
Sustainable Exploitation of Greek Rosmarinus officinalis L. Populations for Ornamental Use through Propagation by Shoot Cuttings and In Vitro Cultures

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Source
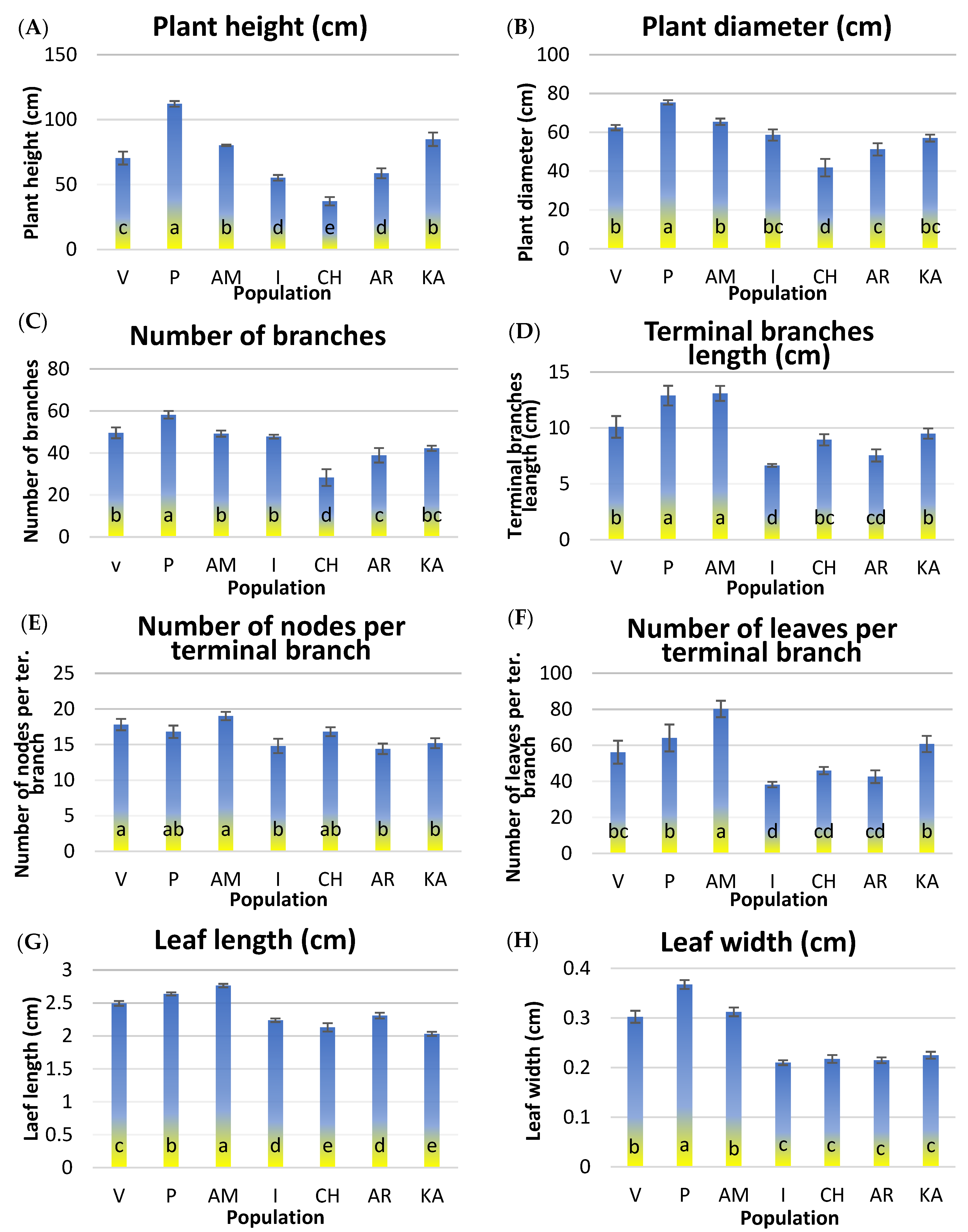
2.2. Morphological Traits
2.3. Propagation by Shoot Cuttings
2.3.1. Plant Material
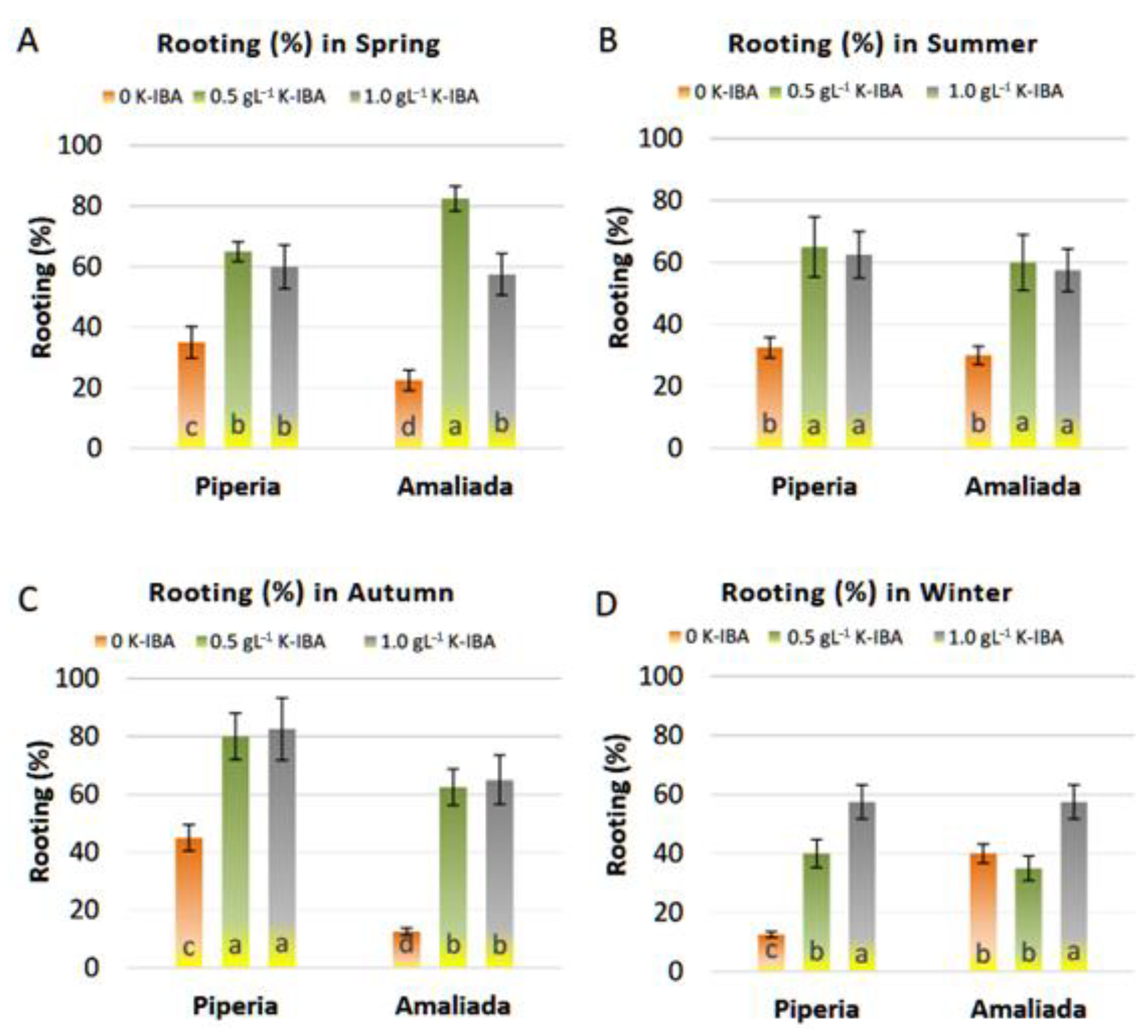
2.3.2. Effect of K-IBA and Season on Rooting of Cuttings
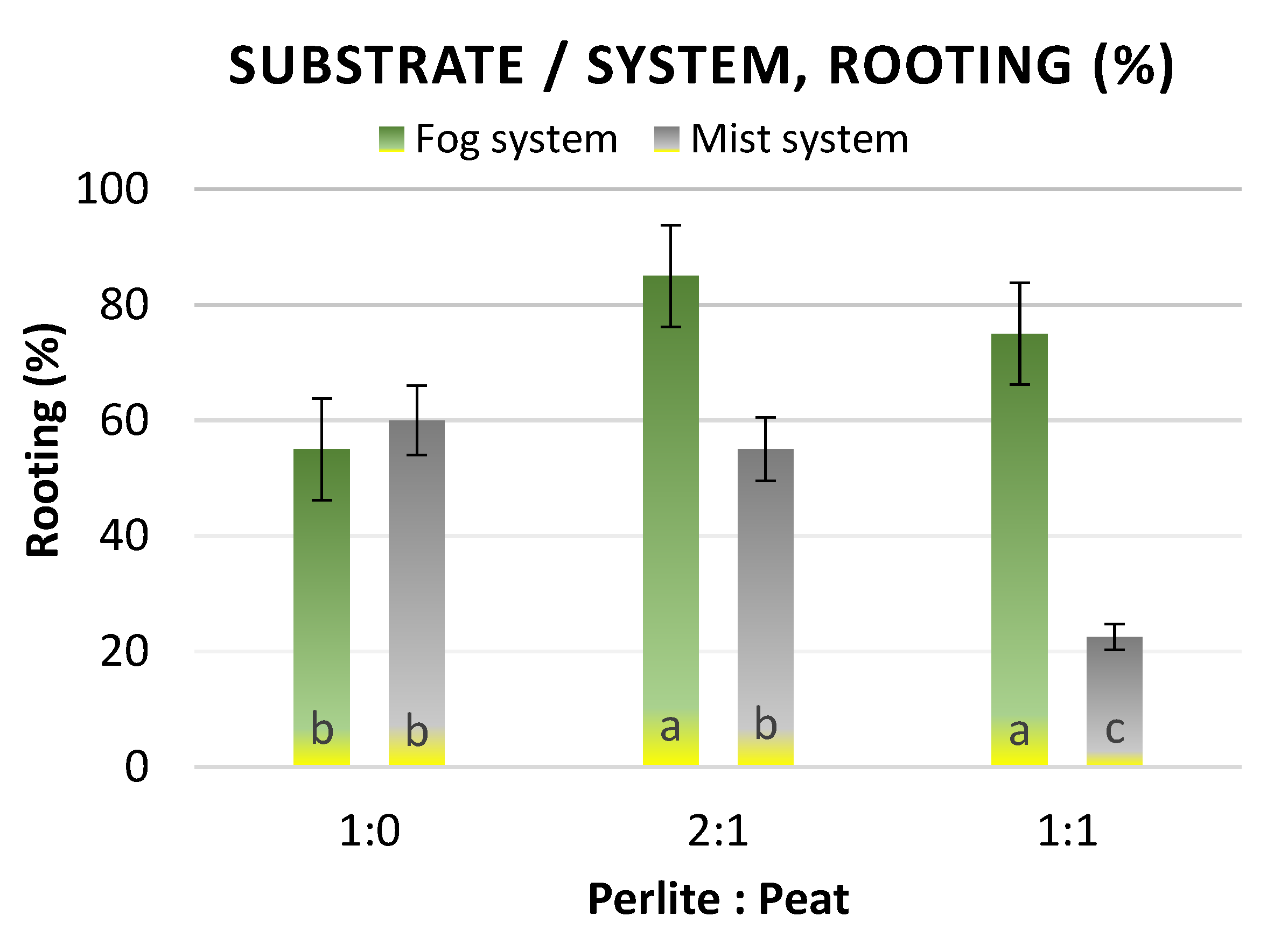
2.3.3. Effect of Substrate and Rooting System on Rooting of Cuttings
2.3.4. Statistical Analysis
2.4. Propagation In Vitro
2.4.1. Plant Material
2.4.2. Explant Disinfestation and Establishment of In Vitro Cultures
2.4.3. Effect of Growth Regulators on Shoot Multiplication and Rooting
2.4.4. Plantlet Acclimatization
2.5. Statistical Analysis
3. Results and Discussion
3.1. Morphological Analysis of the Traits
3.2. Propagation by Shoot Cuttings
3.2.1. Effect of K-IBA and Season on Rooting of Cuttings
3.2.2. Effect of Substrate and Rooting System on Rooting of Cuttings
3.3. Propagation In Vitro
3.3.1. Explant Disinfestation and Establishment of In Vitro Cultures
3.3.2. Effect of Plant Growth Regulators on Shoot Multiplication and Rooting
3.3.3. Plantlet Acclimatization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- González-Minero, F.J.; Bravo-Díaz, L.; Ayala-Gómez, A. Rosmarinus officinalis L. (Rosemary): An ancient plant with uses in personal healthcare and cosmetics. Cosmetics 2020, 7, 77. [Google Scholar] [CrossRef]
- Hammer, M.; Junghanns, W. Rosmarinus officinalis L.: Rosemary. In Medicinal, Aromatic and Stimulant Plants, 1st ed.; Novak, J., Blüthner, W.-D., Eds.; Springer: Cham, Switzerland, 2020; Volume 12, pp. 501–521. [Google Scholar] [CrossRef]
- Kiuru, P.; Muriuki, S.J.N.; Wepukhulu, S.B.; Muriuki, S.J.M. Influence of growth media and regulators on vegetative propagation of rosemary (Rosmarinus officinalis L.). East Afr. Agric. For. J. 2015, 81, 105–111. [Google Scholar] [CrossRef]
- Mehrabani, V.L.; Kamran, R.V.; Hassanpouraghdam, M.B.; Kavousi, E.; Aazami, M.A. Auxin concentration and sampling time affect rooting of Chrysanthemum morifolium L. and Rosmarinus officinalis L. Azar. J. Agric. 2016, 3, 11–16. Available online: http://azarianjournals.ir/wp-content/uploads/aja15121402.pdf (accessed on 2 February 2022).
- German, T.; Mekonnen, M.; Mengesha, M.; Philiphos, M. Rosemary Production and Utilization, 1st ed.; Ethiopian Institute of Agricultural Research: Addis Ababa, Ethiopia, 2003; p. 27. Available online: https://www.researchgate.net/publication/313030290_Cultivation_Processing_and_Utilization_of_Rosemary_Rosemarinus_officinalis_L (accessed on 15 February 2022).
- Andrade, J.M.; Faustino, C.; Garcia, C.; Ladeiras, D.; Reis, C.P.; Rijo, P. Rosmarinus officinalis L.: An update review of its phytochemistry and biological activity. Future Sci. OA 2018, 4, FSO283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrubba, A.; La Torre, R.; Piccaglia, R.; Grandi, S. Chemical and botanical characterization of a Rosmarinus officinalis biotype from Sicily. Acta Hortic. 2006, 723, 197–202. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Knapen, A.; Barberá, G.G.; Navarro, J.A. Root characteristics of representative Mediterranean plant species and their erosion-reducing potential during concentrated runoff. Plant Soil 2007, 294, 169–183. [Google Scholar] [CrossRef]
- De Mastro, G.; Ruta, C.; Mincione, A.; Poiana, M. Bio-morphological and chemical characterization of rosemary (Rosmarinus officinalis L.) biotypes. Acta Hortic. 2004, 629, 471–482. [Google Scholar] [CrossRef]
- Lagouri, V.; Alexandri, G. Antioxidant properties of Greek O. dictamnus and R. officinalis methanol—HPLC determination of phenolic acids. Int. J. Food Prop. 2013, 16, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Malvezzi, L.; Mendes, E.; Militao, L.; Tundisi, L.L.; Ataide, J.A.; Souto, E.B.; Mazzola, P.G. Rosemary (Rosmarinus officinalis L., syn Salvia rosmarinus Spenn.) and its topical applications: A review. Plants 2020, 9, 651. [Google Scholar] [CrossRef]
- Nunziata, A.; Cervelli, C.; De Benedetti, L. Genotype confidence percentage of SSR HRM profiles as a measure of genetic similarity in Rosmarinus officinalis. Plant Gene 2018, 14, 64–68. [Google Scholar] [CrossRef]
- Ganopoulos, I.V.; Kazantzis, K.; Chatzicharisis, I.; Karayiannis, I.; Tsaftaris, A.S. Genetic diversity, structure and fruit trait associations in Greek sweet cherry cultivars using microsatellite based (SSR/ISSR) and morpho-physiological markers. Euphytica 2011, 181, 237–251. [Google Scholar] [CrossRef]
- Sarrou, E.; Martens, S.; Chatzopoulou, P. Metabolite profiling and antioxidative activity of Sage (Salvia fruticosa Mill.) under the influence of genotype and harvesting period. Ind. Crops Prod. 2016, 94, 240–250. [Google Scholar] [CrossRef]
- Motta, E.V.; Sampaio, B.L.; Costa, J.C.; Teixeira, S.P.; Bastos, J.K. Quantitative analysis of phenolic metabolites in Copaifera langsdorffii leaves from plants of different geographic origins cultivated under the same environmental conditions. Phytochem. Anal. 2019, 30, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Paradiković, N.; Zeljković, S.; Tkalec, M.; Vinković, T.; Dervić, I.; Marić, M. Influence of rooting powder on propagation of sage (Salvia officinalis) and rosemary (Rormarinus officinalis) with green cuttings. Poljoprivreda 2013, 19, 10–15. [Google Scholar]
- Elhaak, M.A.; Matter, M.Z.; Zayed, M.A.; Gad, D.A. Propagation principles in using Indole-3-butyric acid for rooting rosemary stem cuttings. J. Hortic. 2015, 2, 121. [Google Scholar] [CrossRef]
- Talia, M.A.C.; Viola, F.; Forleo, L.R. Vegetative propagation of two species of the Mediterranean maquis (Rosmarinus officinalis L., Viburnum tinus L.) for applications in naturalistic engineering. Italus Hortus 2004, 11, 89–92. [Google Scholar]
- Abu-Zahra, T.R.; Al-Shadaideh, A.N.; Abubaker, S.; Qrunfleh, I. Influence of auxin concentrations on different ornamental plants. Int. J. Bot. 2013, 9, 96–99. [Google Scholar] [CrossRef]
- Koleva-Gudeva, L.; Trajkova, F.; Mihajlov, L.; Troiciki, J. Influence of different auxins on rooting of rosemary, sage and elderberry. Annu. Res. Rev. Biol. 2017, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lasheen, F.F.; Hewidy, M.; Emam, O.N.K. Impact of rooting promoters (PHOS ROOT-X) with or without using IBA on rooting of terminal and middle cuttings of rosemary (Rosmarinus officinalis L.). Scient. J. Flowers Ornam. Plants 2021, 8, 153–163. [Google Scholar] [CrossRef]
- Shahhoseini, R.; Moghaddam, M.; Kiani, D.; Mansori, R. Effect of different concentrations of IBA and NAA on rooting of semi-hardwood cuttings of rosemary (Rosmarinus officinalis L.). Iran. J. Med. Arom. Plants 2015, 31, 574–585. [Google Scholar]
- Davies, F.; Geneve, R.; Wilson, S.; Hartmann, H.; Kester, D. Hartmann and Kester’s Plant Propagation: Principles and Practices, 9th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2017; p. 915. [Google Scholar]
- Varban, D.; Varban, R.; Ghete, A.; Odagiu, A.; Czako, R. The influence of some products on the rooting of Rosmarinus officinalis L. cuttings. Hop. Med. Plants 2018, 26, 94–100. [Google Scholar]
- Mendoza-Hernandez, D.; Fornes, F.; Belda, R. Compost and vermicompost of horticultural waste as substrates for cutting rooting and growth of rosemary. Sci. Hortic. 2014, 178, 192–202. [Google Scholar] [CrossRef]
- Silva, C.; Pedras, J. Early rooting in rosemary’s (Rosmarinus officinalis L.) cuttings under the influence of chemical treatments and collecting time. Acta Hortic. 1999, 502, 34. [Google Scholar] [CrossRef]
- Souza, D.M.S.C.; Fernandes, S.B.; Molinari, L.V.; Avelar, M.L.M.; Brondani, G.E. Activated charcoal application for the micropropagation of Cattleya crispata (Thunb.) Van den Berg. Nativa 2021, 9, 352–358. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Tsoktouridis, G.; Maloupa, E. In vitro propagation of medicinal and aromatic plants: The case of selected Greek species with conservation priority. Vitr. Cell. Dev. Biol. Plant 2019, 55, 635–646. [Google Scholar] [CrossRef]
- Chaturvedi, H.C.; Misra, P.; Sharma, M. In vitro multiplication of Rosmarinus officinalis L. Zeitschr. Pflanzenphys. 1984, 113, 301–304. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, R.; Li, Z.; Qi, C.; Liu, B.; Duan, R.; Liu, Y. Callus induction and plant regeneration from rosemary leaves. Biosci. Methods 2012, 3, 21–26. [Google Scholar] [CrossRef]
- Boix, Y.F.; Arruda, R.C.O.; Defaveri, A.C.A.; Sato, A.; Lage, C.L.S.; Victorio, C.P. Callus in Rosmarinus officinalis L. (Lamiaceae): A morphoanatomical, histochemical and volatile analysis. Plant Biosyst. 2013, 147, 751–757. [Google Scholar] [CrossRef]
- Aman, N.; Afrasiab, H. Primary and secondary somatic embryogenesis from leaf explants of rosemary (Rosmarinus officinalis L.-Lamiaceae). Pakist. J. Bot. 2014, 46, 903–909. [Google Scholar]
- Al Masoody, M.M.M.; Stanica, F. Effect of growth regulators on in vitro callus formation of rosemary plant (Rosmarinus officinalis L.). Bull. UASVM Hortic. 2015, 72, 131–137. [Google Scholar] [CrossRef] [Green Version]
- El-Zefzafy, M.; Dawoud, G.; Shahhat, I. Physiological and phytochemical responses of rosemary (Rosmarinus officinalis L.) plant on in vitro callus formation. Eur. J. Med. Plants 2016, 17, 1–16. [Google Scholar] [CrossRef]
- Irum, S.; Khan, F.A.; Erum, S.; Nouroz, F.; Muhammad, A.; Kanwal, S. In vitro propagation of selected medicinal plants species. Asian J. Agric. Rural Dev. 2017, 7, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Darwesh, H.Y.; Alayafi, A.A. In virto propagation response of Rosmarinus officinalis L. to biotic and abiotic elicitors on phenolic content and photosynthetic pigments. J. Agric. Sci. 2018, 10, 301. [Google Scholar] [CrossRef] [Green Version]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; The Iowa State University Press: Ames, IA, USA, 1980; p. 507. [Google Scholar]
- Klockars, A.J.; Sax, G. Post hoc comparisons: The Scheffe test. In Multiple Comparisons; Klockars, A.J., Sax, G., Eds.; Sage Publications: Beverly Hills, CA, USA, 1986; Volume 61, pp. 58–61. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Zhao, F.; Chen, Y.P.; Salmaki, Y.; Drew, B.T.; Wilson, T.C.; Scheen, A.C.; Celep, F.; Bräuchler, C.; Bendiksby, M.; Wang, Q.; et al. An updated tribal classification of Lamiaceae based on plastomephylogenomics. BMC Biol. 2021, 19, 1–27. [Google Scholar] [CrossRef]
- Carrubba, A.; Abbate, L.; Sarno, M.; Sunseri, F.; Mauceri, A.; Lupini, A.; Mercati, F. Characterization of Sicilian rosemary (Rosmarinus officinalis L.) germplasm through a multidisciplinary approach. Planta 2020, 251, 37. [Google Scholar] [CrossRef]
- Herrera, J. Flower size variation in Rosmarinus officinalis: Individuals, populations and habitats. Ann. Bot. 2005, 95, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Cervelli, C.; Ruffoni, B.; Shachter, A.; Dudai, N. Volatile diversity in wild populations of rosemary (Rosmarinus officinalis L.) from the Tyrrhenian Sea vicinity cultivated under homogeneous environmental conditions. Ind. Crops Prod. 2016, 84, 381–390. [Google Scholar] [CrossRef]
- Del Baño, M.J.; Lorente, J.; Castillo, J.; Benavente-García, O.; Del Río, J.A.; Ortuño, A.; Quirin, K.W.; Gerard, D. Phenolic diterpenes, flavones, and rosmarinic acid distribution during the development of leaves, flowers, stems, and roots of Rosmarinus officinalis. Antioxidant activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef]
- Martín, J.P.; Bermejo, J.E.H. Genetic variation in the endemic and endangered Rosmarinus tomentosus Huber-Morath & amp; Maire (Labiatae) using RAPD markers. Heredity 2000, 85, 434–443. [Google Scholar] [CrossRef] [Green Version]
- Zaouali, Y.; Bouzaine, T.; Boussaid, M. Essential oils composition in two Rosmarinus officinalis L. varieties and incidence for antimicrobial and antioxidant activities. Food Chem. Toxicol. 2010, 48, 3144–3152. [Google Scholar] [CrossRef] [PubMed]
- Room, A. A Dictionary of True Etymologies, 1st ed.; Routledge (Taylor & Francis Group): New York, NY, USA, 1988; p. 198. [Google Scholar]
- Munné-Bosch, S.; Lopez-Carbonell, M.; Alegre, L.; Van Onckelen, H.A. Effect of drought and high solar radiation on 1-aminocyclopropane-1-carboxylic acid and abscisic acid concentrations in Rosmarinus officinalis plants. Physiol. Plant. 2002, 114, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Olmos, E.; Sánchez-Blanco, M.J.; Ferréndez, T.; Alarcón, J.J. Subcellular effects of drought stress in Rosmarinus officinalis. Plant Biol. 2007, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, J.; Morales, M.; Ferrandez, T.; Sanchez-Blanco, M. Effects of water and salt stresses on growth, water relations and gas exchange in Rosmarinus officinalis. J. Hortic. Sci. Biotechnol. 2006, 81, 845–853. [Google Scholar] [CrossRef]
- Nogués, I.; Muzzini, V.; Loreto, F.; Bustamante, M. Drought and soil amendment effects on monoterpene emission in rosemary plants. Sci. Total Environ. 2015, 538, 768–778. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Romano, D. Response of Mediterranean ornamental plants to drought stress. Horticulturae 2019, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Cassells, A.C. Problems in tissue culture: Culture contamination. In Micropropagation; Springer: Dordrecht, The Netherlands, 1991; pp. 31–44. [Google Scholar] [CrossRef]
- Tsoulpha, P.; Alexandri, S.; Tsaktsira, M. Critical factors affecting an efficient micropropagation protocol for Pyrus spinosa Forskk. J. Appl. Hortic. 2018, 20, 190–195. [Google Scholar] [CrossRef]
- Sakr, S.S.; Amin, A.Y.; El-Mewafy, E.A.; Eid, N.M. In vitro comparative study on Rosmarinus officinalis L. cultivars. Middle East J. Agric. Res. 2018, 7, 703–715. [Google Scholar]
- Thomas, T.D. The role of activated charcoal in plant tissue culture. Biotechnol. Adv. 2008, 26, 618–631. [Google Scholar] [CrossRef]
- Tien Vinh, D.; Hoa, M.T.; Khai, P.C.; Van Minh, T. Micropropagation of lavender (Lavandula angustifolia). J. Innov. Pharm. Biol. Sci. 2017, 4, 7–11. [Google Scholar]
- Otroshy, M.; Moradi, K. Micropropagation of medicinal plant Dracocephalum kotschyi Boiss. via nodal cutting technique. J. Med. Plant Res. 2011, 5, 5967–5972. [Google Scholar] [CrossRef]
- Van Tan, P. Micropropagation of Curcuma sp., a threatened medicinal plant. Adv. Biosci. Biotechnol. 2016, 7, 418. [Google Scholar] [CrossRef] [Green Version]
- Kereša, S.; Andrijanić, Z.; Kremer, D.; HabušJerčić, I.; Barić, M.; BateljaLodeta, K.; Bolarić, S.; Bošnjak-Mihovilović, A. Efficient micropropagation and rooting of Micromeria croatica (Pers.) Schott (Lamiaceae). Poljoprivreda 2018, 24, 27–33. [Google Scholar] [CrossRef]
- Pan, M.J.; Staden, J.V. The use of charcoal in in vitro culture—A review. Plant Growth Regul. 1998, 26, 155–163. [Google Scholar] [CrossRef]
- Buckseth, T.; Singh, R.K.; Sharma, A.K.; Sharma, S.; Moudgil, V.; Saraswati, A. Optimization of activated charcoal on in vitro growth and development of potato (Solanum tuberosum L.). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 3543–3548. [Google Scholar] [CrossRef]
- Mascarello, C.; Sacco, E.; Pamato, M.; Di Silvestro, D.; Bassolino, L.; Cervelli, C.; Ruffoni, B. Rosmarinus officinalis L.: Micropropagation and callus induction for cell biomass development. Acta Hortic. 2017, 1155, 631–636. [Google Scholar] [CrossRef]
- Husain, Z.M.A.; Jawad, L.K. Effect of some growth regulators on the multiplication and stimulating the production of the volatile oil of Rosemary officinalis in vitro. Plant Arch. 2019, 19, 1773–1782. [Google Scholar]
- Singh, N.K.; Sehgal, C.B. Micropropagation of ‘Holy Basil’ (Ocimum sanctum Linn.) from young inflorescences of mature plants. Plant Growth Regul. 1999, 29, 161–166. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Bilichowski, I.; Mikiciuk-Olasik, E.; Wysokińska, H. In vitro cultures of Salvia officinalis L. as a source of antioxidant compounds. Acta Soc. Bot. Pol. 2005, 74, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Dode, L.B.; Bobrowski, V.L.; Braga, E.J.B.; Seixas, F.K.; Schuch, M.W. In vitro propagation of Ocimum basilicum L. (Lamiaceae). Acta Sci. Biol. Sci. 2003, 25, 435–437. [Google Scholar] [CrossRef]
- Ghaderi, S.; Ebrahimi, S.N.; Ahadi, H.; Moghadam, S.E.; Mirjalili, M.H. In vitro propagation and phytochemical assessment of Perovskia abrotanoides Karel. (Lamiaceae)—A medicinally important source of phenolic compounds. Biocatal. Agric. Biotechnol. 2019, 19, 101–113. [Google Scholar]
- Dumas, E.; Monteuuis, O. In vitro rooting of micropropagated shoots from juvenile and mature Pinus pinaster explants: Influence of activated charcoal. Plant Cell Tissue Organ Cult. 1995, 40, 231–235. [Google Scholar] [CrossRef]
- Economou, A.S. From macrocutting rooting to microplant establishment: Key points to consider for maximum success in woody plants. Acta Hortic. 2013, 988, 43–46. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
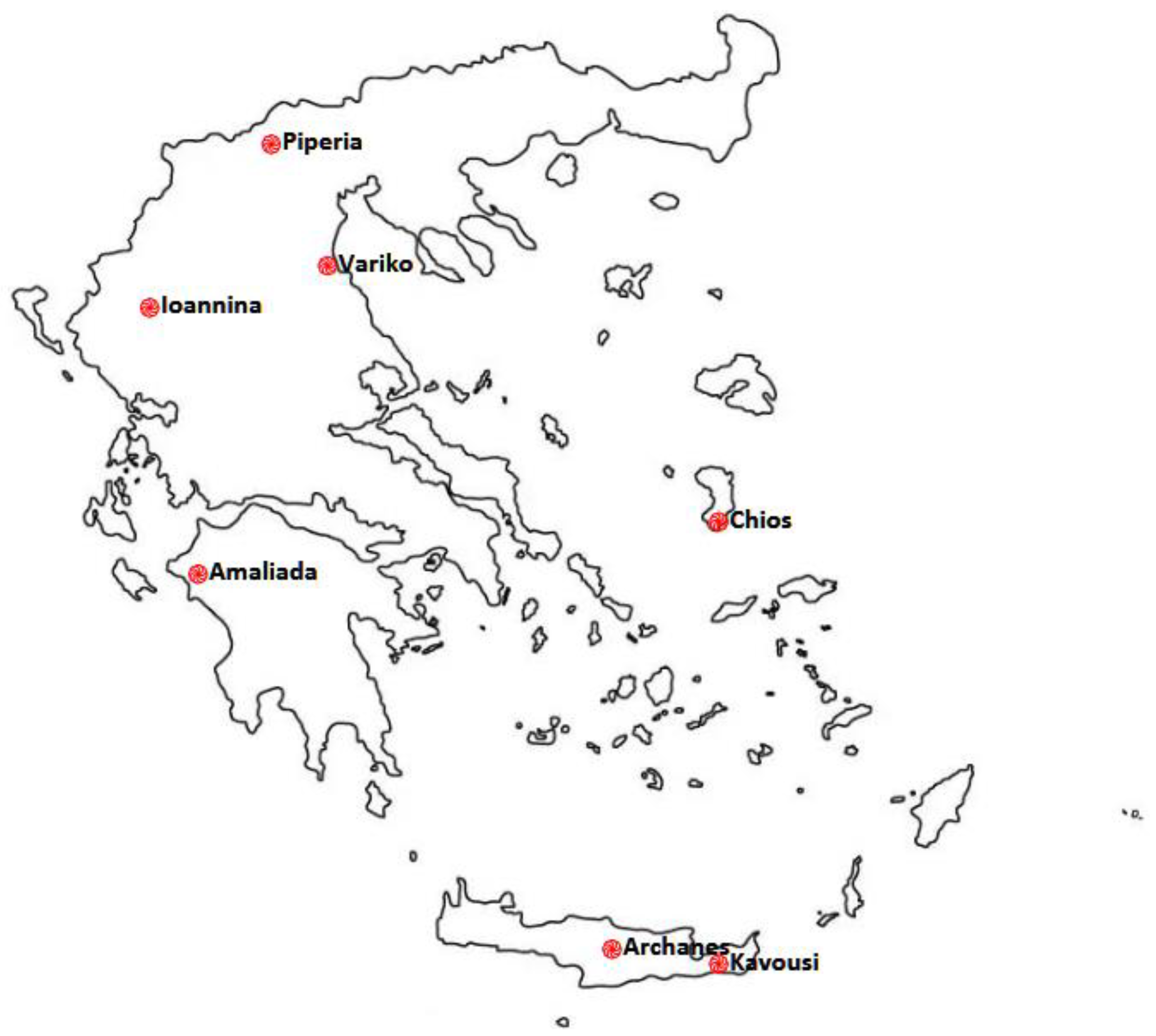
| Origin—Population | Plants per Population | Latitude N | Longitude E | Εlevation (m) |
|---|---|---|---|---|
| Amaliada (AM) | 9 | 37.769800° | 21.303847° | 11 |
| Ioannina (I) | 3 | 39.645606° | 20.849649° | 481 |
| Chios (CH) | 3 | 38.243976° | 26.035575° | 57 |
| Variko (V) | 9 | 40.192397° | 22.550020° | 12 |
| Piperia (P) | 14 | 40.964379° | 22.017295° | 143 |
| Archanes (AR) | 6 | 35.233768° | 25.152781° | 382 |
| Kavousi (KA) | 3 | 35.118470° | 25.856935° | 173 |
| Morphological Traits | Details | Measurements per Plant of each Population |
|---|---|---|
| Plant canopy height | 1 |
| Plant canopy diameter | 1 |
| Terminal shoots of new vegetation per plant | 1 |
| Terminal shoots of new vegetation | 10 |
| Nodes per terminal branch | 10 |
| Leaves per terminal branch | 10 |
| Measured from the base to the tip of adult leaf | 30 |
| Measured at the widest part of adult leaf | 30 |
| Code Number | PVP * | Ascorbic and Citric Acid ** | NaOCl (%) | Disinfestation Duration (min) | Contamination-Free Explants (%) |
|---|---|---|---|---|---|
| 1 | − | − | 0.5 | 7 | 88.9 ab *** |
| 2 | − | − | 0.625 | 8 | 88.9 ab |
| 3 | − | − | 0.5 | 5 | 88.9 ab |
| 4 | − | − | 0.625 | 5 | 100.0 a |
| 5 | − | − | 0.3 | 7 | 100.0 a |
| 6 | − | − | 0.3 | 5 | 0.0 f |
| 7 | − | − | 0.3 | 3 | 0.0 f |
| 8 | − | − | 0.4 | 4 | 0.0 f |
| 9 | − | − | 0.4 | 3 | 22.2 e |
| 10 | − | − | 0.4 | 4 | 0.0 f |
| 11 | − | − | 0.3 | 5 | 0.0 f |
| 12 | − | − | 0.3 | 6 | 44.4 cde |
| 13 | − | − | 0.4 | 3 | 0.0 f |
| 14 | − | − | 0.3 | 4 | 22.2 e |
| 15 | + | − | 0.5 | 5 | 100.0 a |
| 16 | + | − | 0.15 | 30 | 66.7 bc |
| 17 | + | − | 0.15 | 20 | 55.6 cd |
| 18 | + | − | 0.15 | 15 | 33.3 de |
| 19 | + | − | 0.15 | 10 | 33.3 de |
| 20 | − | + | 0.6 | 7 | 100.0 a |
| Nutrient Medium | IAA (mg·L−1) | BAP (mg·L−1) |
|---|---|---|
| MS1 | 0 | 0 |
| MS2 | 0.1 | 0 |
| MS3 | 0.1 | 0.25 |
| MS4 | 0.1 | 0.5 |
| MS5 | 0.1 | 1 |
| Population | K-IBA g·L−1 | Spring | Summer | Autumn | Winter | |
|---|---|---|---|---|---|---|
| Number of roots | ‘Piperia’ | 0 | 3.11 ± 0.51 * b ** | 3.35 ± 0.42 b | 3.34 ± 0.59 b | 2.77 ± 0.18 b |
| 0.5 | 5.73 ± 0.29 a | 5.72 ± 0.38 a | 5.94 ± 0.51 a | 4.33 ± 0.19 a | ||
| 1 | 6.10 ± 1.10 a | 6.28 ± 0.52 a | 6.41 ± 0.31 a | 4.75 ± 0.42 a | ||
| ‘Amaliada’ | 0 | 2.79 ± 0.22 b | 3.11 ± 0.25 b | 5.18 ± 0.70 a | 2.68 ± 0.37 b | |
| 0.5 | 5.86 ± 0.47 a | 5.95 ± 0.36 a | 5.70 ± 0.55 a | 4.54 ± 0.50 a | ||
| 1 | 6.07 ± 0.85 a | 6.16 ± 0.68 a | 6.69 ± 0.46 a | 4.71 ± 0.46 a | ||
| Length of roots (cm) | ‘Piperia’ | 0 | 7.70 ± 0.61 a | 8.19 ± 0.91 a | 8.58 ± 0.68 a | 5.91 ± 0.49 a |
| 0.5 | 7.60 ± 0.57 a | 7.58 ± 0.58 ab | 8.93 ± 0.75 a | 5.57 ± 0.52 a | ||
| 1 | 7.17 ± 1.02 a | 7.31 ± 0.64 b | 7.77 ± 0.34 a | 6.36 ± 0.47 a | ||
| ‘Amaliada’ | 0 | 7.56 ± 0.93 a | 7.95 ± 0.61 a | 7.34 ± 0.55 a | 6.18 ± 0.38 a | |
| 0.5 | 7.44 ± 0.74 a | 7.63 ± 0.69 ab | 7.88 ± 0.28 a | 5.42 ± 0.40 a | ||
| 1 | 8.21 ± 0.39 a | 6.27 ± 0.45 b | 8.51 ± 0.47 a | 5.34 ± 0.51 a |
| Number of Roots | Length of Roots (cm) | |||
|---|---|---|---|---|
| Perlite: Peat | Fog | Mist | Fog | Mist |
| 1:0 | 5.74 a * | 5.86 a | 3.71 a | 3.57 a |
| 2:1 | 5.59 a | 6.20 a | 4.14 a | 3.86 a |
| 1:1 | 5.47 a | 6.01 a | 3.92 a | 3.78 a |
| Media | Population | Shoot Formation Frequency (%) | Number of Shoots | Length of Shoots (cm) | Root Formation Frequency (%) | Length of Roots (cm) |
|---|---|---|---|---|---|---|
| MS1 | ‘Amaliada’ | 91.6 ± 14.4 * a ** | 2.00 ± 0.21 a | 0.68 ± 0.04 e | 16.6 ± 14.4 b | 6.25 ± 0.75 ab |
| ‘Piperia’ | 33.3 ± 14.4 d | 0.33 ± 0.14 e | 1.08 ± 0.19 ab | 25.0 ± 25.0 ab | 2.50 ± 0.29 f | |
| MS2 | ‘Amaliada’ | 66.6 ± 14.4 bc | 1.50 ± 0.42 ab | 0.91 ± 0.08 bc | 41.6 ± 14.4 ab | 2.80 ± 0.21 ef |
| ‘Piperia’ | 8.3 ± 14.4 e | 0.08 ± 0.08 f | 0.35 ± 0.15 f | 0 c | 0 g | |
| MS3 | ‘Amaliada’ | 75.0 ± 25.0 ab | 1.25 ± 0.28 bc | 0.79 ± 0.07 de | 33.3 ± 14.4 ab | 3.85 ± 0.72 d |
| ‘Piperia’ | 66.6 ± 14.4 bc | 0.67 ± 0.14 cd | 0.93 ± 0.16 bc | 8.3 ± 14.4 bc | 3.75 ± 0.75 de | |
| MS4 | ‘Amaliada’ | 58.3 ± 14.4 bc | 1.08 ± 0.31 bc | 0.94 ± 0.05 bc | 50.0 ± 25.0 a | 7.08 ± 0.71 a |
| ‘Piperia’ | 66.6 ± 14.4 bc | 0.67 ± 0.14 cd | 1.26 ± 0.18 a | 41.6 ± 14.4 ab | 5.30 ± 0.54 bc | |
| MS5 | ‘Amaliada’ | 50.0 ± 25.0 cd | 0.92 ± 0.36 bcd | 0.85 ± 0.08 cd | 25.0 ± 25.0 ab | 4.00 ± 0.58 d |
| ‘Piperia’ | 58.3 ± 14.4 bc | 0.58 ± 0.15 de | 0.83 ± 0.05 cd | 16.6 ± 14.4 b | 4.25 ± 0.75 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostas, S.; Kaplani, A.; Koulaouzidou, E.; Kotoula, A.-A.; Gklavakis, E.; Tsoulpha, P.; Hatzilazarou, S.; Nianiou-Obeidat, I.; Kanellis, A.K.; Economou, A. Sustainable Exploitation of Greek Rosmarinus officinalis L. Populations for Ornamental Use through Propagation by Shoot Cuttings and In Vitro Cultures. Sustainability 2022, 14, 4059. https://doi.org/10.3390/su14074059
Kostas S, Kaplani A, Koulaouzidou E, Kotoula A-A, Gklavakis E, Tsoulpha P, Hatzilazarou S, Nianiou-Obeidat I, Kanellis AK, Economou A. Sustainable Exploitation of Greek Rosmarinus officinalis L. Populations for Ornamental Use through Propagation by Shoot Cuttings and In Vitro Cultures. Sustainability. 2022; 14(7):4059. https://doi.org/10.3390/su14074059
Chicago/Turabian StyleKostas, Stefanos, Aikaterini Kaplani, Efthymia Koulaouzidou, Aikaterini-Angeliki Kotoula, Evangelos Gklavakis, Parthena Tsoulpha, Stefanos Hatzilazarou, Irini Nianiou-Obeidat, Angelos K. Kanellis, and Athanasios Economou. 2022. "Sustainable Exploitation of Greek Rosmarinus officinalis L. Populations for Ornamental Use through Propagation by Shoot Cuttings and In Vitro Cultures" Sustainability 14, no. 7: 4059. https://doi.org/10.3390/su14074059