

Development of Halotolerant Microbial Consortia for Salt Stress Mitigation and Sustainable Tomato Production in Sodic Soils: An Enzyme Mechanism Approach
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates Used in the Study
2.2. Extracellular Enzyme Assay
2.3. Effect of PGPR Isolates on Tomato Hybrid NS585 in Sodic Soil
2.4. Bio-Formulation of Microbial Consortia
2.5. Screening of Salt Tolerance and Sodium Uptake
2.6. Quantification of ACC Deaminase Activity
2.7. Indole Acetic Acid Production (IAA)
2.8. Effect of Different Microbial Consortia on Tomato Hybrid NS585
2.9. Estimation of Na/K ion Uptake by Plant
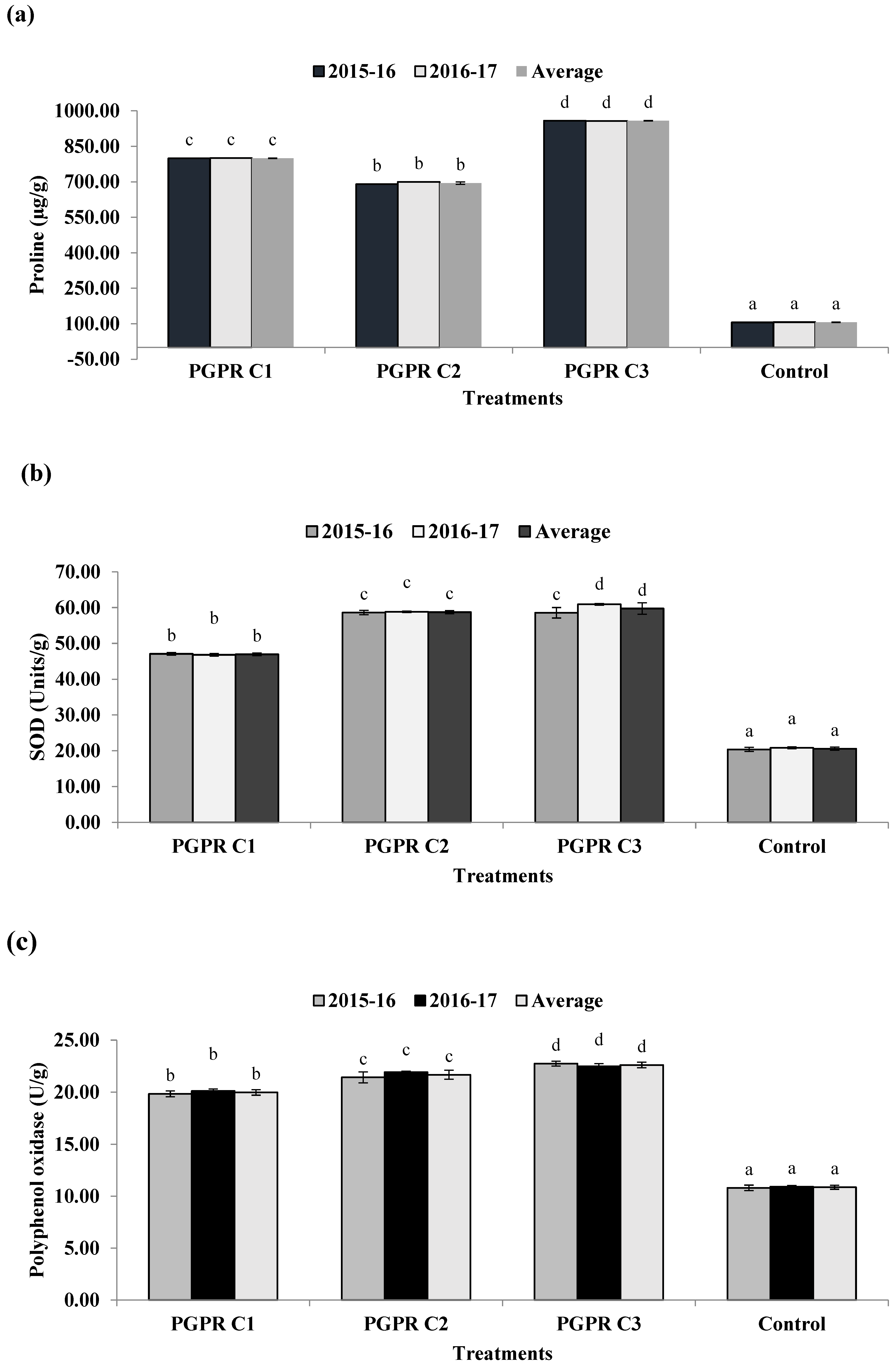
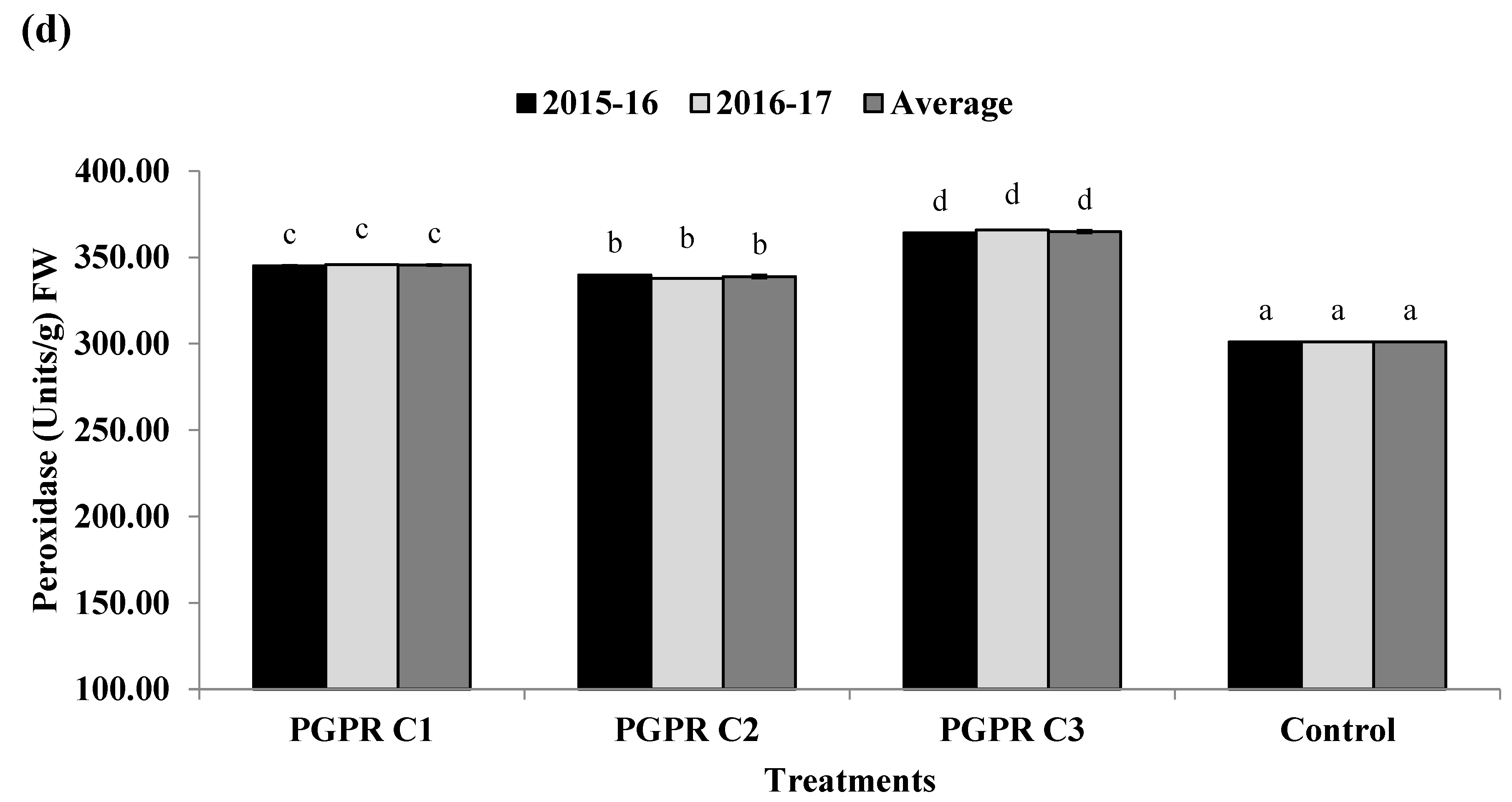
2.10. Quantification of Plant Stress Enzymes and Compounds against Salinity Stress
2.11. Statistical Analysis
3. Result
3.1. Extracellular Enzyme Assay
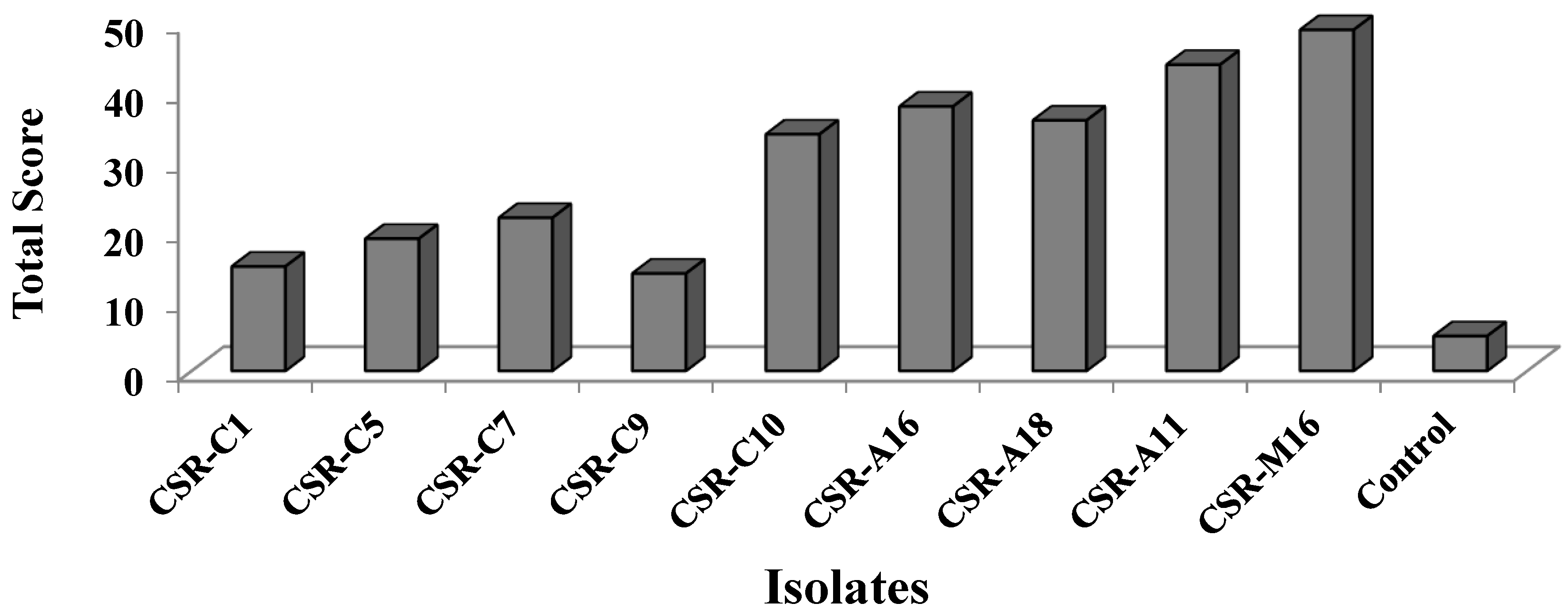
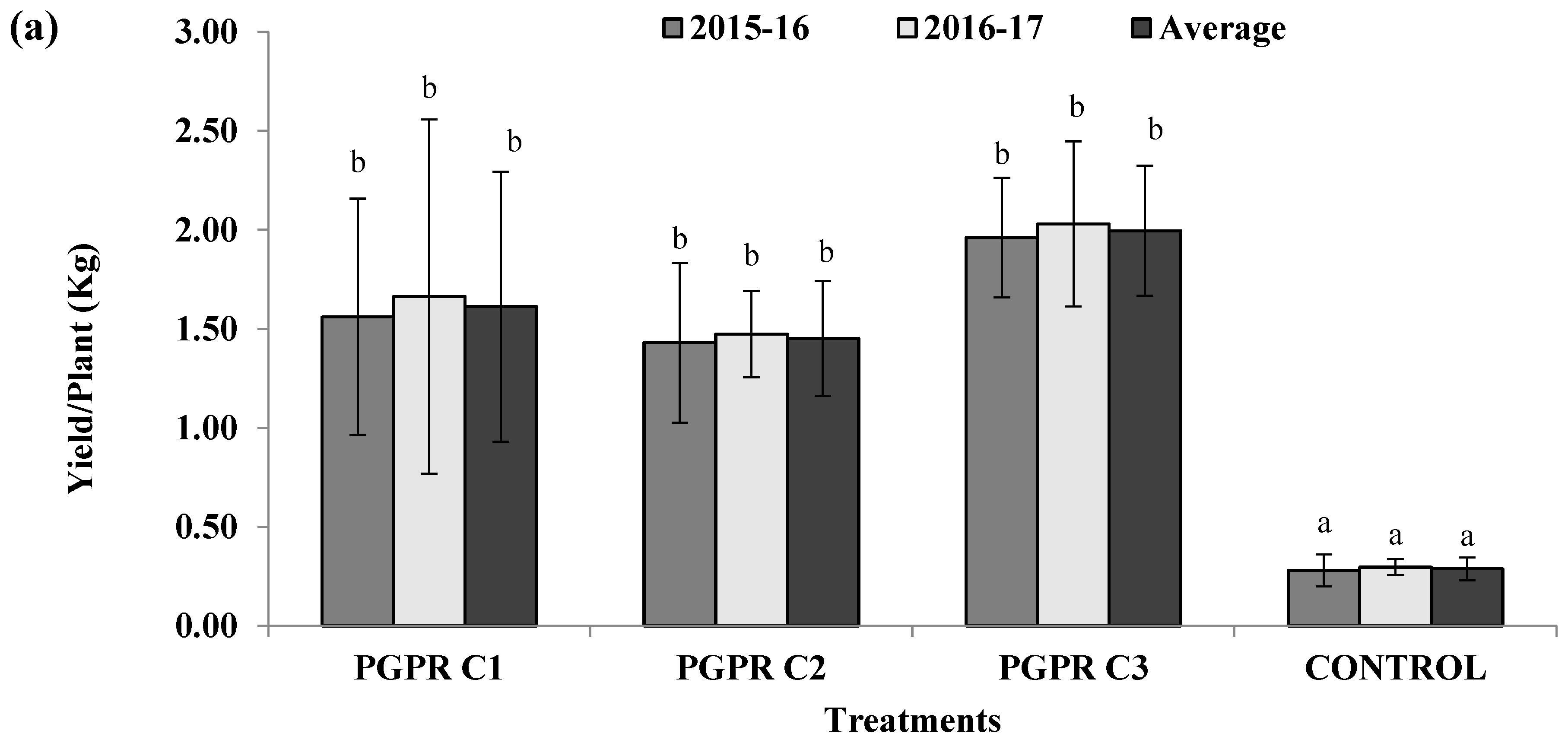
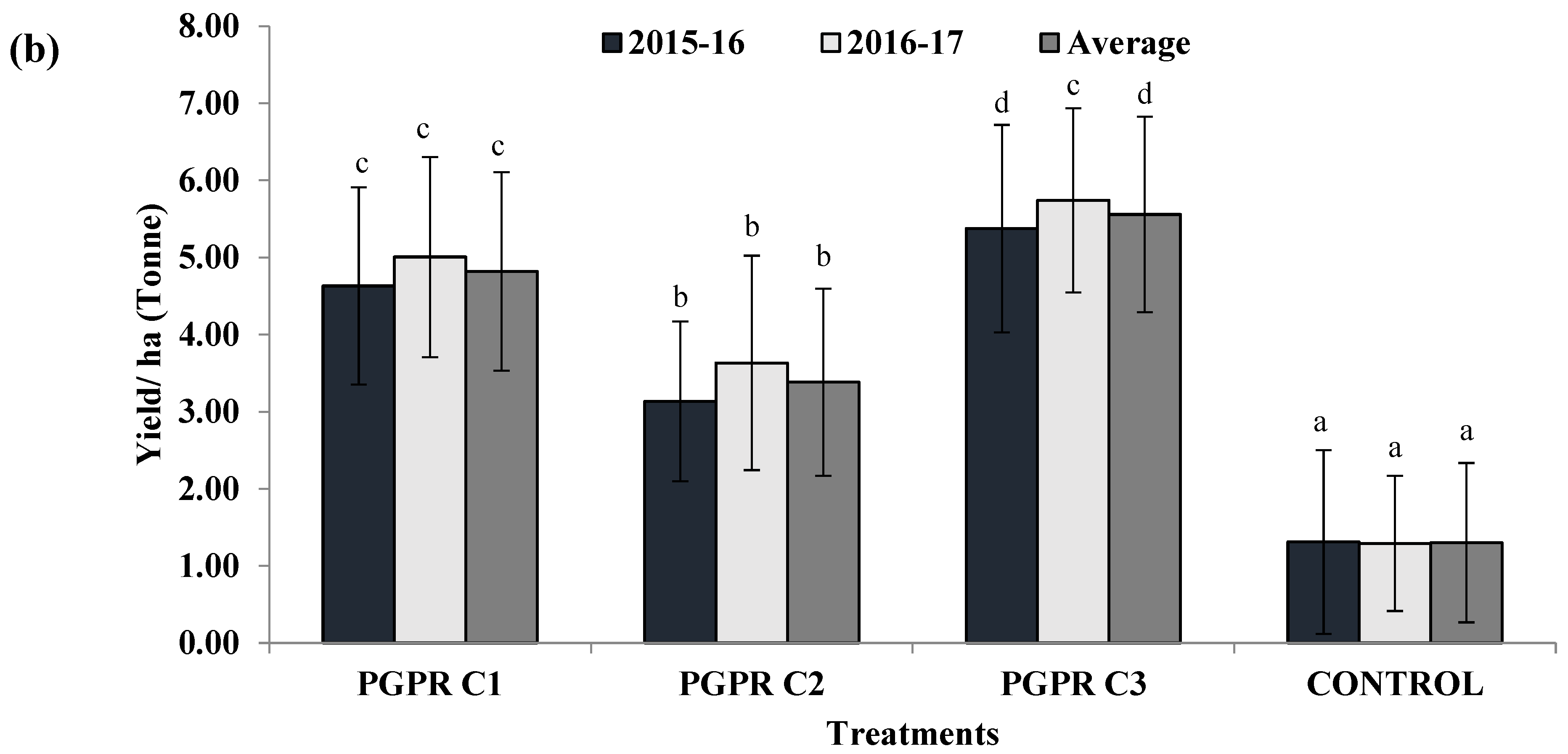
3.2. Effect of H-PGPR Isolates on Tomato Hybrid NS585 in Sodic Soil
3.3. Compatibility Assessment and Formulation of Bio-Consortia
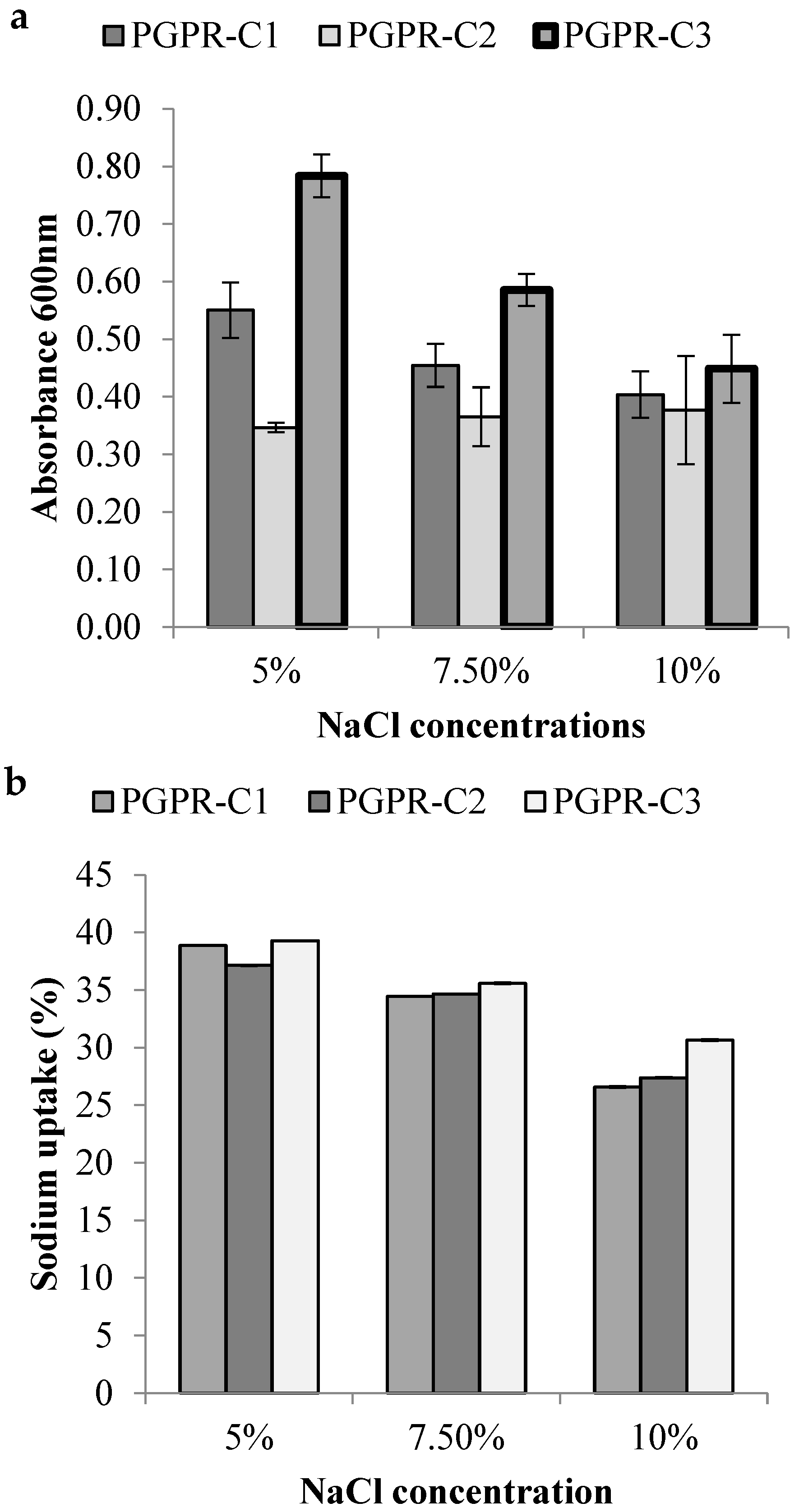
3.4. Screening of Salt Tolerance and Sodium Uptake
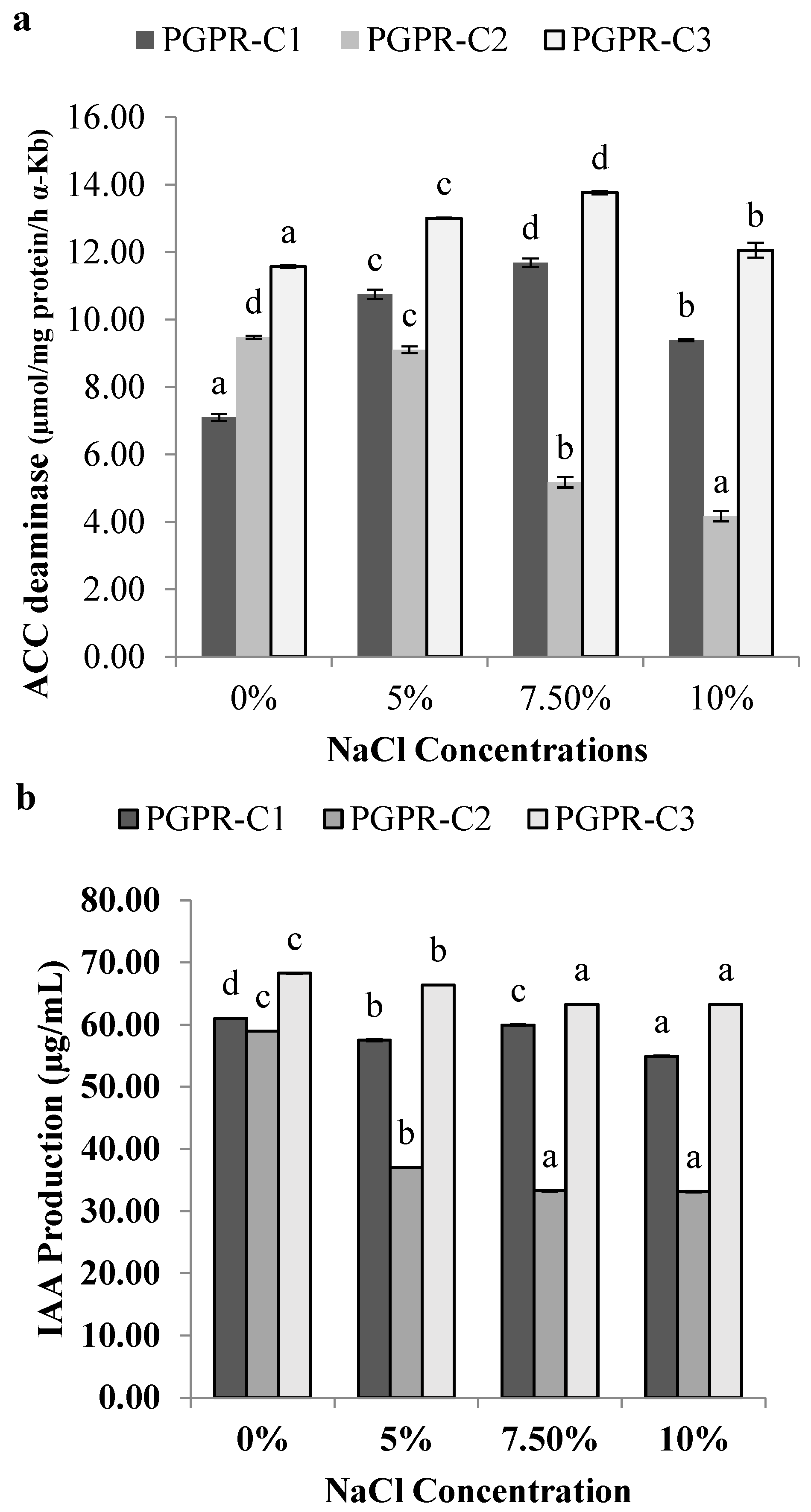
3.5. Quantification of ACC Deaminase Activity
3.6. Indole Acetic Acid Production
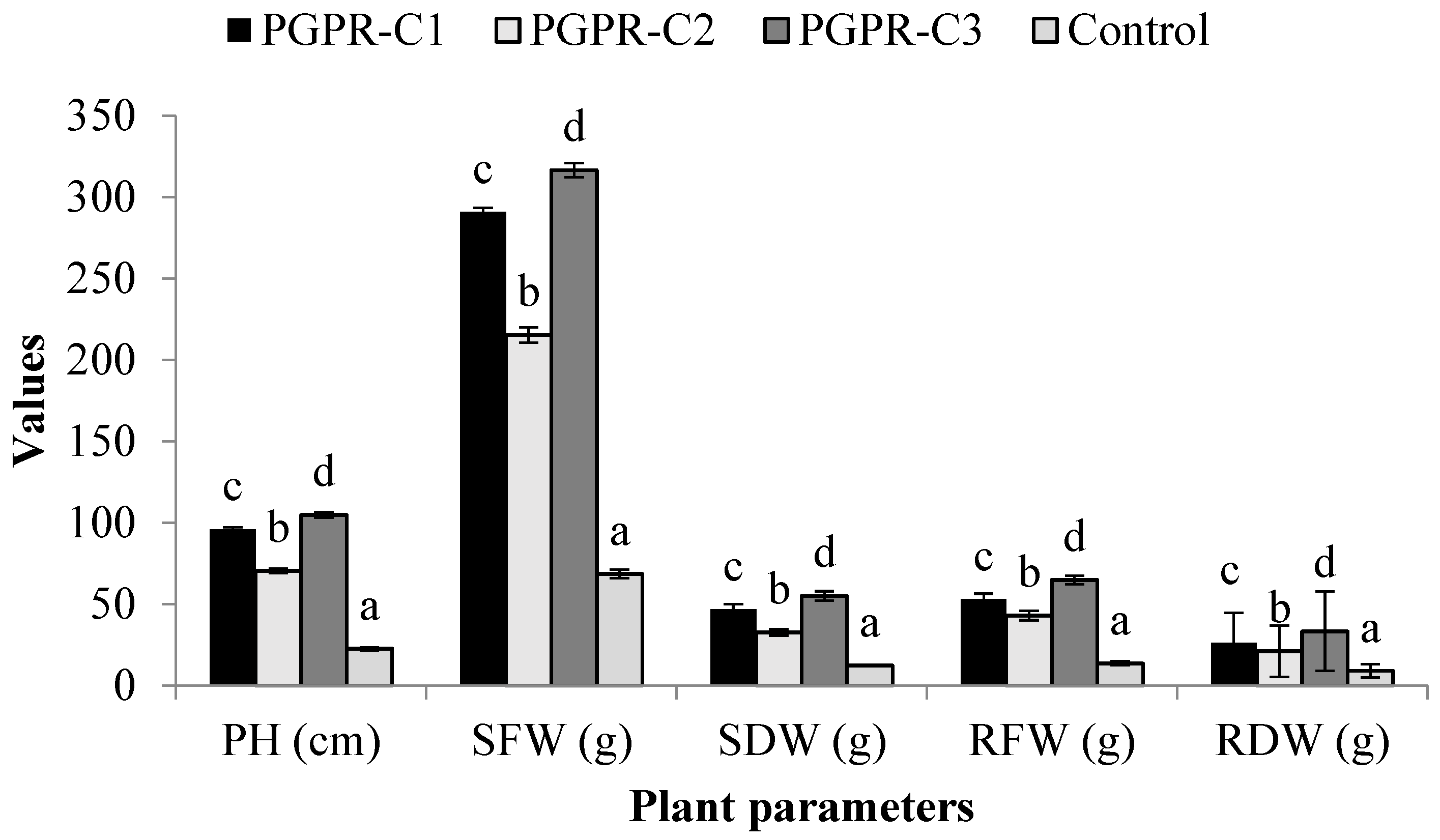
3.7. Field Trials to Evaluate Effect of Different Microbial Consortia on Tomato Hybrid NS585
3.8. Estimation of Na/K ion Uptake by Plant
3.9. Quantification of Plant Stress Enzymes and Compounds against Salinity Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fazeli-Nasab, B.; Sayyed, R.Z. Plant growth promoting rhizobacteria and salinity stress: A journey into the soil. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management, Abiotic Stress Management; Sayyed, R.Z., Arora, N.K., Reddy, M.S., Eds.; Springer: Singapore, 2019; Volume 1, pp. 21–34. [Google Scholar]
- Bannari, A.; Al-Ali, Z.M. Assessing climate change impact on soil salinity dynamics between 1987–2017 in arid landscape using Landsat TM, ETM+ and OLI Data. Remot. Sens. 2020, 12, 2794. [Google Scholar] [CrossRef]
- Schirawski, J.; Perlin, M.H. Plant-microbe interaction 2017—The good, the bad and the diverse. Int. J. Mol. Sci. 2018, 19, 1374. [Google Scholar] [CrossRef] [Green Version]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.; Reddy, M.; El Enshasy, H.; Gafur, A.; et al. Bacterial plant biostimulants: A sustainable way towards improving growth, productivity, and health of crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Gondek, M.; Weindorf, D.C.; Thiel, C.; Kleinheinz, G. Soluble salts in compost and their effects on soil and plants: A review. Compost. Sci. Util. 2020, 28, 59–75. [Google Scholar] [CrossRef]
- Liu, L.; Xia, W.; Li, H.; Zeng, H.; Wei, B.; Han, S.; Yin, C. Salinity inhibits rice seed germination by reducing α-Amylase activity via decreased bioactive Gibberellin content. Front. Plant Sci. 2018, 9, 275. [Google Scholar] [CrossRef]
- Guo, J.; Du, M.; Tian, H.; Wang, B. Exposure to high salinity during seed development markedly enhances seedling emergence and fitness of the progeny of the extreme halophyte Suaeda salsa. Front. Plant Sci. 2020, 11, 1291. [Google Scholar] [CrossRef]
- Qadir, M.; Schubert, S. Degradation process and nutrient constraints in sodic soils. Land Degrad. Dev. 2002, 13, 275–294. [Google Scholar] [CrossRef]
- Gupta, R.K.; Abrol, I.P. Salinity build up and changes in the rice-wheat system of the Indo-Gangetic plains. Exp. Agric. 2000, 36, 273–284. [Google Scholar] [CrossRef]
- Tank, N.; Saraf, M. Salinity-resistant plant growth promoting rhizobacteria ameliorates sodium chloride stress on tomato plants. J. Plant Interact. 2010, 5, 51–58. [Google Scholar] [CrossRef]
- Damodaran, T.; Rai, R.B.; Mishra, V.K.; Sharma, D.K.; Ram, R.A.; Rai, S.; Kumar, H. Integrated Farming System and Livelihood Security: An Approach; Central Soil Salinity Research Institute (CSSRI): Karnal, India, 2011; pp. 1–108. [Google Scholar]
- Ayyam, V.; Palanivel, S.; Chandrakasan, S. Approaches in land degradation management for productivity enhancement. In Coastal Ecosystems of the Tropics—Adaptive Management; Ayyam, V., Palanivel, S., Chandrakasan, S., Eds.; Springer: Singapore, 2019; pp. 463–491. [Google Scholar]
- Keren, R. Salt-affected soils, reclamation. In Encyclopedia of Soils in the Environment; Academic Press: Cambridge, MA, USA, 2005; pp. 454–461. [Google Scholar]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-tolerant plant growth promoting rhizobacteria for enhancing crop productivity of saline soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [Green Version]
- Shilev, S. Plant-growth-promoting bacteria mitigating soil salinity stress in plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Bottini, R.; Cassan, F.; Piccoli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biotechol. 2004, 65, 497–503. [Google Scholar] [CrossRef]
- Dodd, I.C.; Zinovkina, N.Y.; Safronova, V.I.; Belimov, A.A. Rhizobacterial mediation of plant hormone status. Ann. Appl. Biol. 2010, 157, 361–379. [Google Scholar] [CrossRef]
- Naz, I.; Bano, A.; Hassan, T.U. Isolation of phytohormones producing plant growth promoting rhizobacteria from weeds growing in Khewra salt range, Pakistan and their implication in providing salt tolerance to Glycine max L. Afri. J. Biotechnol. 2009, 8, 5762–5766. [Google Scholar]
- Patel, T.; Saraf, M. Biosynthesis of phytohormones from novel rhizobacterial isolates and their in vitro plant growth promoting efficacy. J. Plant Interact. 2017, 12, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, S.K.; Maurya, S.K.; Singh, D.P. Salinity tolerance in free living plant growth promoting rhizobacteria. Indian J. Sci. Res. 2012, 3, 73–78. [Google Scholar]
- Timmusk, S.; Abd El-Daim, I.A.; Copolovici, L.; Tanilas, T.; Kannaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenström, E.; Niinemets, U. Drought tolerance of Wheat improved by rhizosphere bacteria from harsh environments: Enhanced biomass production and reduced emissions of stress volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [Green Version]
- Bano, A.; Fatima, M. Salt tolerance in Zea mays (L). following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, D.P. Effect of salt-tolerant plant growth promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol. 2015, 17, 288–293. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, 1089. [Google Scholar] [CrossRef] [Green Version]
- Ansari, F.A.; Ahmad, I.; Pichtel, J. Growth stimulation and alleviation of salinity stress to wheat by the biofilm forming Bacillus pumilus strain FAB10. Appl. Soil. Ecol. 2019, 3, 45–54. [Google Scholar] [CrossRef]
- Tahir, M.; Ahmad, I.; Shahid, M.; Shah, G.M.; Farooq, A.B.U.; Akram, M.; Tabassum, S.A.; Naeem, M.A.; Khalid, U.; Ahmad, S.; et al. Regulation of antioxidant production, ion uptake and productivity in potato (Solanum tuberosum L.) plant inoculated with growth promoting salt tolerant Bacillus strains. Ecotox. Environ. Safe 2019, 178, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Kavadia, A.; Omirou, M.; Fasoula, D.; Ioannides, I.M. The importance of microbial inoculants in a climate-changing agriculture in eastern Mediterranean region. Atmosphere 2020, 11, 1136. [Google Scholar] [CrossRef]
- Damodaran, T.; Mishra, V.K.; Jha, S.K.; Pankaj, U.; Gupta, G.; Gopal, R. Identification of rhizosphere bacterial diversity with promising salt tolerance, PGP traits and their exploitation for seed germination enhancement in sodic soil. J. Agric. Res. 2019, 8, 36–43. [Google Scholar] [CrossRef]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A Rapid and easy method for the detection of microbial cellulases on agar plates using gram’s Iodine. Curr. Microbiol. 2008, 57, 503–550. [Google Scholar] [CrossRef]
- Kavitha, T.; Nelson, R.; Jesi, S.J. Screening of rhizobacteria for plant growth promoting traits and antifungal activity against charcoal rot pathogen Macrophomina phaseolina. Int. J. Pharma Bio. Sci. 2013, 4, 177–186. [Google Scholar]
- Harley, M.; Prescott, J. Laboratory Exercise in Microbiology, 5th ed.; The McGraw-Hill Companies: New York City, NY, USA, 2002. [Google Scholar]
- Rai, R.B.; Damodaran, T.; Kannan, R.; Rathore, R.S.; Srivastava, A.P.; Sharma, D.K.; Mishra, V.K.; Jha, S.K.; Sah, V. Low Cost Multiplication Technology of Salt Tolerant Bio-Growth Enhancers (Bacillus, Pseudomonads & Trichoderma) for Increasing Productivity of Agri-Horti Crops in Normal and Sodic Soils. Patent No. 3857/DEL/2012, 14 December 2012. [Google Scholar]
- Herdiyantoro, D.; Setiawati, M.R.; Simarmata, T.; Nurlaeny, N.; Joy, B.; Hamdani, J.S.; Handayani, I. The ability of potassium solubilizing rhizo-bacteria isolated from maize rhizosphere for microbial fertilizer. Proc. IOP Conf. Ser. Earth Environ. Sci. 2018, 205, 012011. [Google Scholar] [CrossRef]
- Damodaran, T.; Rai, R.B.; Jha, S.K.; Kannan, R.; Pandey, B.K.; Sah, V.; Mishra, V.K.; Sharma, D.K. Rhizosphere and endophytic bacteria for induction of salt tolerance in gladiolus grown in sodic soils. J. Plant Interact. 2013, 9, 577–584. [Google Scholar] [CrossRef] [Green Version]
- Honma, M.; Shimomura, T. Metabolism of 1-aminocyclo-propanr-1-carboxylic acid. Agric. Biol. Chem. 1978, 42, 1825–1831. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, A.R.; Fish, W.W.; Perkins-veazie, P. A Rapid hexane-free method for analyzing lycopene content in watermelon. J. Food Sci. 2003, 68, 328–332. [Google Scholar] [CrossRef]
- Watson, R.; Pritchard, J.; Malone, M. Direct measurement of sodium and potassium in the transpiration stream of salt-excluding and non-excluding varieties of wheat. J. Exp. Bot. 2001, 52, 1873–1881. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Thimmaiah, S.R. Standard Methods of Biochemical Analysis; Kalyani Publishers: New Delhi, India, 2012; pp. 421–426. [Google Scholar]
- El-Moshaty, B.; Pike, F.L.S.M.; Novacky, A.J.; Sehgal, O.P. Lipid peroxidation & superoxide production in cowpea (Vigina auguiculata) leaves infected with tobacco ring spot virus or southern bean mosaic virus. Physiol. Mol. Plant Path. 1993, 43, 109–119. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Oktay, M.; Kufrevioglu, I.; Kocacaliskan, I.; Sakiraglu, H. Polyphenoloxidase from Amasya Apple. J. Food Sci. 1995, 60, 494–496. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1982; p. 507. [Google Scholar]
- Subiramani, S.; Ramalingam, S.; Muthu, T.; Nile, S.H.; Venkidasamy, B. Development of abiotic stress tolerance in crops by plant growth promoting rhizobacteria (PGPR). In Phyto-Microbiome in Stress Regulation; Kumar, M., Etesami, H., Kumar, V., Eds.; Springer: Singapore, 2020; pp. 125–145. [Google Scholar]
- Feng, K.; Yu, J.; Cheng, Y.; Ruan, M.; Wang, R.; Ye, Q.; Zhou, G.; Li, Z.; Yao, Z.; Yang, Y.; et al. The SOD Gene family in tomato: Identification, phylogenetic relationships, and expression patterns. Front. Plant Sci. 2016, 30, 1279. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Hong, Y.; Zhu, G.; Li, Y.; Niu, Q.; Yao, J.; Hua, K.; Bai, J.; Zhu, Y.; Shi, H.; et al. Loss of salt tolerance during tomato domestication conferred by variation in a Na+/K+ transporter. EMBO J. 2020, 18, e103256. [Google Scholar] [CrossRef]
- Damodaran, T.; Mishra, V.K. Rhizosphere engineering: An innovative approach for sustainable crop production in sodic soils. In Innovative Saline Agriculture; Dagar, J., Sharma, P., Sharma, D., Singh, A., Eds.; Springer: New Delhi, India, 2016; p. 224. [Google Scholar]
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. Springer Plus 2013, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kusale, S.P.; Attar, Y.C.; Sayyed, R.Z.; El Enshasy, H.; Hanapi, S.Z.; Ilyas, N.; Elgorban, A.; Bahkali, A.; Marraiki, N. Inoculation of Klebsiella variicola alleviated salt stress and improved growth and nutrients in wheat and maize. Agronomy 2021, 11, 927. [Google Scholar] [CrossRef]
- Karthika, S.; Midhun, S.J.; Jisha, M. A potential antifungal and growth-promoting bacterium Bacillus sp. KTMA4 from tomato rhizosphere. Microb. Pathog. 2020, 142, 104049. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Microbiol. Res. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Asadullah; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of Salt Tolerant PGPR in growth and yield augmentation of Wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Fayzullaeva, M.; Sulaymonov, K.; Kadirova, D.; Jabbarov, Z.; Sayyed, R.Z. Isolation and characterization of endophytic bacteria from ginger (Zingiber officinale Rosc.). Ann. Phytomed. 2020, 9, 116–121. [Google Scholar] [CrossRef]
- Mokrani, S.; Nabti, E.; Cruz, C. Current advances in plant growth promoting bacteria alleviating salt stress for sustainable agriculture. Appl. Sci. 2020, 10, 7025. [Google Scholar] [CrossRef]
- Kapadia, C.; Sayyed, R.Z.; Enshasy, H.E.; Vaidya, H.; Sharma, D.; Patel, V.; Malek, R.; Syed, A.; Elgorban, A.; Ahmad, K.; et al. Halotolerant microbial consortia for sustainable mitigation of salinity stress, growth promotion, and mineral uptake in tomato plant and soil nutrient enrichment. Sustainability 2021, 13, 8369. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, D.P.; Saikia, R. Genetic diversity of plant growth promoting rhizobacteria isolated from rhizospheric soil of wheat under saline condition. Curr. Microbiol. 2009, 59, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Bano, A. Role of PGPR in the reclamation and revegetation of saline land. Pak. J. Bot. 2019, 51, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boubekri, K.; Soumare, A.; Mardad, I.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. The screening of potassium- and phosphate-solubilizing Actinobacteria and the assessment of their ability to promote wheat growth parameters. Microorganisms 2021, 9, 470. [Google Scholar] [CrossRef]
- Korir, H.; Mungai, N.W.; Thuita, M.; Hamba, Y.; Masso, C. Co-inoculation effect of rhizobia and plant growth promoting rhizobacteria on common bean growth in a low phosphorus soil. Front. Plant Sci. 2017, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.; Zebelo, S.; Mcnear, D.; Kloepper, J.; Fadamiro, H. Plant growth-promoting rhizobacteria induce changes in Arabidopsis thaliana gene expression of nitrate and ammonium uptake genes. J. Plant Interact. 2019, 14, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.D.; Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonas improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sellitto, V.M.; Golubkina, N.A.; Pietrantonio, L.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Florin, I.; Caruso, G. Tomato yield, quality, mineral composition and antioxidants as affected by beneficial microorganisms under soil salinity induced by balanced nutrient solutions. Agriculture 2019, 9, 110. [Google Scholar] [CrossRef] [Green Version]
- Mahanta, D.; Rai, R.K.; Mishra, S.D.; Raja, A.; Purakayastha, T.J.; Varghese, E. Influence of phosphorus and biofertilizers on soybean and wheat root growth and properties. Field Crops Res. 2014, 166, 1–9. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops-what is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Strahsburger, E.; Zapata, F.; Pedroso, I.; Fuentes, D.; Tapia, P.; Ponce, R.; Valdes, J. Draft genome sequence of Exiguobacterium aurantiacum strain PN47 isolate from saline ponds, known as “Salar del Huasco”, located in the Altiplano in the North of Chile. Braz. J. Microbiol. 2018, 49, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defense mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, A.; Yadav, S.; Singh, I.K. Reactive oxygen species-mediated signaling during abiotic stress. Plant Gene 2019, 18, 100173. [Google Scholar] [CrossRef]
- Myrene, R.D.; Devaraj, V.R. Biochemical responses of Hyacinth bean (Lablab purpureus) to salinity stress. Acta Physiol. Plant. 2010, 32, 341–353. [Google Scholar]
- Acuña, J.J.; Campos, M.; de la Luz Mora, M.; Jaisi, D.P.; Jorquera, M.A. ACCD-producing rhizobacteria from an Andean altiplano native plant (Parastrephia quadrangularis) and their potential to alleviate salt stress in wheat seedlings. Appl. Soil Ecol. 2019, 136, 184–190. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Parameters | Values |
|---|---|
| Soil pH | 9.14 |
| EC | 0.52 ds m−1 |
| Sodium (Na+) | 15.79 meq L−1 |
| Potassium (K+) | 0.13 meq L−1 |
| Carbonate | 2.63 meq L−1 |
| Bicarbonate | 1.62 meq L−1 |
| Isolates Name | Isolates Code | Amylase | Protease | Cellulase | Lipase |
|---|---|---|---|---|---|
| Pseudomonas sp. | CSR-C1 | - | - | - | - |
| Agromyces tropicus | CSR-C5 | - | - | - | - |
| Bacillus flexus | CSR-C7 | ++ | - | - | + |
| Bacillus tequilensis | CSR-C9 | ++ | - | ++ | - |
| Pseudomonas mendocina | CSR-C10 | - | - | - | - |
| Lysinibacillus sphaericus | CSR-A16 | ++ | ++ | ++ | ++ |
| Bacillus subtilis | CSR-A18 | + | + | ++ | + |
| Lysinibacillus fusiformis | CSR-A11 | + | ++ | ++ | ++ |
| Bacillus licheniformis | CSR-M16 | ++ | ++ | ++ | + |
| Plant Parameters | |||||
|---|---|---|---|---|---|
| Treat. | Pl. Ht. | SFWt. | SDWt. | RFWt. | RDWt. |
| (cm) | (g) | (g) | (g) | (g) | |
| CSR-C1 | 32.23 b | 45.38 cd | 5.40 bc | 8.90 b | 1.66 a |
| CSR-C5 | 38.37 cd | 43.78 c | 4.45 ab | 10.33 b | 2.03 ab |
| CSR-C7 | 39.97 d | 47.86 d | 5.56 bc | 3.97 a | 2.83 b |
| CSR-C9 | 36.33 c | 40.71 b | 3.57 a | 9.67 b | 1.93 ab |
| CSR-C10 | 57.57 e | 63.24 e | 6.54 cd | 17.57 de | 6.40 d |
| CSR-A16 | 66.37 f | 75.73 fg | 7.54 de | 16.43 d | 6.43 d |
| CSR-A18 | 66.33 f | 73.02 f | 8.07 ef | 14.57 c | 4.83 c |
| CSR-A11 | 65.93 f | 78.37 g | 9.02 f | 17.23 de | 6.82 de |
| CSR-M16 | 71.85 g | 78.28 g | 9.02 f | 18.57 e | 7.73 e |
| Control | 20.34 a | 24.70 a | 3.72 a | 2.70 a | 1.49 a |
| F value | 488.88 * | 344.31 * | 23.02 * | 135.49 * | 50.23 * |
| Percent Increment (%) | ||||||
|---|---|---|---|---|---|---|
| Consortia | Isolates | Pl. Ht. | SFWt. | SDWt. | RFWt. | RDWt. |
| PGPR-C1 | CSR-A11 | 31.26 | 73.04 | 80.65 | 67.48 | 81.56 |
| CSR-A18 | 30.84 | 74.88 | 82.69 | 72.5 | 75.46 | |
| CSR-M16 | 25.09 | 73.07 | 80.65 | 64.96 | 70.5 | |
| PGPR-C2 | CSR-A11 | 6.39 | 63.6 | 72.37 | 59.87 | 76.99 |
| CSR-A18 | 5.82 | 66.08 | 75.28 | 66.07 | 69.37 | |
| CSR-A16 | 5.76 | 64.82 | 76.9 | 61.74 | 69.51 | |
| PGPR-C3 | CSR-A11 | 37.12 | 25.26 | 83.59 | 73.41 | 85.5 |
| CSR-A16 | 36.7 | 27.77 | 86.28 | 74.64 | 80.78 | |
| CSR-M16 | 31.47 | 25.34 | 83.59 | 76.79 | 76.79 | |
| Year | PGPR | TSS% | Na+/K+ | Lycopene (mg/100 g) |
|---|---|---|---|---|
| 2015–16 | PGPR-C1 | 5.47 b | 0.21 b | 9.52 b |
| PGPR-C2 | 5.47 b | 0.45 c | 12.50 c | |
| PGPR-C3 | 8.07 c | 0.15 a | 17.23 d | |
| Control | 3.00 a | 0.91 d | 3.98 a | |
| Total | 5.50 | 0.43 | 10.81 | |
| 2016–17 | PGPR-C1 | 5.40 b | 0.21 b | 9.67 b |
| PGPR-C2 | 5.43 b | 0.46 c | 12.93 c | |
| PGPR-C3 | 8.03 c | 0.17 a | 17.19 d | |
| Control | 3.07 a | 0.92 d | 4.07 a | |
| Average | Total | 5.48 | 0.46 | 10.97 |
| PGPR-C1 | 5.45 b | 0.22 b | 9.60 b | |
| PGPR-C2 | 5.45 b | 0.46 c | 12.71 c | |
| PGPR-C3 | 8.05 c | 0.17 a | 17.21 d | |
| Control | 3.03 a | 0.92 d | 4.03 a | |
| Total | 5.49 | 0.44 | 10.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damodaran, T.; Jha, S.K.; Kumari, S.; Gupta, G.; Mishra, V.K.; Sharma, P.C.; Gopal, R.; Singh, A.; Jat, H.S. Development of Halotolerant Microbial Consortia for Salt Stress Mitigation and Sustainable Tomato Production in Sodic Soils: An Enzyme Mechanism Approach. Sustainability 2023, 15, 5186. https://doi.org/10.3390/su15065186
Damodaran T, Jha SK, Kumari S, Gupta G, Mishra VK, Sharma PC, Gopal R, Singh A, Jat HS. Development of Halotolerant Microbial Consortia for Salt Stress Mitigation and Sustainable Tomato Production in Sodic Soils: An Enzyme Mechanism Approach. Sustainability. 2023; 15(6):5186. https://doi.org/10.3390/su15065186
Chicago/Turabian StyleDamodaran, Thukkaram, Sunil Kumar Jha, Sangeeta Kumari, Garima Gupta, Vinay K. Mishra, Parbodh C. Sharma, Ram Gopal, Arjun Singh, and Hanuman S. Jat. 2023. "Development of Halotolerant Microbial Consortia for Salt Stress Mitigation and Sustainable Tomato Production in Sodic Soils: An Enzyme Mechanism Approach" Sustainability 15, no. 6: 5186. https://doi.org/10.3390/su15065186