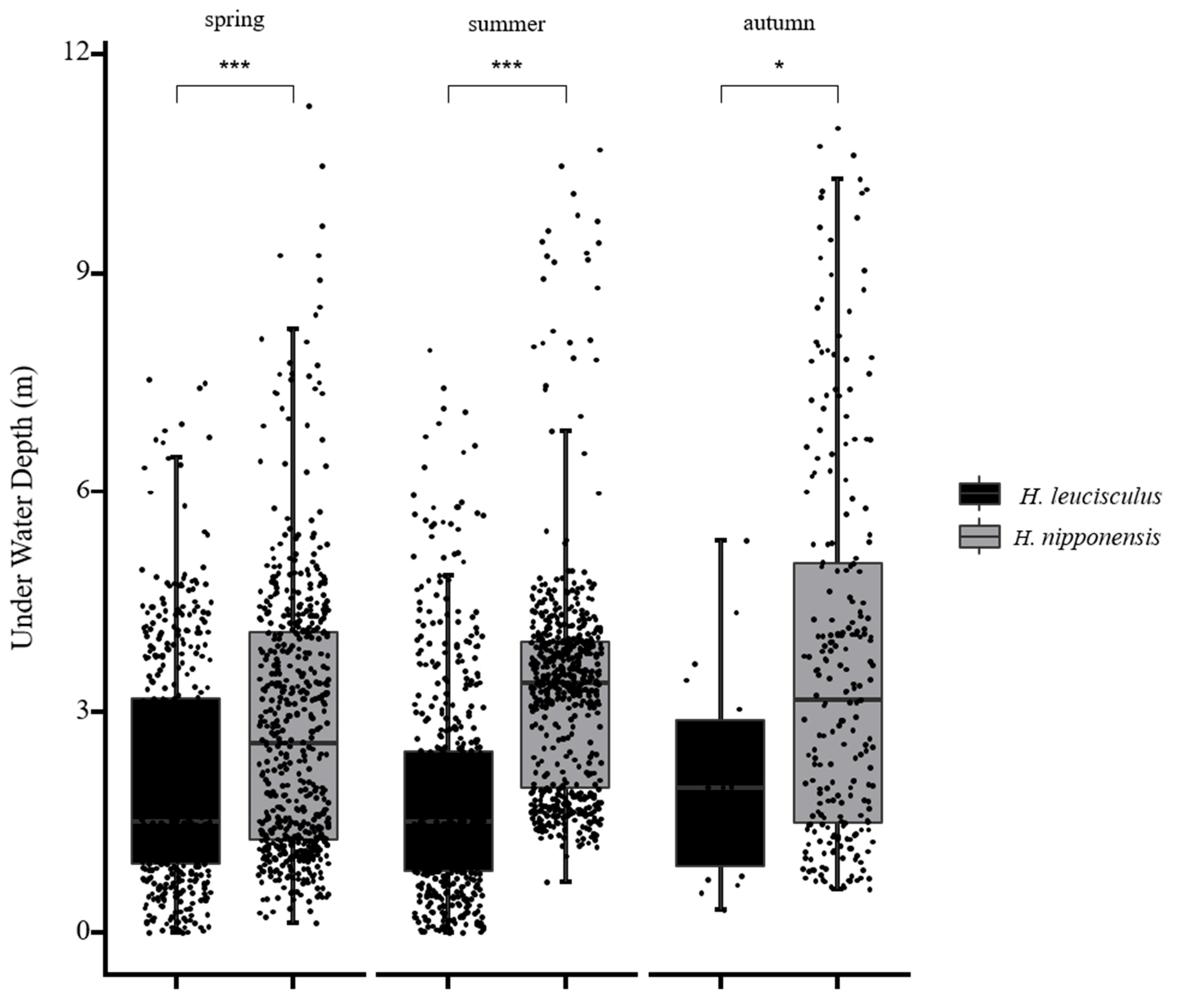
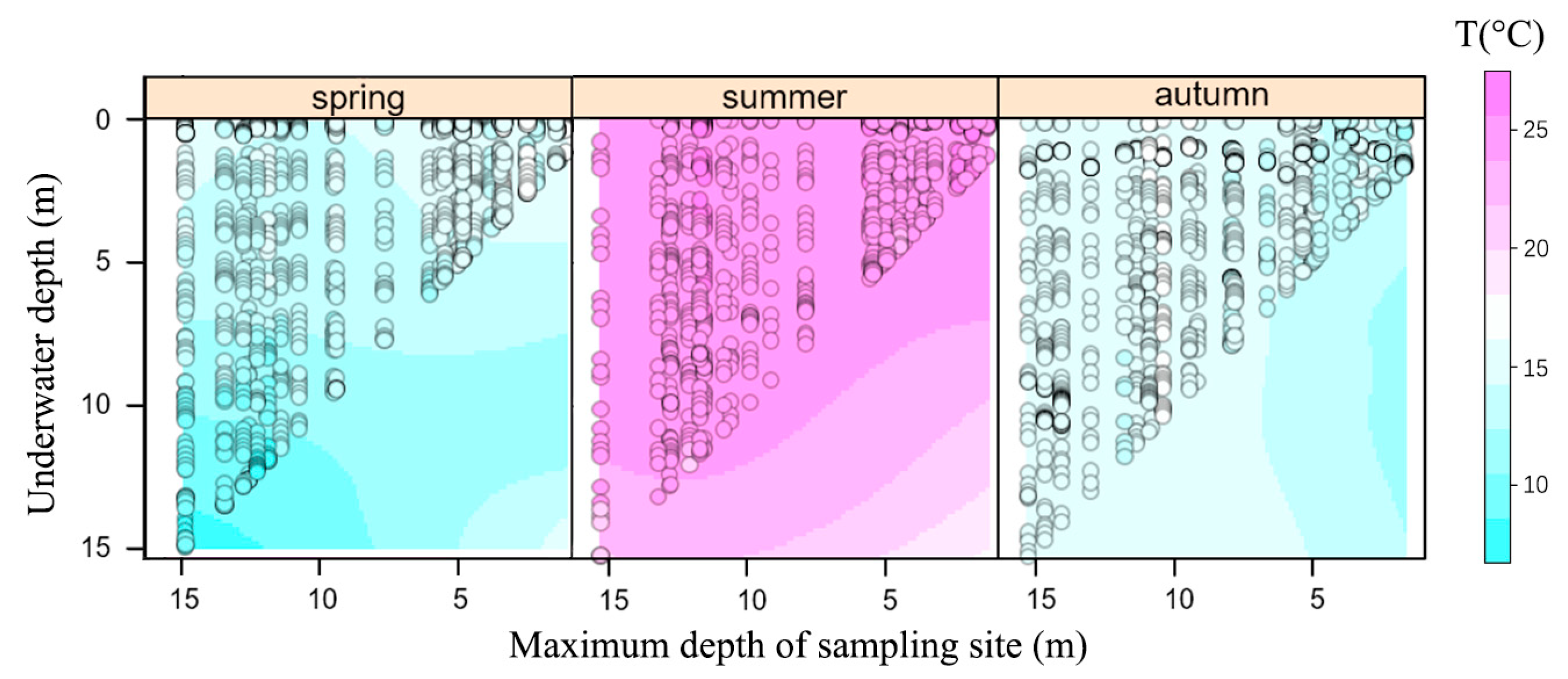
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- FAO. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2022. [Google Scholar]
- Shelton, W.L.; Rothbard, S. Exotic species in global aquaculture—A review. Isr. J. Aquacult. Bamid. 2006, 58, 3–28. [Google Scholar]
- Subasinghe, R.; Soto, D.; Jia, J. Global aquaculture and its role in sustainable development. Rev. Aquacult. 2009, 1, 2–9. [Google Scholar] [CrossRef]
- Casal, C.M.V. Global Documentation of Fish Introductions: The Growing Crisis and Recommendations for Action. Biol. Invasions 2006, 8, 3–11. [Google Scholar] [CrossRef]
- Witt, A.B.R. Use of Non-Native Species for Poverty Alleviation in Developing Economies. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P., Eds.; Invading Nature—Springer Series in Invasion Ecology; Springer: Cham, Switzerland, 2017; Volume 12. [Google Scholar]
- Lin, Y.; Gao, Z.; Zhan, A. Introduction and use of non-native species for aquaculture in China: Status, risks and management solutions. Rev. Aquacult. 2015, 7, 28–58. [Google Scholar] [CrossRef]
- Kang, B.; Vitule, J.R.S.; Li, S.; Shuai, F.; Huang, L.; Huang, X.; Huang, X.; Fang, J.; Shi, X.; Zhu, Y.; et al. Introduction of non-native fish for aquaculture in China: A systematic review. Rev. Aquacult. 2022, 15, 676–703. [Google Scholar] [CrossRef]
- Gozlan, R.E. Introduction of non-native freshwater fish: Is it all bad? Fish Fish. 2008, 9, 106–115. [Google Scholar] [CrossRef]
- Manchester, S.J.; Bullock, J.M. The impacts of non-native species on UK biodiversity and the effectiveness of control. J. Appl. Ecol. 2000, 37, 845–864. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Pilotto, F.; Innocenti, G.; Cianfanelli, S.; Haase, P. Two centuries for an almost complete community turnover from native to non-native species in a riverine ecosystem. Glob. Chang. Biol. 2021, 27, 606–623. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.M.; Bradshaw, C.J.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- Ohlberger, J.; Mehner, T.; Staaks, G.; Hölker, F. Temperature-related physiological adaptations promote ecological divergence in a sympatric species pair of temperate freshwater fish, Coregonus spp. Funct. Ecol. 2008, 22, 501–508. [Google Scholar] [CrossRef]
- Henkanaththegedara, S.M.; Stockwell, C.A. Intraguild predation may facilitate coexistence of native and non-native fish. J. Appl. Ecol. 2014, 51, 1057–1065. [Google Scholar] [CrossRef]
- Bøhn, T.; Amundsen, P.A.; Sparrow, A. Competitive exclusion after invasion? Biol. Invasions 2008, 10, 359–368. [Google Scholar] [CrossRef]
- Garrison, L.P. Spatial and dietary overlap in the Georges Bank ground fish community. Can. J. Fish Aquat. Sci. 2000, 57, 1679–1691. [Google Scholar] [CrossRef]
- Juncos, R.; Milano, D.; Macchi, P.J.; Vigliano, P.H. Niche segregation facilitates coexistence between native and introduced fishes in a deep Patagonian lake. Hydrobiologia 2015, 747, 53–67. [Google Scholar] [CrossRef]
- Westrelin, S.; Balzani, P.; Haubrock, P.J.; Santoul, F. Interannual variability in the trophic niche of young-of-year fish belonging to four piscivorous species coexisting in a natural lake. Freshwater Biol. 2023, 68, 487–501. [Google Scholar] [CrossRef]
- Winemiller, K.O. Spatial and Temporal Variation in Tropical Fish Trophic Networks. Ecol. Monogr. 1990, 60, 331–367. [Google Scholar] [CrossRef]
- Araújo, M.S.; Bolnick, D.I.; Layman, C.A. The ecological causes of individual specialisation. Ecol. Lett. 2011, 14, 948–958. [Google Scholar] [CrossRef]
- Sánchez-Hernández, J.; Nunn, A.D.; Adams, C.E.; Amundsen, P.-A. Causes and consequences of ontogenetic dietary shifts: A global synthesis using fish models. Biol. Rev. 2019, 94, 539–554. [Google Scholar] [CrossRef]
- Westrelin, S.; Cucherousset, J.; Roy, R.; Tissot, L.; Santoul, F.; Argillier, C. Habitat partitioning among three predatory fish in a temperate reservoir. Ecol. Freshw. Fish 2021, 31, 129–142. [Google Scholar] [CrossRef]
- Chrétien, E.; Chapman, L.J. Habitat heterogeneity facilitates coexistence of native fishes with an introduced predator: The resilience of a fish community 5 decades after the introduction of Nile perch. Biol. Invasions 2016, 18, 3449–3464. [Google Scholar] [CrossRef]
- Freitas, C.; Villegas-Ríos, D.; Moland, E.; Olsen, E.M. Sea temperature effects on depth use and habitat selection in a marine fish community. J. Anim. Ecol. 2021, 90, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.M.; Tófoli, R.M.; da Silva, J.C.B.; Gomes, L.C.; Agostinho, A.A. Effects of habitat complexity on trophic interactions of three congeneric fish species. Aquat. Ecol. 2022, 56, 877–889. [Google Scholar] [CrossRef]
- Mehner, T.; Busch, S.; Helland, I.P.; Emmrich, M.; Freyhof, J. Temperature-related nocturnal vertical segregation of coexisting coregonids. Ecol. Freshw. Fish 2010, 19, 408–419. [Google Scholar] [CrossRef]
- Perales, K.M.; Hansen, G.J.A.; Hein, C.L.; Mrnak, J.T.; Roth, B.M.; Walsh, J.R.; Vander Zanden, M.J. Spatial and temporal patterns in native and invasive crayfishes during a 19-year whole-lake invasive crayfish removal experiment. Freshw. Biol. 2021, 66, 2105–2117. [Google Scholar] [CrossRef]
- Li, W.J.; Gao, X.; Liu, H.Z.; Cao, W.X. Coexistence of Two Closely Related Cyprinid Fishes (Hemiculter bleekeri and Hemiculter leucisculus) in the Upper Yangtze River, China. Diversity 2020, 12, 284. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, R.; Cai, L. Fish Resources and Fishery in Lake Bosten; Xinjiang Science and Technology Press: Urumqi, China, 2005. [Google Scholar]
- Jiang, X.; Wang, J.; Tang, W.; Sun, Z.; Pan, B. Non-native freshwater fish species in the Yellow River Basin: Origin, distribution and potential risk. Environ. Biol. Fish. 2021, 104, 253–264. [Google Scholar] [CrossRef]
- Ji, F.; Ma, X.; Qiu, L.; Kang, Z.; Shen, J. Quantifying the effects of introduced Bighead Carp (Cyprinidae; Aristichthys nobilis) stocking on dominant fish species in the Ulungur Lake, China. Biol. Invasions 2022, 24, 1253–1265. [Google Scholar] [CrossRef]
- Lai, Y. The Relationship between the Disappearence of Calanoida Population and the Introduction of Hypomesus olidus in Lake Bosten. J. Hydroecol. 2009, 2, 136–139, (In Chinese with English abstract). [Google Scholar]
- Xie, Y.; Li, B. Behaviors of pond smelt (Hypomesus olidus) and the changes in its population structure in two reservoirs. Acta Ecol. Sin. 1991, 11, 37–43, (In Chinese with English abstract). [Google Scholar]
- Chen, P.; Ma, Y.; Xie, C.; Qi, F. Preliminary study on community structure of fishes in Bositeng Lake. Freshw. Fish. 2014, 44, 36–42, (In Chinese with English abstract). [Google Scholar]
- Cao, X.; Dong, S.; Yang, Z.; Gong, S. Species of genus Hypomesus for transplantation. Hebei Fish. 2009, 6, 10–12. (In Chinese) [Google Scholar]
- Gong, X.; Wu, Y.; Cui, Z.; Bao, B. Molecular identification of transplanted Hypomesus species in seven places. J. Fish. China 2012, 12, 1802–1808, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Xu, Y. The production of Lake Bosten’s pond smelt exceeds 1700 tons. Mod. Fisheries Inf. 2010, 25, 3. (In Chinese) [Google Scholar]
- Swanson, C.; Reid, T.; Young, P.S.; Cech, J.J. Comparative environmental tolerances of threatened delta smelt (Hypomesus transpacificus) and introduced wakasagi (H. nipponensis) in an altered California estuary. Oecologia 2000, 123, 384–390. [Google Scholar] [CrossRef]
- Zhou, X.; Hu, Z.; Liu, Q.; Yang, L.; Wang, Y. Feeding ecology of the non-indigenous fish Hypomesus nipponensis in Lake Ulungur, China: Insight into the relationship between its introduction and the collapse of the native Eurasian perch population. Mar. Freshw. Res. 2013, 64, 549–557. [Google Scholar] [CrossRef]
- Sakamoto, D.; Nemoto, T.; Sunoh, N.; Iwasaki, J.; Niwa, S.; Arayama, K.; Suzuki, N.; Takagi, K.; Sakuramoto, K. Population size estimation of the pond smelt Hypomesus nipponensis in Lake Kasumigaura and Lake Kitaura, Japan. Fish. Sci. 2014, 80, 907–914. [Google Scholar] [CrossRef]
- Yin, C.; Chen, Y.; Guo, L.; Ni, L. Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China. Water 2021, 13, 1800. [Google Scholar] [CrossRef]
- Zhang, T.; Yu, X.; Xian, Y.; Song, M.; Li, X.; Cai, L.; Niu, J. Appraisal and suggestions on fishery resources effectiveness evaluation of breeder releasing in Lake Bosten and Ulungur Lake. Chin. Fish. Econ. 2020, 38, 93–99, (In Chinese with English abstract). [Google Scholar]
- Cao, G. Rediscovery of Bigstract.ery resources effectiveness. Sci. Fish Farm. 1999, 1, 40. (In Chinese) [Google Scholar]
- Ren, B.; Ren, M.; Guo, Y.; Zhang, R.; Ma, Y.; Liu, Y.; Aizezi; Tuersun; Abudu. Studies on the biological characteristics of Aspiorhynchus laticeps (Day). Chin. J. Fish. 2006, 2, 9–22. (In Chinese) [Google Scholar]
- Ma, Y.; Zhang, R.; Li, H.; Tuersun, G.Y. Research of Habitat and Community Conservation Biology of Aspiorhynchus laticeps. J. Hydroecol. 2010, 31, 38–42. (In Chinese) [Google Scholar]
- Xie, Y.; Li, B. Observation on the embryonic development of pond smelt. J. Fish. China 1987, 11, 307–314. [Google Scholar]
- Wang, S.; Dou, H. Lake Records of China; Science Press: Beijing, China, 1989. [Google Scholar]
- Dytham, C. Choosing and Using Statistics: A Biologist’s Guide, 3rd ed.; Wiley-Blackwell Publishing Ltd.: Oxford, UK, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 6 March 2023).
- Mangiafico, S. rcompanion: Functions to Support Extension Education Program Evaluation, R Package Version 2.4.21; R Foundation for Statistical Computing: Vienna, Austria, 2023. Available online: https://CRAN.R-project.org/package=rcompanion(accessed on 6 March 2023).
- Carbia, P.S.; Brown, C.; Park, J.M.; Gaston, T.F.; Raoult, V.; Williamson, J.E. Seasonal and developmental diet shifts in sympatric and allopatric intertidal gobies determined by stomach content and stable isotope analysis. J. Fish Biol. 2020, 97, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, S.V.; Johnson, T.B.; Metcalfe, B.; Fisk, A.T. Spatial distribution of lake trout (Salvelinus namaycush) across seasonal thermal cycles in a large lake. Freshw. Biol. 2021, 66, 615–627. [Google Scholar] [CrossRef]
- Hunter, E.; Metcalfe, J.D.; Reynolds, J.D. Migration route and spawning area fidelity by North Sea plaice. Proc. R. Soc. B. 2003, 270, 2097–2103. [Google Scholar] [CrossRef]
- Carassou, L.; Whitfield, A.K.; Moyo, S.; Richoux, N.B. Dietary tracers and stomach contents reveal pronounced alimentary flexibility in the freshwater mullet (Myxus capensis, Mugilidae) concomitant with ontogenetic shifts in habitat use and seasonal food availability. Hydrobiologia 2017, 799, 327–348. [Google Scholar] [CrossRef]
- Heithaus, R.M.; Dill, M.L. Does tiger shark predation risk influence foraging habitat use by bottlenose dolphins at multiple spatial scales? Oikos 2006, 114, 257–264. [Google Scholar] [CrossRef]
- Heggenes, J.; Baglinière, J.L.; Cunjak, R.A. Spatial niche variability for young Atlantic salmon (Salmo salar) and brown trout (S. trutta) in heterogeneous streams. Ecol. Freshw. Fish 1999, 8, 1–21. [Google Scholar] [CrossRef]
- Day, T.; McPhail, J.D. The effect of behavioural and morphological plasticity on foraging efficiency in the threespine stickleback (Gasterosteus sp.). Oecologia 1996, 108, 380–388. [Google Scholar] [CrossRef]
- Jolles, J.W.; Briggs, H.D.; Araya-Ajoy, Y.G.; Boogert, N.J. Personality, plasticity and predictability in sticklebacks: Bold fish are less plastic and more predictable than shy fish. Anim. Behav. 2019, 154, 193–202. [Google Scholar] [CrossRef]
- Sandlund, O.T.; Museth, J.; Naesje, T.F.; Rognerud, S.; Saksgard, R.; Hesthagen, T.; Borgstrom, R. Habitat use and diet of sympatric Arctic charr (Salvelinus alpinus) and whitefish (Coregonus lavaretus) in five lakes in southern Norway: Not only interspecific population dominance? Hydrobiologia 2010, 650, 27–41. [Google Scholar] [CrossRef]
- Ye, S.; Li, Z.; Lek-Ang, S.; Feng, G.; Lek, S.; Cao, W. Community structure of small fishes in a shallow macrophytic lake (Niushan Lake) along the middle reach of the Yangtze River, China. Aquat. Living Resour. 2006, 19, 349–359. [Google Scholar] [CrossRef]
- Warfe, D.M.; Barmuta, L.A. Habitat structural complexity mediates the foraging success of multiple predator species. Oecologia 2004, 141, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Ba, W.; Du, P.; Liu, T.; Bao, A.; Chen, X.; Liu, J.; Qin, C. Impacts of climate change and agricultural activities on water quality in the Lower Kaidu River Basin, China. J. Geogr. Sci. 2020, 30, 164–176, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Pushchin, I.; Kondrashev, S.; Kamenev, Y. Retinal ganglion cell topography and spatial resolution in the Japanese smelt Hypomesus nipponensis (McAllister, 1963). J. Anat. 2021, 238, 905–916. [Google Scholar] [CrossRef]
- Fritsches, K.A.; Marshall, N.; Justin, W.E.J. Retinal specializations in the blue marlin: Eyes designed for sensitivity to low light levels. Mar. Freshwater Res. 2003, 54, 333–341. [Google Scholar] [CrossRef]
- Mehner, T. Diel vertical migration of freshwater fishes—Proximate triggers, ultimate causes and research perspectives. Freshwater Biol. 2012, 57, 1342–1359. [Google Scholar] [CrossRef]
- Andrzejaczek, S.; Gleiss, A.C.; Pattiaratchi, C.B.; Meekan, M.G. Patterns and drivers of vertical movements of the large fishes of the epipelagic. Rev. Fish Biol. Fish. 2019, 29, 335–354. [Google Scholar] [CrossRef]
- Yang, T.; Sun, J.; Wang, Z.; Qiu, Z.; Du, L.; Zhang, P. Investigation on the habitats of the pond smelt in the Chaihe Reservoir. Reserv. Fish. 1998, 6, 31–32. (In Chinese) [Google Scholar]
- Hilton, Z.; Wellenreuther, M.; Clements, K.D. Physiology underpins habitat partitioning in a sympatric sister-species pair of intertidal fishes. Funct. Ecol. 2008, 22, 1108–1117. [Google Scholar] [CrossRef]
- Busch, S.; Kirillin, G.; Mehner, T. Plasticity in habitat use determines metabolic response of fish to global warming in stratified lakes. Oecologia 2012, 170, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Hayden, B.; Harrod, C.; Kahilainen, K.K. Lake morphyometry and resource polymorphism determine niche segregation between cool- and cold-water adapted fish. Ecology 2014, 95, 538–552. [Google Scholar] [CrossRef]
- Yamada, J. The normal developmental stages of the pond smelt, Hypomesus olidus (Pallas). Bull. Fac. Fish. Hokkaido Univ. 1963, 14, 121–126. [Google Scholar]
- Hiroki, A. Early life ecology of Japanese smelt (Hypomesus nipponensis) in Lake Abashiri, a brackish water, eastern Hokkaido, Japan. Sci. Rep. Hokkaido Fish. Exp. Stn. 2004, 67, 1–79. [Google Scholar]
- Kudo, T.; Mizuguchi, K. Growth of large and small forms of pond smelt Hypomesus nipponensis in Lake Kasumigaura, Japan. Fish. Sci. 2000, 66, 432–441. [Google Scholar] [CrossRef]
- Chen, W.; Zhong, Z.; Dai, W.; Fan, Q.; He, S. Phylogeographic structure, cryptic speciation and demographic history of the sharp-belly (Hemiculter leucisculus), a freshwater habitat generalist from southern China. BMC Evol. Biol. 2017, 17, 216. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Factors | d.f. | H | p |
|---|---|---|---|---|
| Hemiculter leucisculus | Habitat | 2 | 20.18 | <0.001 |
| Season | 2 | 73.36 | <0.001 | |
| Habitat:Season | 4 | 11.01 | 0.026 | |
| Residuals | 117 | |||
| Hypomesus nipponensis | Habitat | 2 | 7.39 | 0.02 |
| Season | 2 | 24.88 | <0.001 | |
| Habitat:Season | 4 | 40.61 | <0.001 | |
| Residuals | 117 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, D.; Zhang, Y.; Li, J.; Wang, P.; Ye, S.; Zhang, T.; Li, W.; Liao, C.; Guo, C.; Liu, J. Thermal Preference May Facilitate Spatial Coexistence of Two Invasive Fish Species in Lake Bosten, China. Sustainability 2023, 15, 7592. https://doi.org/10.3390/su15097592
Song D, Zhang Y, Li J, Wang P, Ye S, Zhang T, Li W, Liao C, Guo C, Liu J. Thermal Preference May Facilitate Spatial Coexistence of Two Invasive Fish Species in Lake Bosten, China. Sustainability. 2023; 15(9):7592. https://doi.org/10.3390/su15097592
Chicago/Turabian StyleSong, Dan, Yinzhe Zhang, Junfeng Li, Puze Wang, Shaowen Ye, Tanglin Zhang, Wei Li, Chuansong Liao, Chuanbo Guo, and Jiashou Liu. 2023. "Thermal Preference May Facilitate Spatial Coexistence of Two Invasive Fish Species in Lake Bosten, China" Sustainability 15, no. 9: 7592. https://doi.org/10.3390/su15097592