


Potential Efficiency of Wild Plant Species (Pluchea dioscoridis (L.) DC.) for Phytoremediation of Trace Elements on Contaminated Locations


Abstract
:1. Introduction
2. Materials and Methods
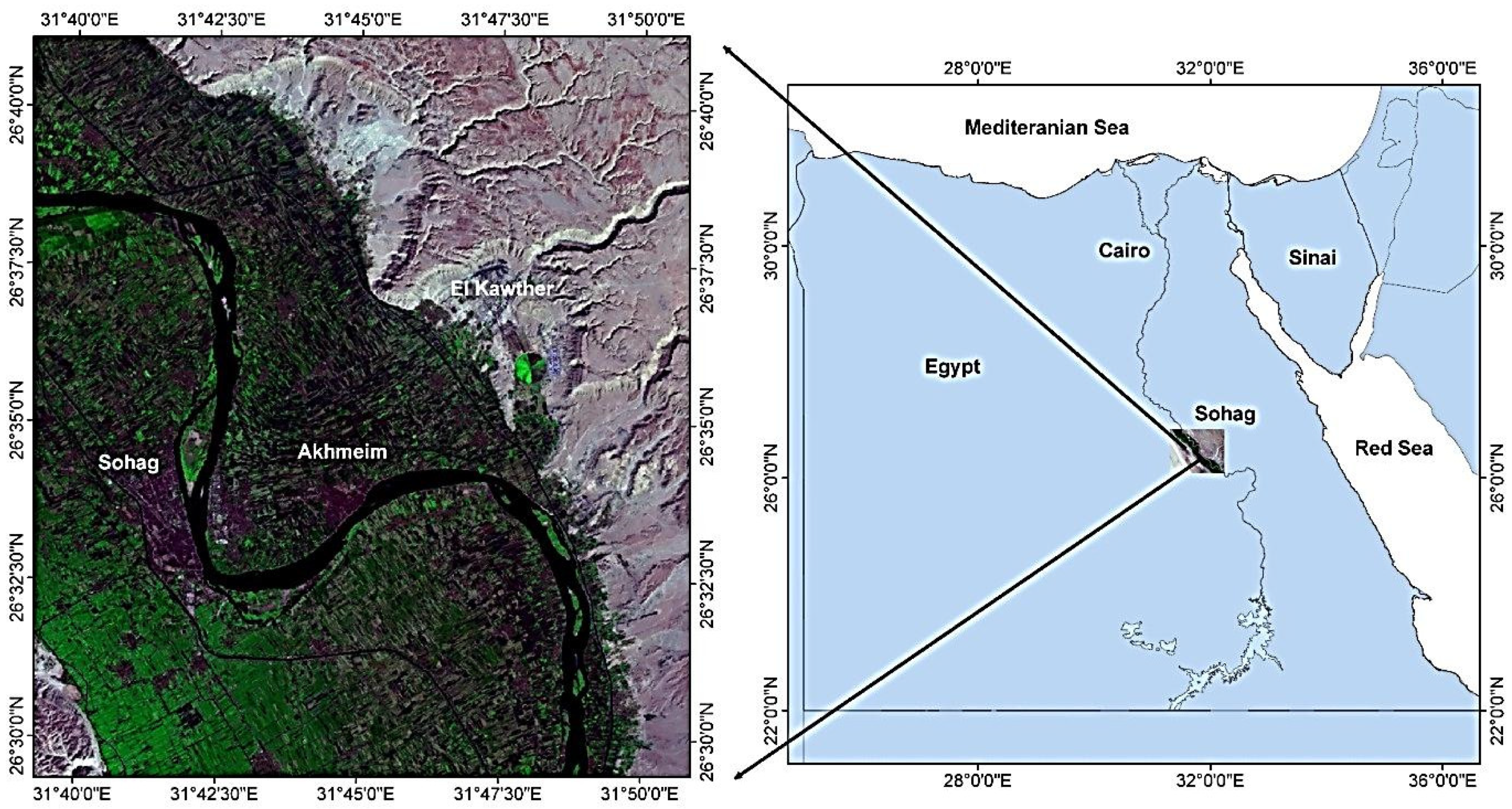
2.1. Description of the Study Area
2.2. Growth Parameters
2.3. Shoot Biomass Estimation
2.4. Root Biomass Estimation
2.5. Estimation of Pigments, Alkaloids and Flavonoids
2.6. Proline and Soluble Carbohydrates Estimation
2.7. Plant Analysis for NPK and TEs
2.8. Soil Sampling and Analysis
2.9. BCF, TF, ECS, ECR, and MAI Calculation
2.10. Data Analysis
3. Results
3.1. Growth Parameters
3.2. Estimation of the Shoot and Root Biomass
3.3. Changes in Pigments, Alkaloids, and Flavonoids
3.4. Changes in Proline Content and Soluble Carbohydrate
3.5. Nutrients (NPK) and Trace Elements (TEs) Accumulation in Plant
3.6. Soil Analysis
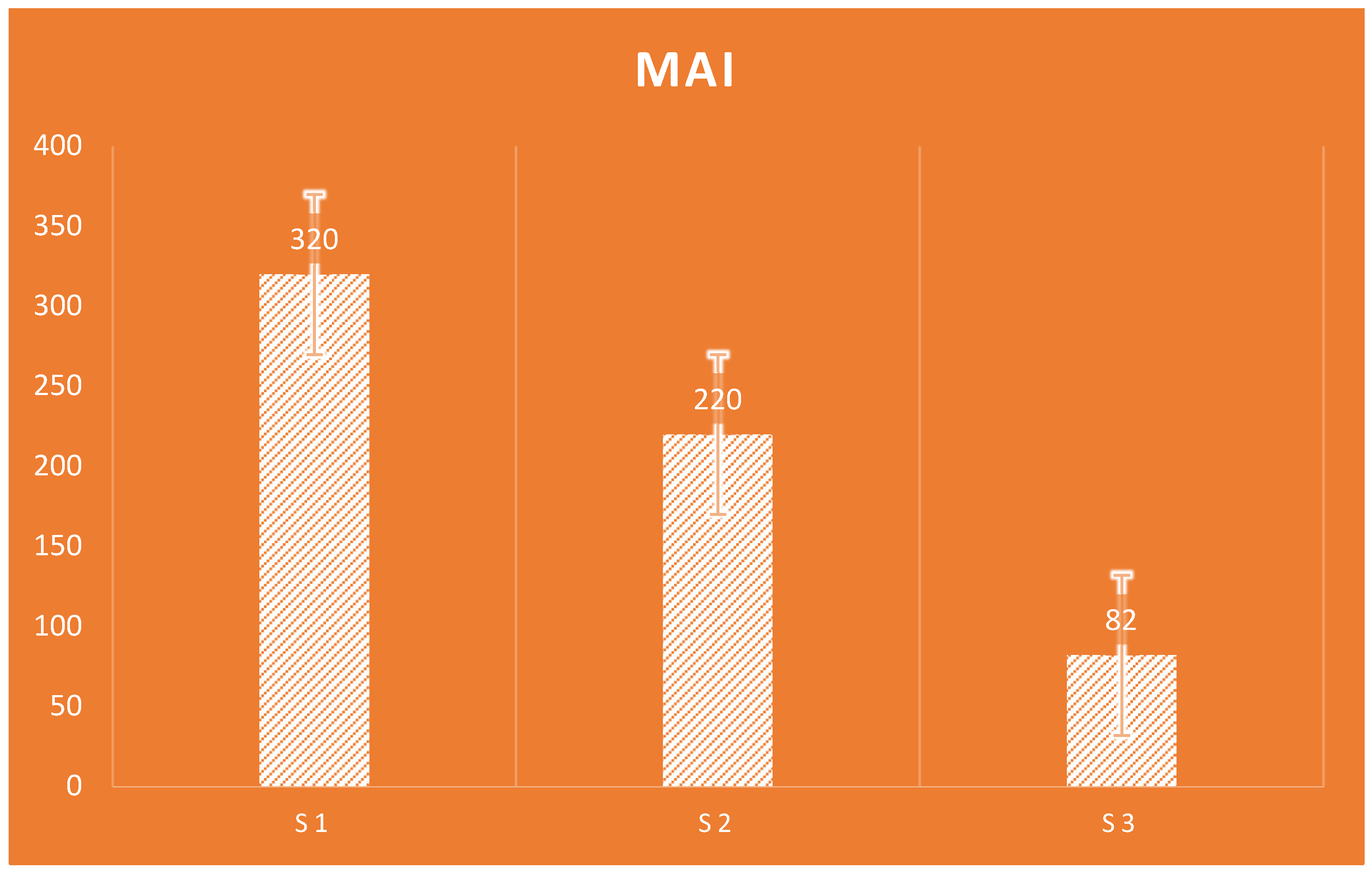
3.7. BCF, TF, ECS, ECR, MAI Indices
3.8. Plant-Soil Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TEs | Trace Elements |
| Car/Chl | Carotenoids/Chlorophyll |
| TF | Translocation factors |
| ECR | Enrichment coefficient for root |
| ECS | Enrichment coefficient for shoot |
| MAI | Metal accumulation index |
| ROS | Reactive oxygen species |
| LA | Leaf area |
| SLA | Specific leaf area |
| WC | Water content |
| RWC | Relative water content |
| FW | Fresh weight |
| DW | Dry weight |
| TW | Turbid weight |
| NPK | Nitrogen, Phosphorus, Potassium |
| AQ | Aqua regia |
| BCF | Bioconcentration factors |
| EC | Enrichment coefficients |
References
- Ng, J.C.; Juhasz, A.; Smith, E.; Naidu, R. Assessing the bioavailability and bioaccessibility of metals and metalloids. Environ. Sci. Pollut. Res. 2015, 22, 8802–8825. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Nawaz, M.F.; Gul, S.; Rizwan, M.; Ali, S.; Kareem, A. Uptake and distribution of minerals and heavy metals in commonly grown leafy vegetable species irrigated with sewage water. Environ. Monit. Assess. 2016, 188, 541. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, Y.; Yun, Z.; He, B.; Jiang, G. Estimation of bioaccessibility and potential human health risk of mercury in Chinese patent medicines. J. Environ. Sci. 2016, 39, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Sumiahadi, A.; Acar, R. A review of phytoremediation technology: Heavy metals uptake by plants. IOP Conf. Ser. Earth Environ. Sci. 2018, 142, 012023. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Rizwan, M.; Farid, M.; Shakoor, M.B.; Gill, R.A.; Najeeb, U.; Iqbal, N.; Ahmad, R. Citric acid assisted phytoremediation of copper by Brassica napus L. Ecotoxicol. Environ. Saf. 2015, 120, 310–317. [Google Scholar] [CrossRef]
- El-Khatib, A.A.; Youssef, N.A.; Barakat, N.A.; Samir, N.A. Responses of Eucalyptus globulus and Ficus nitida to different potential of heavy metal air pollution. Int. J. Phytoremediation 2020, 22, 986–999. [Google Scholar] [CrossRef]
- Naglaa, A.Y. Changes in the morphological traits and the essential oil content of sweet basil (Ocimum basilicum L.) as induced by cadmium and lead treatments. Int. J. Phytoremediation 2020, 23, 291–299. [Google Scholar]
- Qu, H.J.; Zhang, H.D.; Tan, Z.C.; Chen, G.C. The remediation potential of Quercus nuttallii and Quercus phellos to heavy metal polluted soil in lead-zinc miningarea: A field test. Acta Ecol. Sin. 2020, 40, 6969–6981. [Google Scholar]
- Asad, S.A.; Farooq, M.; Afzal, A.; West, H. Integrated phytobial heavy metal remediation strategies for a sustainable clean environment—A review. Chemosphere 2019, 217, 925–941. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Chopra, P.; Chhillar, H.; Ahanger, M.A.; Hussain, S.J.; Maheshwari, C. Regulatory hubs and strategies for improving heavy metal tolerance in plants: Chemical messengers, omics and genetic engineering. Plant Physiol. Biochem. 2021, 164, 260–278. [Google Scholar] [CrossRef]
- Sharma, P.; Bhardwaj, R.; Arora, N.; Arora, H.K.; Kumar, A. Effects of 28-homobrassinolide on nickel uptake, protein content and antioxidative defense system in Brassica juncea. Biol. Plant 2008, 52, 767–770. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Rizvi, H.; Rinklebe, J.; Tsang, D.C.; Meers, E.; Ok, Y.S.; Ishaque, W. Phytomanagement of heavy metals in contaminated soils using sunflower: A review. Crit. Rev. Environ. Sci. Technol. 2016, 46, 1498–1528. [Google Scholar] [CrossRef]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of heavy metals by Alternanthera bettzickiana: Growth and physiological response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef]
- Parmar, P.; Patel, M.J.; Dave, B.; Subramanian, R.B. Nickel accumulation by Colocassia esculentum and its impact on plant growth and physiology. Afr. J. Agric. Res. 2012, 7, 3579–3587. [Google Scholar]
- Rizwan, M.; Ali, S.; Qayyum, M.F.; Ok, Y.S.; Rehman, M.Z.; Abbas, Z.; Hannan, F. Use of maize (Zea mays L.) for phytomanagement of Cd-contaminated soils: A critical review. Environ. Geochem. Health 2017, 39, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Lavid, N.; Schwartz, A.; Yarden, O.; Tel-Or, E. The involvement of polyphenols and peroxidase activities in heavy metal accumulation by epidermal glands of the waterlily (Nymphaeaceae). Planta 2012, 212, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Boulos, L. Flora of Egypt: Checklist, Revised Annotated Edition; Al-Hadara Publishing: Cairo, Egypt, 2009; Volume 410. [Google Scholar]
- Shaltout, K.; El-Kady, H.F.; El-Sheikh, M.A. Species diversity of the ruderal habitats in the Nile Delta. Taekholmia 1999, 19, 203–225. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Slima, D.F. The biology of Egyptian woody perennials. Assiut Univ. Ass. Uni. Bull. Environ. Res. 2007, 10, 85–103. [Google Scholar]
- Eltaher, G.T.; Ahmed, D.A.; EL-beheiry, M.; Sharaf El-Din, A. Biomass estimation and heavy metal accumulation by Pluchea dioscoridis (L.) DC. in the Middle Nile Delta, (Egypt): Perspectives for phytoremediation. S. Afr. J. Bot. 2019, 127, 153–166. [Google Scholar] [CrossRef]
- Nassar, M.; Elshamy, A.; El Gendy, A. Phenolics, essential oil and biological activity of Conyza dioscoridis growing in Egypt. Planta Med. 2014, 80, LP15. [Google Scholar] [CrossRef]
- El Shamy, A.; El Gendy, A.; Farrag, A.; Nassar, M. Antidiabetic and antioxidant activities of phenolic extracts of Conyza dioscoridis L. shoots. Int. J. Pharm. Pharm. Sci. 2015, 7, 65–72. [Google Scholar]
- El-Hefny, M.; Ashmawy, N.; Salem, M.; Salem, A. Antibacterial activities of the phytochemicals-characterized extracts of Callistemon viminalis, Eucalyptus camaldulensis and Conyza dioscoridis against the growth of some phytopathogenic bacteria. Microb. Pathog. 2017, 113, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Atta, A.H.; Mouneir, S.M. Antidiarrhoeal activity of some Egyptian medicinal plant extracts. J. Ethnopharm. 2004, 92, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Awaad, A.S.; EL-Meligy, R.M.; Qenawy, S.A.; Atta, A.H.; Soliman, G.A. Antiinflammatory, antinociceptive and antipyretic effects of some desert plants. J. Saudi Chem. Soc. 2011, 15, 367–373. [Google Scholar] [CrossRef]
- El-Ghorab, A.H.; Ramadan, M.M.; El-Moez, S.I.A.; Soliman, A.M.M. Essential oil, antioxidant, antimicrobial and anticancer activities of Egyptian Pluchea dioscoridis extract. Res. J. Pharmac. Bio. Chem. Sci. 2015, 6, 1255–1265. [Google Scholar]
- Kareim, M.S. Geomorphology of the east side of the River Nile. Bull. Egypt Geogr. Soc. 2001, 37, 295–358. (In Arabic) [Google Scholar]
- Sawidis, T.; Breuste, J.; Mitrovic, M.; Pavlovic, P.; Tsigaridas, K. Trees as bioindicators of heavy metal pollution in three European cities. Environ. Pollut. 2011, 159, 3560–3570. [Google Scholar] [CrossRef]
- Liu, L.; Guan, D.S.; Chen, Y.Q. Morphological structure of leaves and dust-retainingcapability of common street trees in Guangzhou Municipality. Acta Ecol. Sin. 2013, 33, 2604–2614. [Google Scholar] [CrossRef]
- Tremolieres, M.; Schnitzler, A.; Sanchez-Perez, J.M.; Schmitt, D. Changes in foliar nutrient content and resorption in Fraxinus excelsior L., Ulmus minor Mill. and Clematis vitalba L. after prevention of floods. Ann. Sci. 1999, 56, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K. Practical Plant Physiology; Kalayani Publishers: New Delhi, India, 1977; p. 226. [Google Scholar]
- Boham, B.A.; Kocipai-Abyazan, R. Flavonoids and condensed tannins from leaves of Hawaiian Vaccinium and V. calycinium. Pacific Sci. 1974, 48, 458–463. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307. [Google Scholar] [CrossRef]
- Harborne, J.B. Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis; Chapman A & Hall: London, UK, 1973; p. 279. [Google Scholar]
- Williams, I.; Frank, L. Improved chemical synthesis and enzymatic assay of δ1-pyrroline-5-carboxylic acid. Anal. Biochem. 1975, 64, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substrates. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Jones, J. Laboratory Guide for Conducting Soil Tests and Plant Analysis; CRC Press; Taylor & Francis: Boca Raton, FL, USA, 2001. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: London, UK, 1989. [Google Scholar]
- U.S. EPA. National Ambient Air Quality Standards (NAAQS); U.S. EPA: Washington, DC, USA, 2016. Available online: http://www3.epa.gov/ttn/naaqs/ (accessed on 2 December 2016).
- Liang, J.; Fang, H.; Zhang, T.; Wang, X.; Liu, Y. Heavy metal in leaves of twelve plant species from seven different areas in Shanghai, China. Urban For. Urban Green. 2017, 27, 390–398. [Google Scholar] [CrossRef]
- Guarino, C.; Sciarrillo, R. Ecological and functional evaluation of species candidate for heavy metals phytoremediation in SIN porto torres (Sardinia, Italy). J. Environ. Anal. Toxicol. 2017, 7, 484. [Google Scholar] [CrossRef]
- Luo, Y.; Wu, Y.; Qiu, J.; Wang, H.; Yang, L. Suitability of four woody plant species for the phytostabilization of a zinc smelting slag site after 5 years of assisted revegetation. J. Soils Sediments 2019, 19, 702–715. [Google Scholar] [CrossRef]
- Mohotti, A.; Geeganage, K.; Mohotti, K.; Ariyarathne, M.; Karunaratne, C.; Chandrajith, R. Phytoremedial potentials of Ipomoea aquatica and Colocasia esculenta in soils contaminated with heavy metals through automobile painting, repairing and service centers. Sri Lankan J. Biol. 2016, 1, 27–37. [Google Scholar] [CrossRef]
- Cheraghi, M.; Lorestani, B.; Khorasani, N.; Yousefi, N.; Karami, M. Findings on the phytoextraction and phytostabilization of soils contaminated with heavy metals. Biol. Trace Elem. Res. 2011, 144, 1133–1141. [Google Scholar] [CrossRef]
- Tiwari, S.; Agrawal, M.; Marshall, F.M. Evaluation of ambient air pollution impact on carrot plants at a sub urban site using open top chambers. Environ. Monit. Assess. 2006, 119, 15–30. [Google Scholar] [CrossRef]
- Seyyednejad, S.M.; Niknejad, M.; Koochak, H. A review of some different effects of air pollution on plants. Res. J. Environ. Sci. 2011, 10, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Barden, J.A. Apple tree growth, et photosynthesis, dark respiration and specific leaf weight as affected by continuous and intermittent shade. J. Am. Soc. Hortic. Sci. 1977, 102, 191–194. [Google Scholar]
- Rai, V.; Vajpayee, P.; Singh, S.N.; Mehrotra, S. Effect of chromium accumulation on photosynthetic pigments, oxidative stress defense system, nitrate reduction, proline level and eugenol content of Ocimum tenuiflorum L. Plant Sci. 2004, 167, 1159–1169. [Google Scholar] [CrossRef]
- Sandelius, A.; Naslund, K.; Carlsson, A.; Pleijel, H.; Sellden, G. Exposure of spring wheat (Triticum aestivum) to ozone in open-top chambers. Effects on acyl lipid composition and chlorophyll content of flag leaves. New Phytol. 1995, 131, 231–239. [Google Scholar] [CrossRef]
- Diatta, J.; Youssef, N.; Tylman, O.; Grzebisz, W.; Markert, B.; Drobek, L.; Wünschmann, S.; Bebek, M.; Mitko, K.; Lejwoda, P. Acid rain induced leakage of Ca, Mg, Zn, Fe from plant photosynthetic organs—Testing for deciduous and dicotyledons. Ecol. Indic. 2020, 121, 107210. [Google Scholar] [CrossRef]
- Yoon, Y.; Cao, X.; Zhou, Q.; Ma, L.Q. Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef]
- Mahar, A.; Wang, P.; Ali, A.; Awasthi, M.K.; Lahori, A.H.; Wang, Q.; Li, R.; Zhang, Z. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: A review. Ecotoxicol. Environ. Saf. 2016, 126, 111–121. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Rizwan, M.; Hussain, A.; Saqib, M.; Ali, S.; Sohail, M.I.; Shafiq, M.; Hafeez, F. Alleviation of cadmium (Cd) toxicity and minimizing its uptake in wheat (Triticum aestivum) by using organic carbon sources in Cd-spiked soil. Environ. Pollut. 2018, 241, 557–565. [Google Scholar] [CrossRef]
- De Filippis, L.F.; Pallaghy, C.K. Heavy metals: Sources and biological effects. In Algae and Water Pollution: Advances in Limnology Series; Rai, L.C., Caur, J.P., Soeder, C.J., Eds.; Schweizerbart: Stuttgart, Germany, 1994; Volume 42, pp. 32–77. [Google Scholar]
- Delfine, S.; Alvino, A.; Villana, M.C.; Loreto, F. Restriction to carbon dioxide and photosynthesis in spinach leaves recovering from salt stress. Plant Physiol. 1999, 199, 1101–1106. [Google Scholar] [CrossRef]
- Monni, S.; Uhlig, C.; Hansen, E.; Magel, E. Ecophysiological responses of Empetrum nigrum to heavy metall pollution. Environ. Pollut. 2001, 112, 121–129. [Google Scholar] [CrossRef]
- Anderson, M.J.; Evans, P.K. The chlorophyll a/b-proteins of PS I and PS II are immunologically related. FEBS Lett. 1986, 199, 227–233. [Google Scholar] [CrossRef]
- Hamidi-Moghaddam, A.; Arouiee, H.; Moshtaghi, N.; Azizi, M.; Shoor, M.; Sefidkon, F. Visual quality and morphological responses of rosemary plants to UV-B radiation and salinity stress. J. Ecol. Eng. 2019, 20, 34–43. [Google Scholar] [CrossRef]
- McElroy, J.S.; Kopsell, D.A. Physiological role of carotenoids and other antioxidants in plants and application to turfgrass stress management. N. Z. J. Crop Hortic. Sci. 2009, 37, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Polle, A.; Chakrabarti, K.; Chakrabarti, S.; Seifert, F.; Schramel, P.; Rennenberg, H. Antioxidants and manganese deficiency in needles of Norway Spruce (Picea abies L.) trees. Plant Physiol. 1992, 99, 1084–1089. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Harbinson, J. Oxygen metabolism and the regulation of photosynthetic electron transport CH Foyer. In Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants; Mullineaux, P., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1994; pp. 1–42. [Google Scholar]
- Flowers, T.J.; Troke, P.F.; Yeo, A.R. The mechanism of salt tolerance in halophites. Annu. Rev. Plant Physiol. 1977, 28, 89–121. [Google Scholar] [CrossRef]
- Shvaleva, A.L.; Costa, F.; Silva, E.; Breia, E.; Jouve, L.; Hausman, J.F.; Almeida, M.H.; Maroco, J.P.; Rodrigues, M.L.; Pereira, J.S.; et al. Metabolic responses to water deficit in two Eucalyptus globules clones with contrasting drought sensitivity. Tree Physiol. 2006, 26, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naya, D.E.; Lardies, M.A.; Bozinovik, F. The effect of diet quality on physiological and life-history traits in the harvestman Pachylus paessleri. New Phytol. 2007, 131, 231–239. [Google Scholar] [CrossRef]
- Kupper, H.; Kupper, F.; Spiller, M. In situ detection of heavy metals substituted chlorophylls in water plants. Photosynth. Res. 1998, 58, 123–133. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Craker, L.E.; Xing, B. Effects of Cd, Pb, and Cu on growth and essential oil contents in dill, peppermint, and basil. Environ. Exp. Bot. 2006, 58, 9–16. [Google Scholar] [CrossRef]
- Shahid, M. Biogeochemical Behavior of Heavy Metals in Soil-Plant System, 1st ed.; 1. Soil and Environmental Science; Higher Education Commission: Islamabad, Pakistan, 2017; 577.14–dc23; ISBN 978-969-417-195-1. [Google Scholar]
- Hatamian, M.; Nejad, A.R.; Kafi, M.; Souri, M.K.; Shahbazi, K. Growth characteristics of ornamental Judas tree (Cercis siliquastrum L.) seedling under different concentrations of lead and cadmium in irrigation water. Acta Sci. Polonorum Hort Cultus 2019, 18, 87–96. [Google Scholar] [CrossRef]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Khan, A.H.A.; Nawaz, I.; Qu, Z.; Butt, T.A.; Yousaf, S.; Iqbal, M. Reduced growth response of ornamental plant Nicotiana alata L. uponselected heavy metals uptake, with co-application of ethylenediaminetetraacetic acid. Chemosphere 2020, 241, 125006. [Google Scholar] [CrossRef]
- Sinha, S.; Pandey, K.; Gupta, A.K.; Bhatt, K. Accumulation of metals in vegetables and crops grown in the area irrigated with river water. Bull. Environ. Contam. Toxicol. 2005, 74, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Sivaci, A.; Elmas, E. The combined effects of cadmium and salinity on some pigments and total phenolic compounds of Myriophyllum heterophyllum Michx. and Potamogeton crispus L. Afr. J. Agric. Res. 2012, 7, 3813–3818. [Google Scholar]
- Park, J.H.; Lamb, D.; Paneerselvam, P.; Choppala, G.; Bolan, N. Role of organic amendments on enhanced bioremediation of heavy metal (loid) contaminated soils. J. Hazard. Mater. 2011, 185, 549–574. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Rehman, M.Z.; Maqbool, A. A critical review on the effects of zinc at toxic levels of cadmium in plants. Environ. Sci. Pollut. Res. 2019, 26, 6279–6280. [Google Scholar] [CrossRef]
- Rosselli, W.; Keller, C.; Boschi, K. Phytoextraction capacity of trees growing on metal contaminated soil. Plant Soil 2003, 256, 265–272. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site | Ind. No | Growth Parameters | Biomass Estimation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Stem | Flower | Root | |||||||||
| SLA (cm2/g d.w.) | RWC (%) | LA (cm2) | Fw (g) | Dw (g) | Fw (g) | Dw (g) | Fw (g) | Dw (g) | Fw (g) | Dw (g) | ||
| S1 | 1 | 75.91 | 149.03 | 4.51 | 7.56 | 6.71 | 21.61 | 18.81 | 2.32 | 2.11 | 13.9 | 12.1 |
| 2 | 51.76 | 156.24 | 3.5 | 7.03 | 5.91 | 21.73 | 19.97 | 3.65 | 2.52 | 12.4 | 11.8 | |
| 3 | 67.93 | 135.32 | 3.55 | 6.31 | 5.36 | 20.66 | 18.88 | 2.77 | 1.31 | 11.9 | 10.0 | |
| 4 | 71.35 | 130.66 | 4.59 | 7.84 | 5.4 | 21.20 | 18.93 | 3.93 | 2.42 | 13.7 | 11.82 | |
| 5 | 78.34 | 122.76 | 3.47 | 6.18 | 4.67 | 19.59 | 17.83 | 3.85 | 2.89 | 11.04 | 9.7 | |
| S2 | 6 | 74.18 | 190.1 | 3.51 | 8.96 | 7.02 | 23.68 | 20.51 | 3.98 | 2.47 | 12.9 | 10.1 |
| 7 | 89.54 | 160.87 | 4.42 | 7.91 | 6.39 | 23.72 | 20.17 | 3.99 | 2.23 | 13.4 | 11.7 | |
| 8 | 67.09 | 154.87 | 3.3 | 7.94 | 6.10 | 27.88 | 25.39 | 3.92 | 2.89 | 12.6 | 10.9 | |
| 9 | 75.81 | 140.64 | 4.6 | 8.66 | 7.79 | 21.41 | 19.09 | 3.67 | 2.29 | 14.1 | 12.5 | |
| 10 | 61.40 | 180.54 | 3.32 | 7.90 | 6.82 | 22.79 | 20.19 | 4.87 | 3.43 | 15.8 | 14.13 | |
| S3 | 11 | 86.16 | 206.81 | 6.11 | 10.08 | 9.7 | 27.67 | 25.75 | 3.62 | 3.01 | 19.1 | 16.0 |
| 12 | 101.14 | 276.81 | 5.44 | 9.17 | 8.08 | 30.36 | 28.59 | 4.68 | 3.17 | 17.3 | 15.1 | |
| 13 | 84.83 | 200.61 | 5.18 | 9.96 | 7.78 | 28.57 | 26.19 | 5.45 | 4.45 | 19.3 | 16.4 | |
| 14 | 97.97 | 249.91 | 6.42 | 10.92 | 9.16 | 27.45 | 25.77 | 4.12 | 3.61 | 16.9 | 13.0 | |
| 15 | 83.98 | 211.01 | 5.52 | 9.88 | 8.44 | 28.86 | 25.98 | 4.49 | 3.61 | 16.3 | 14.9 | |
| Accumulation | Degree of Freedom | FW. of Leaf (g) | DW. of Leaf (g) | FW. of Stem (g) | DW. of Stem (g) | FW. of Root (g) | DW. of Root (g) | FW. of Flower (g) | DW. of Flower (g) | Leaf Area (LA) (cm2) | SLA (cm2/g d.w) | Leaf WC (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Replication | 3 | 119.216 | 67.614 | 195.68 | 122.298 | 171.83 | 95.719 | 83.47 | 57.614 | 219.027 | 152.671 | 221.79 |
| Accumulation | 9 | 35. 45 | 28.015 | 23.0465 | 2.154 | 20.211 | 15.474 | 17.298 | 18.015 | 56.284 | 26.819 | 30.02 |
| Correlation (r-values) | 20 | 9.123 ** | 7.512 ** | 5.285 ** | 0.363 * | 7.131 ** | 7.561 ** | 6.65 * | 6.702 ** | 10.695 ** | 9.871 | 5.022 ** |
| Coefficient variation (%) | 9.83 | 9.73 | 9.09 | 7.14 | 7.38 | 11.55 | 9.93 | 6.67 | 9.39 | 9.89 | 5.64 | 6.31 |
| Biochemical Parameters | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chl a mg/g f.w | Chl b mg/g f.w | Car. mg/g f.w | Chl. a + b mg/g f.w | Chl. a/b Ratio | Car/Chl mg/g f.w | Alkaloides % | Flavonoids mg/g f.w | Proline (mg/g d.w) | Soluble Carbohydrates (mg/g d.w) | |
| S1 | 1.38 ± 0.01 b | 1.55 ± 0.02 b | 1.41 ± 0.03 b | 2.93 ± 0.51 a | 0.89 ± 0.01 a | 0.48 ± 0.03 a,b | 5 ± 0.24 b | 22.30 ± 2.438 d | 0.52 ± 0.16 a | 90.35 ± 6.9 d |
| S2 | 1.99 ± 0.13 b | 1.78 ± 0.02 b | 1.30 ± 0.04 a | 3.77 ± 0.93 a | 1.11 ± 0.02 a | 0.34 ± 0.02 b | 4.84 ± 0.95 b | 15.39 ± 1.40 c | 0.78 ± 0.11 b | 82.43 ± 9.08 c |
| S3 | 2.87 ± 0.03 a | 2.81 ± 0.02 c | 1.28 ± 0.02 a | 5.68 ± 0.98 b | 1.02 ± 0.03 b | 0.22 ± 0.01 a | 1.84 ± 0.67 a | 18.98 ± 2.322 a | 0.34 ± 0.013 b | 113.10 ± 9.1 b |
| F-value | 11.414 | 5.893 | 5.014 | 8.976 | 5.103 | 13.990 | 2.01 | 55.911 | 6.193 | 9.414 |
| Site | Plant Organ | NPK and TEs (mg/kg) in Plant | |||||||
|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Cu | Pb | Cd | Zn | Mn | ||
| S1 | Root | 2.241 ± 0.231 b,c | 0.669 ± 0.037 a | 0.779 ± 0.053 a | 160.2 ± 29.27 b | 303.20 ± 51.79 b | 86.74 ± 8.48 a | 125.85 ± 38.60 d | 295.0 ± 13.38 b |
| Shoot | 3.24 ± 0.091 b | 0.914 ± 0.061 a | 0.981 ± 0.061 a | 154.87 ± 18.14 b | 329.0 ± 25.83 b,c | 95.65 ± 9.90 a | 118.68 ± 28.54 d | 291.80 ± 17.75 d | |
| S2 | Root | 2.000 ± 0.067 a | 0.206 ± 0.032 a | 0.467 ± 0.036 a | 188.75 ± 16.3 b,c | 201.90 ± 22.94 a,b | 65.56 ± 16.27 b | 111.43 ± 31.56 b | 127.596 ± 11.79 b |
| Shoot | 2.50 ± 0.088 a,b | 0.444 ± 0.018 a | 0.378 ± 0.040 b | 174.0 ± 16.12 a | 211.0 ± 32.92 b | 79.38 ± 11.36 a,b | 113.85 ± 14.32 a | 120.015 ± 12.02 b | |
| S3 | Root | 1.850 ± 0.307 c | 0.367 ± 0.035 b | 0.529 ± 0.069 a | 72.5 ± 12.16 a,b | 60.4 ± 14.06 a,b | 35.60 ± 18.36 c,d | 91.30 ± 15.23 a | 31.367 ± 11.81 b,c |
| Shoot | 2.65 ± 0.095 b,c | 0.438 ± 0.044 d | 0.494 ± 0.027 b,c | 60.7 ± 12.08 b,c | 81.60 ± 11.76 b,c | 46.09 ± 7.65 b,c | 87.70 ± 6.41 a | 21.558 ± 1.06 b | |
| Site | N | P | K | Cu | Pb | Cd | Zn | Mn | pH | OM (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 0.213 ± 0.006 a | 0.0413 ± 0.0035 c | 0.173 ± 0.0069 a | 227.29 ± 21.57 d | 223.0 ± 32.07 c | 81.67 ± 12.05 c | 184.56 ± 9.16 d | 94.24 ± 14.49 c | 8.29 ± 0.22 a | 6.30 ± 0.35 c |
| S2 | 0.190 ± 0.005 b | 0.0353 ± 0.0019 b | 0.314 ± 0.0086 d | 225.11 ± 33.73 d | 222.3 ± 21.08 a | 96.54 ± 11.03 b | 163.02 ± 11.09 c | 89.04 ± 9.31 b | 7.69 ± 0.28 b | 2.60 ± 0.16 a |
| S3 | 0.089 ± 0.004 a | 0.0277 ± 0.0029 a | 0.070 ± 0.0089 b | 97.80 ± 32.18 c | 82.0 ± 11.07 a | 21.35 ± 5.05 a | 133.00 ± 7.05 c | 38.74 ± 6.30 a | 7.28 ± 0.18 a | 3.20 ± 0.25 a |
| TEs | Parameters | Site | F-Value | ||
|---|---|---|---|---|---|
| S1 | S2 | S3 | |||
| Cu | CF | 1.38 ± 0.15 a | 1.60 ± 0.45 a | 1.36 ± 0.55 a | 142.123 |
| TF | 0.96 ± 0.08 a | 0.92 ± 0.091 a | 0.83 ± 0.048 a | 237.387 | |
| ECR | 0.70 ± 0.043 a | 0.83 ± 0.093 a | 0.74 ± 0.074b c | 167.284 | |
| ECS | 0.68 ± 0.056 b,c | 0.77 ± 0.081 a | 0.62 ± 0.056 a | 132.211 | |
| Pb | CF | 2.83 ± 0.85 a,b | 2.26 ± 0.95b c | 1.73 ± 0.65b c | 45.46 |
| TF | 0.92 ± 0.105 b | 0.67 ± 0.046 a | 1.35 ± 0.35 c | 10.34 | |
| ECR | 1.47 ± 0.65 a,b | 1.35 ± 0.351 a,b | 0.73 ± 0.09 a | 22.13 | |
| ECS | 1.35 ± 0.35 a,b | 0.90 ± 0.054 a | 0.99 ± 0.084 b,c | 31.10 | |
| Cd | CF | 2.23 ± 0.975 b | 1.50 ± 0.75 b | 3.82 ± 1.05 b | 42.52 |
| TF | 0.90 ± 0.077 a,b | 0.82 ± 0.087 a | 1.29 ± 0.04 a,b | 6.77 | |
| ECR | 1.17 ± 0.095 c | 0.82 ± 0.15 b,c | 1.66 ± 0.066 c | 38.16 | |
| ECS | 1.06 ± 0.097 b | 0.67 ± 0.051 b | 2.15 ± 0.65 b | 32.87 | |
| Zn | CF | 1.32 ± 0.099 a | 1.43 ± 0.068 a | 1.34 ± 0.35 b | 378.94 |
| TF | 0.94 ± 0.106 a | 0.97 ± 0.093 a | 0.96 ± 0.15 a,b | 150.77 | |
| ECR | 0.68 ± 0.021 a | 0.73 ± 0.059 a,b | 0.68 ± 0.07 b | 371.46 | |
| ECS | 0.64 ± 0.053 a | 0.69 ± 0.057 a | 0.65 ± 0.043 b | 184.33 | |
| Mn | CF | 6.22 ± 1.85 d | 2.78 ± 0.965 b | 1.36 ± 0.145 a | 386.59 |
| TF | 0.98 ± 0.05 b | 0.94 ± 0.081 a,b | 0.68 ± 0.07 a | 32.76 | |
| ECR | 3.13 ± 0.98 c | 1.43 ± 0.094 c,d | 0.80 ± 0.105 a | 99.91 | |
| ECS | 3.09 ± 0.97 c | 1.34 ± 0.079 b | 0.55 ± 0.008 b,c | 363.10 | |
| Plant NPK and TEs | Soil NPK and TEs | Soil pH | Soil OM | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Cu | Pb | Cd | Zn | Mn | |||
| Root | 0.31 | 0.52 | 0.45 | 0.05 | 0.01 | 0.11 | −0.73 | 0.03 | 0.51 * | 0.88 *** |
| N | 0.05 | −0.23 | 0.11 | 0.19 | 0.59 | 0.72 | 0.62 | −0.71 | 0.32 | 0.76 ** |
| P | 0.30 | 0.51 | −0.65 | −0.66 ** | 0.01 | 0.31 | 0.41 | 0.55 | 0.79 | 0.71 ** |
| K | 0.26 | 0.77 ** | −0.81 ** | 0.49 | −0.71 * | −0.23 | −0.41 | −0.24 | 0.22 | 0.60 ** |
| Cu | 0.48 | −0.42 | 0.72 | 0.71 | −0.19 | 0.83 * | 0.52 * | −0.38 * | −0.58 ** | 0.51 * |
| Pb | −0.11 | 0.22 | 0.34 | −0.71 | 0.29 | 0.21 | −0.31 | 0.41 | −0.51 * | 0.86 *** |
| Cd | −0.62 | 0.51 | −0.42 | 0.13 | −0.59 | 0.77 ** | 0.01 | 0.76 ** | −0.77 ** | 0.97 *** |
| Zn | −0.49 | 0.53 | −0.38 | 0.89 *** | 0.38 | 0.65 * | 0.23 | 0.71 * | −0.69 *** | 0.88 *** |
| Mn | −0.23 | −0.73 ** | 0.71 * | −0.59 | 0.58 | −0.51 * | −0.03 | 0.91 *** | −0.49 ** | 0.53 * |
| Shoot | ||||||||||
| N | 0.03 | 0.71 ** | −0.45 * | −0.43 | −0.41 | 0.62 | 0.32 | −0.31 * | 0.33 | 0.51 * |
| P | 0.59 * | 0.02 | 0.33 | −0.49 | −0.29 | 0.32 | 0.61 | 0.41 * | 0.66 * | 0.68 ** |
| K | 0.68 * | 0.50 | 0.73 | −0.31 * | 0.05 | −0.41 | −0.61 | −0.22 | 0.21 | 0.69 ** |
| Cu | −0.31 | −0.58 * | 0.71 | 0.36 | 0.72 | 0.63 | 0.49 | −0.03 | 0.07 | 0.91 *** |
| Pb | −0.07 | 0.43 | 0.29 | 0.50 | 0.61 | 0.11 | 0.71 | −0.91 | −0.72 * | 0.95 *** |
| Cd | 0.43 | 0.01 | −0.41 | 0.84 | 0.35 | 0.87 ** | 0.88 | −0.22 * | −0.51 * | 0.69 ** |
| Zn | −0.41 | 0.42 | 0.24 | 0.61 | 0.50 | 0.41 | 0.07 | 0.62 | −0.71 * | 0.75 ** |
| Mn | −0.03 | −0.03 | −0.65 * | −0.60 | −0.21 | 0.61 * | 0.01 | 0.86 *** | −0.64 ** | 0.61 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, N.; Diatta, J. Potential Efficiency of Wild Plant Species (Pluchea dioscoridis (L.) DC.) for Phytoremediation of Trace Elements on Contaminated Locations. Sustainability 2023, 15, 119. https://doi.org/10.3390/su15010119
Youssef N, Diatta J. Potential Efficiency of Wild Plant Species (Pluchea dioscoridis (L.) DC.) for Phytoremediation of Trace Elements on Contaminated Locations. Sustainability. 2023; 15(1):119. https://doi.org/10.3390/su15010119
Chicago/Turabian StyleYoussef, Naglaa, and Jean Diatta. 2023. "Potential Efficiency of Wild Plant Species (Pluchea dioscoridis (L.) DC.) for Phytoremediation of Trace Elements on Contaminated Locations" Sustainability 15, no. 1: 119. https://doi.org/10.3390/su15010119