
Environmental Factors’ Effects on Stem Radial Variations of Populus euphratica in the Lower Reaches of the Tarim River in Northwestern China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.2.1. Micro-Core Sampling
2.2.2. Micro-Core Sample Processing
2.3. Environmental Data Collection
2.4. Data Analysis
3. Results
3.1. Meteorological Factors and Groundwater Change Characteristics
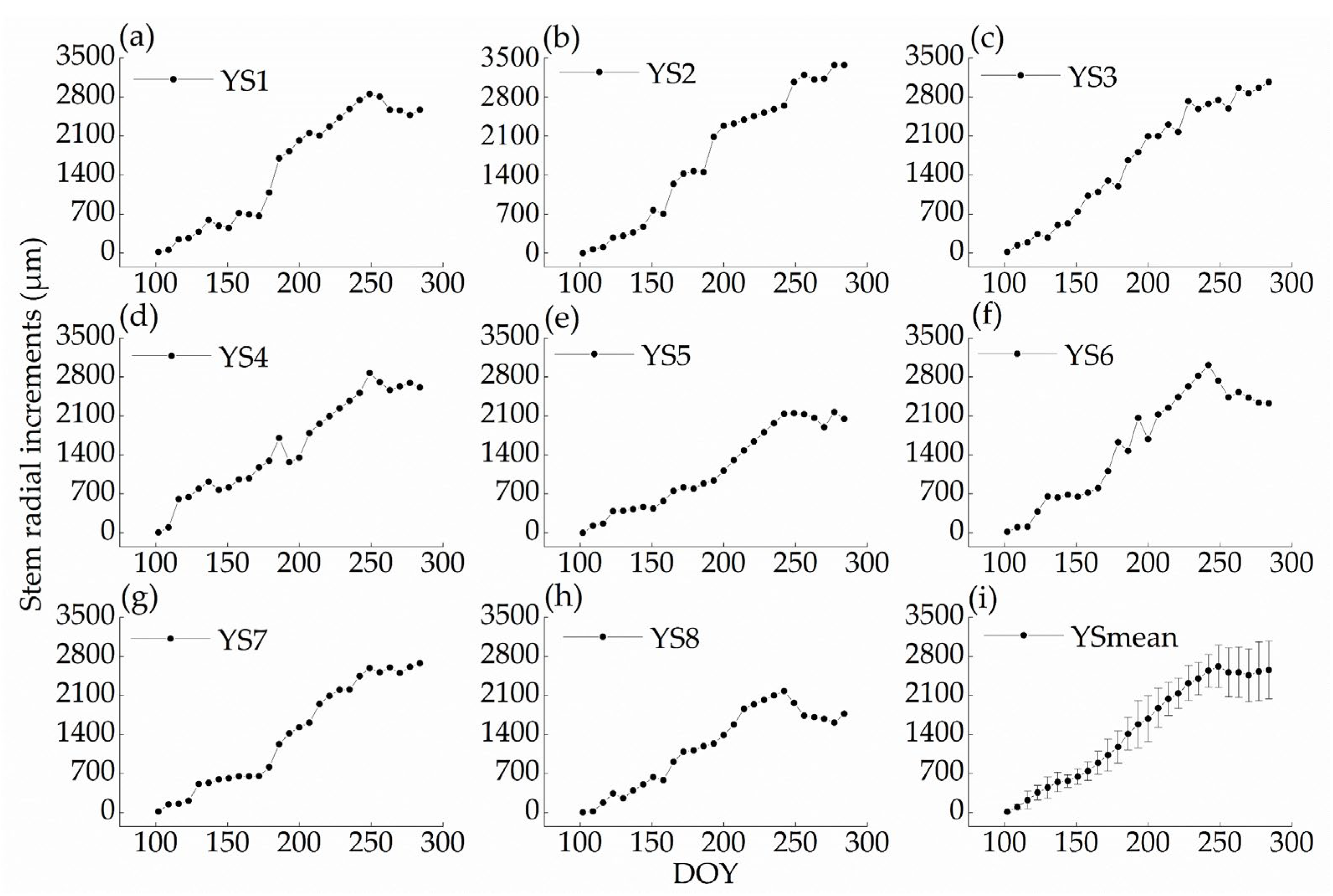
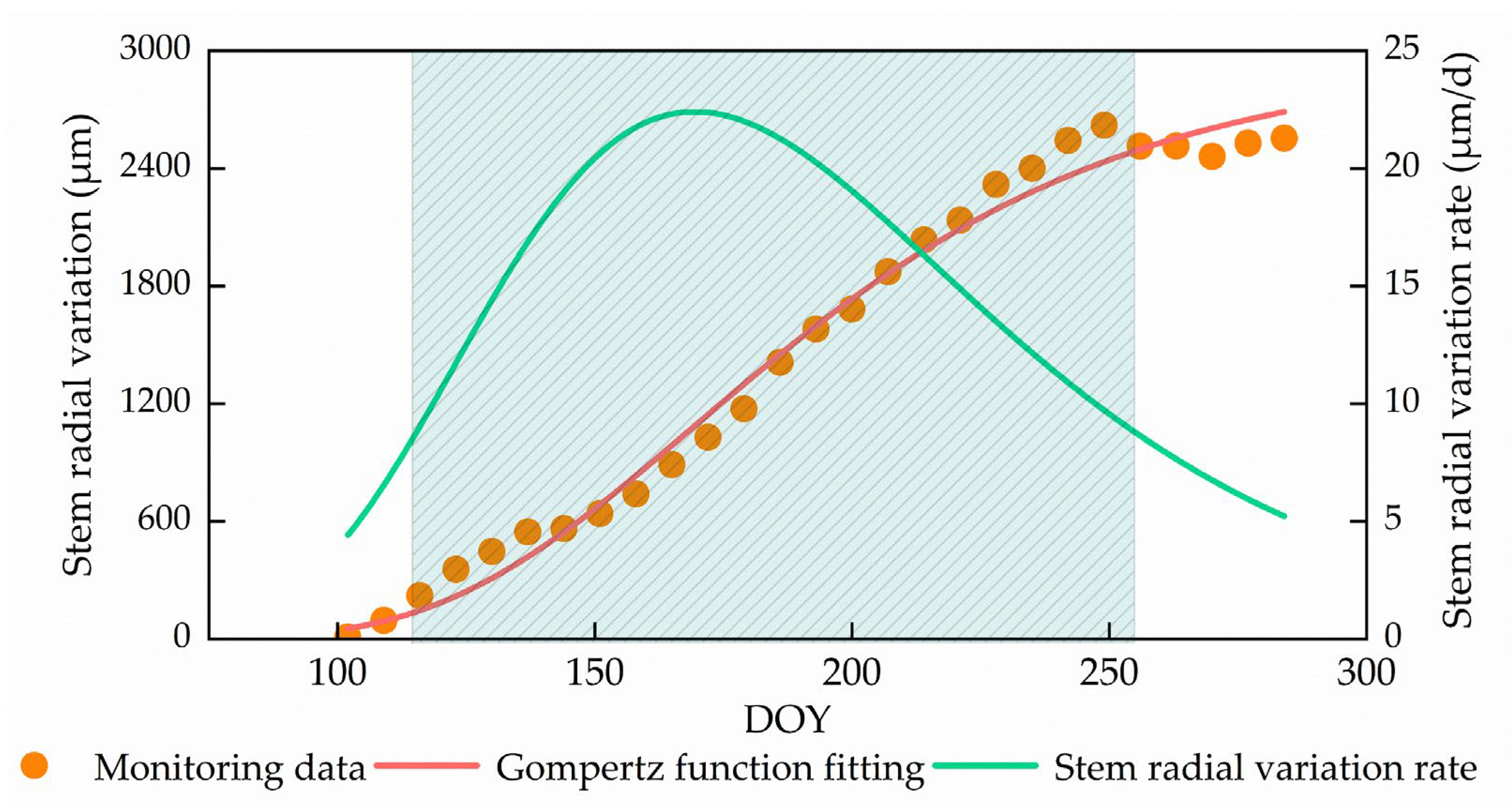
3.2. Stem Radial Variations Dynamics of P. euphratica during the Year
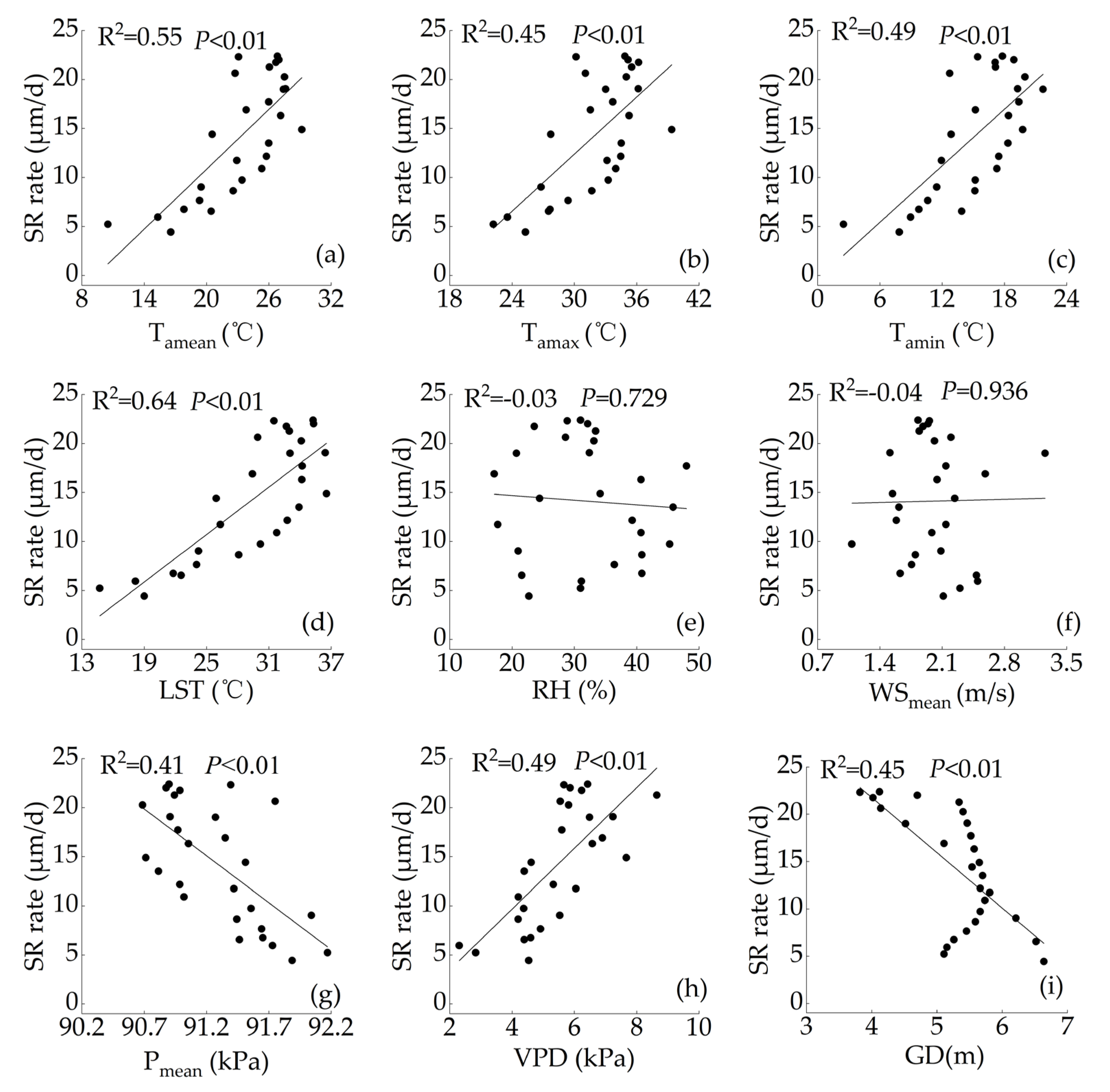
3.3. The Relationship between the Stem Radial Variation Rate of P. euphratica and Environmental Factors
3.4. Relationships between Environmental Factors That Affect Stem Radial Variation in P. euphratica
3.5. Contribution of Hydrothermal Factors to Stem Radial Variation
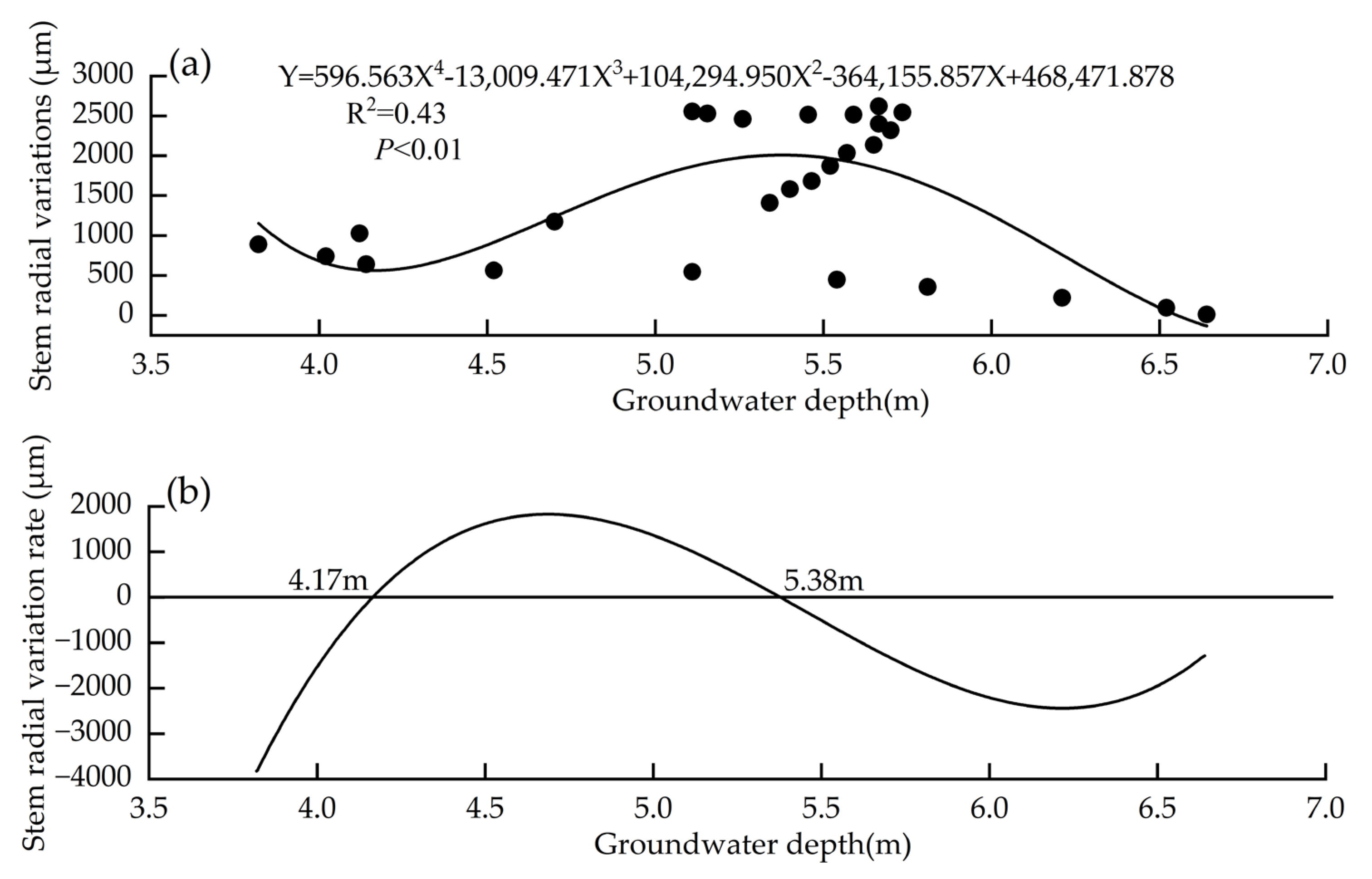
3.6. Range of Groundwater Variability Suitable for P. euphratica Growth
4. Discussion
4.1. Stem Radial Variations of P. euphratica
4.2. The Relationship between the Stem Radial Variation Rate of P. euphratica and Environmental Factors
4.3. Groundwater Levels Suitable for Growing P. euphratica
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Anderson, G.M.; Bartuska, A.; Ciais, P.; Cullenward, D.; Field, C.B.; Freeman, J.; Goetz, S.J.; et al. Climate-driven risks to the climate mitigation potential of forests. Science 2020, 368, eaaz7005. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Karger, D.N.; Wilson, A.M. Spatial detection of alpine tree line ecotones in the Western United States. Remote Sens. Environ. 2020, 240, 111672. [Google Scholar] [CrossRef]
- Huang, J.G.; Bergeron, Y.; Zhai, L.H.; Denneler, B.H. Variation in intra-annual radial growth (xylem formation) of Picea mariana (Pinaceae) along a latitudinal gradient in western Québec, Canada. Am. J. Bot. 2011, 98, 792–800. [Google Scholar] [PubMed] [Green Version]
- Cuny, H.E.; Rathgeber, C.B.; Frank, D.; Fonti, P.; Fournier, M. Kinetics of tracheid development explain conifer tree-ring structure. New Phytol. 2014, 203, 1231–1241. [Google Scholar]
- Cheng, R.M.; Liu, Z.B.; Feng, X.H.; Xiao, W.F. Advances in Research on the Effect of Climatic Change on Xylem Growth of Trees. Sci. Silvae Sin. 2015, 51, 147–154. [Google Scholar]
- Huang, J.G.; Guo, X.; Rossi, S.; Zhai, L.H.; Yu, B.Y.; Zhang, S.K.; Zhang, M.F. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree Physiol. 2018, 38, 1225–1236. [Google Scholar]
- Deslauriers, A.; Rossi, S.; Anfodillo, T.; Saracino, A. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar]
- Oladi, R.; Pourtahmasi, K.; Eckstein, D.; Bräuning, A. Seasonal dynamics of wood formation in Oriental beech (Fagusorientalis Lipsky) along an altitudinal gradient in the Hyrcanian forest, Iran. Trees 2011, 25, 425–433. [Google Scholar]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; Huang, J.; et al. Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere. Glob. Chang. Biol. 2016, 22, 3804–3813. [Google Scholar]
- Liu, J.-z.; Chen, Y.-n.; Chen, Y.-j.; Zhang, N.; Li, W.-h. Degradation of Populus euphratica community in the lower reaches of the Tarim River, Xinjiang, China. J. Environ. Sci. 2005, 17, 740–747. [Google Scholar]
- Kirdyanov, A.; Hughes, M.; Vaganov, E.; Schweingruber, F.; Silkin, P. The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees 2003, 17, 61–69. [Google Scholar]
- Yuan, Y.J.; Li, J.F. Relationship between annual ring climate increment and climateof spruce forest in western Tianshan Mountains. J. Xinjiang Univ. (Nat. Sci. Ed.) 1994, 4, 93–98. [Google Scholar]
- Wimmer, R.; Grabner, M. Effects of climate on vertical resin duct density and radial growth of Norway spruce [Picea abies (L.) Karst]. Trees 1997, 11, 271–276. [Google Scholar]
- Vázquez-González, C.; López-Goldar, X.; Zas, R.; Sampedro, L. Neutral and climate-driven adaptive processes contribute to explain population variation in resin duct traits in a Mediterranean pine species. Front. Sci. 2019, 10, 1613. [Google Scholar]
- Bowman, D.M.J.S.; Williamson, G.J.; Keenan, R.J.; Prior, L.D. A warmer world will reduce tree growth in evergreen broadleaf forests: Evidence from Australian temperate and subtropical eucalypt forests. Glob. Ecol. Biogeogr. 2014, 23, 925–934. [Google Scholar]
- Zhai, L.; Bergeron, Y.; Huang, J.G.; Berninger, F. Variation in intra-annual wood formation, and foliage and shoot development of three major Canadian boreal tree species. Am. J. Bot. 2012, 99, 827–837. [Google Scholar] [PubMed] [Green Version]
- Gou, X.H.; Deng, Y.; Chen, F.H.; Yang, M.X.; Gao, L.L.; Nesje, A.; Fang, K.Y. Precipitation variations and possible forcing factors on the Northeastern Tibetan Plateau during the last millennium. Quat. Res. 2014, 81, 508–512. [Google Scholar]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Pannatier, E.G.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar]
- Zhou, H.; Chen, Y.; Zhu, C.; Li, Z.; Fang, G.; Li, Y.P.; Fu, A. Climate change may accelerate the decline of desert riparian forest in the lower Tarim River, Northwestern China: Evidence from tree-rings of Populus euphratica. Ecol. Indic. 2020, 111, 105997. [Google Scholar]
- Zhou, H.; Chen, Y.; Hao, X.; Zhao, Y.; Fang, G.; Yang, Y. Tree rings: A key ecological indicator for reconstruction of groundwater depth in the lower Tarim River, Northwest China. Ecohydrology 2019, 12, e2142. [Google Scholar]
- Aishan, T.; Halik, Ü.; Betz, F.; Tiyip, T.; Ding, J.; Nuermaimaiti, Y. Stand structure and height-diameter relationship of a degraded Populus euphratica forest in the lower reaches of the Tarim River, Northwest China. J. Arid Land 2015, 7, 544–554. [Google Scholar]
- Ye, M.; Xu, H.L.; Song, Y.D. Analysis of water resources utilization and its change trend in tarim river basin. Chin. Sci. Bull. 2006, S1, 14–20. [Google Scholar]
- Xu, H.; Mao, Y.E.; Li, J. Changes in groundwater levels and the response of natural vegetation to transfer of water to the lower reaches of the Tarim River. J. Environ. Sci. 2007, 19, 1199–1207. [Google Scholar]
- Zhao, Z.Y.; Wang, R.H.; Zhang, H.Z. The ecological mechanism of natural vegetation restoration in the lower reaches of the Tarim River. Arid Area Res. 2005, 22, 94–100. [Google Scholar]
- Liu, H.L. Study on the Response of Radial Growth of Populus euphratica to Water in Different Sections of Tarim River. Master’s Thesis, Xinjiang Normal University, Urumqi, China, 2015. [Google Scholar]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by micro sampling tree species: An example at the Alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar]
- Creber, G.T.; Chaloner, W.G. Influence of environmental factors on the wood structure of living and fossil trees. Bot. Rev. 1984, 50, 357–448. [Google Scholar]
- Wertz, E.L.; Georgr, S.S.; Zeleznik, J.D. Vessel anomalies in Quercus macrocarpa tree rings associated with recent floods along the red river of the North, United States. Water Resour. Res. 2013, 49, 630–634. [Google Scholar]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree steams. LAWA J. 2006, 27, 89–97. [Google Scholar]
- Rathgeber, C.B.K.; Rossi, S.; Bontemps, J.D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.Z. Dynamic Monitoring of Tree Cambium Activity and Annual Radial Growth in Qilian Mountains. Ph.D. Thesis, Lanzhou University of China, Lanzhou, China, 2018. [Google Scholar]
- Abernethy, K.; Bush, E.R.; Forget, P.M.; Mendoza, I.; Morellato, L.P.C. Current issues in tropical phenology: A synthesis. Biotropica 2018, 50, 477–482. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. Res. 2003, 33, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Tixier, A.; Guzman-Delgado, P.; Sperling, O.; Roxas, A.A.; Laca, E.; Zwieniecki, M.A. Comparison of phenological traits, growth patterns, and seasonal dynamics of non-structural carbohydrate in Mediterranean tree crop species. Sci. Rep. 2020, 10, 347. [Google Scholar] [CrossRef] [Green Version]
- Luo, T.X.; Liu, X.S.; Zhang, L.; Li, X.; Pan, Y.D.; Wright, L.J. Summer solstice marks a seasonal shift in temperature sensitivity of stem growth and nitrogen use efficiency in cold-limited forests. Agric. Meteorol. 2018, 248, 469–478. [Google Scholar] [CrossRef]
- Mcdowell, N.; Pockman, W.T.; Allen, C.D.; David, D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought. New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Keenan, R.J. Climate change impacts and adaptation in forest management: A review. Ann. For. Sci. 2015, 72, 145–167. [Google Scholar] [CrossRef] [Green Version]
- Denne, M.P. Temperature and tracheid development in Pinus sylvestris seedlings. J. Exp. Bot. 1971, 22, 362–370. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Songtrirat, P.; Buajan, S.; Preechamart, S.; Chareonwong, U.; Muangsong, C. Climate control of cambial dynamics and tree-ring width in two tropical pines in Thailand. Agric. For. Meteorol. 2021, 303, 108394. [Google Scholar] [CrossRef]
- Oberhuber, W.; Stumböck, M.; Kofler, W. Climate-tree-growth relationships of Scots pine stands (Pinus sylvestris L.) exposed to soil dryness. Trees 1998, 13, 19–27. [Google Scholar] [CrossRef]
- Liu, C.; Du, T.; Li, F.; Kang, S.; Li, S.; Tong, L. Trunk sap flow characteristics during two growth stages of apple tree and its relationships with affecting factors in an arid region of northwest China. Agric. Water Manag. 2012, 104, 193–202. [Google Scholar] [CrossRef]
- McDowell, N.G.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669–672. [Google Scholar] [CrossRef]
- Volland-Voigt, F.; Bräuning, A.; Ganzhi, O.; Peters, T.; Maza, H. Radial stem variations of Tabebuia chrysantha (Bignoniaceae) in different tropical forest ecosystems of southern Ecuador. Trees 2010, 25, 39–48. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Alexander, M.R.; Xia, J.; Wang, F.; Zhang, F.; Man, Z.; Pederson, N. Drought limit wood production of Juniperus przewalskii even as growing reasons lengthens in a cold and arid environment. Catena 2021, 196, 104936. [Google Scholar] [CrossRef]
- Zhou, H.H.; Chen, Y.N.; Li, W.H.; Chen, Y.P. Photosynthesis of Populus euphratica in relation to groundwater depths and high temperature in arid environment, northwest China. Photosynthetica 2010, 48, 257–268. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Xu, C.; Li, W. Groundwater depth affects the daily course of gas exchange parameters of Populus euphratica in arid areas. Environ. Earth Sci. 2012, 66, 433–440. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DBH | rmax | rmean | tp | Start Growth | Stop Growth | Length of Time | Annual Growth Volume |
|---|---|---|---|---|---|---|---|
| cm | μm/d | μm/d | DOY | DOY | DOY | d | μm |
| 26.57 ± 7.43 | 22.42 | 13.71 | 169.43 | 114.34 | 255.06 | 140.72 | 2620.89 |
| Model | Regression Equation | R2 | p |
|---|---|---|---|
| Linear Function | Y = 68.918X + 1110.999 | −0.04 | 0.792 |
| Quadratic Function | Y = −931.447X2 + 9661.474X − 23,111.022 | 0.37 | <0.01 |
| Cubic Function | Y = −536.574X3 + 7586.062X2 − 34,667.570X + 52,382.228 | 0.41 | <0.01 |
| Quartic Function | Y = 596.563X4 − 13,009.471X3 + 104,294.950X2 − 364,155.857X + 468,471.878 | 0.43 | <0.01 |
| Logarithmic Function | Y = 884.683 × In(X + 0.033) | −0.03 | <0.01 |
| Exponential Function | Y = 1253.566 × 1.031X | −0.04 | <0.01 |
| Power Function | Y = 861.356 × X0.324 | −0.03 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Q.; Ye, M.; Zhao, X.; Pan, X. Environmental Factors’ Effects on Stem Radial Variations of Populus euphratica in the Lower Reaches of the Tarim River in Northwestern China. Sustainability 2023, 15, 11556. https://doi.org/10.3390/su151511556
He Q, Ye M, Zhao X, Pan X. Environmental Factors’ Effects on Stem Radial Variations of Populus euphratica in the Lower Reaches of the Tarim River in Northwestern China. Sustainability. 2023; 15(15):11556. https://doi.org/10.3390/su151511556
Chicago/Turabian StyleHe, Qingzhi, Mao Ye, Xin Zhao, and Xiaoting Pan. 2023. "Environmental Factors’ Effects on Stem Radial Variations of Populus euphratica in the Lower Reaches of the Tarim River in Northwestern China" Sustainability 15, no. 15: 11556. https://doi.org/10.3390/su151511556