
Facilitating Conservation and Bridging Gaps for the Sustainable Exploitation of the Tunisian Local Endemic Plant Marrubium aschersonii (Lamiaceae)

 , , , , , , , , and
, , , , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
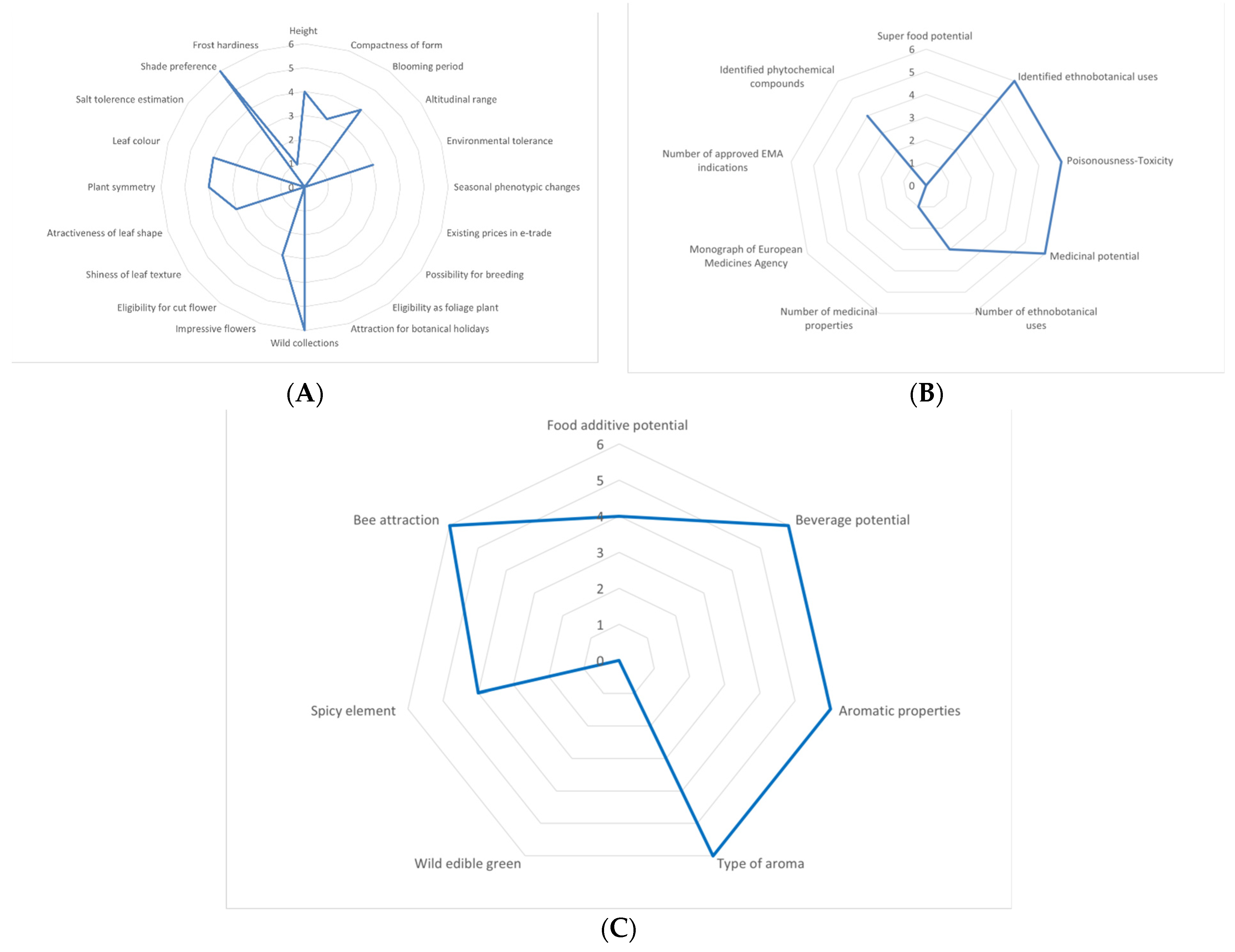
2.1. Multifaceted Evaluation in Different Economic Sectors
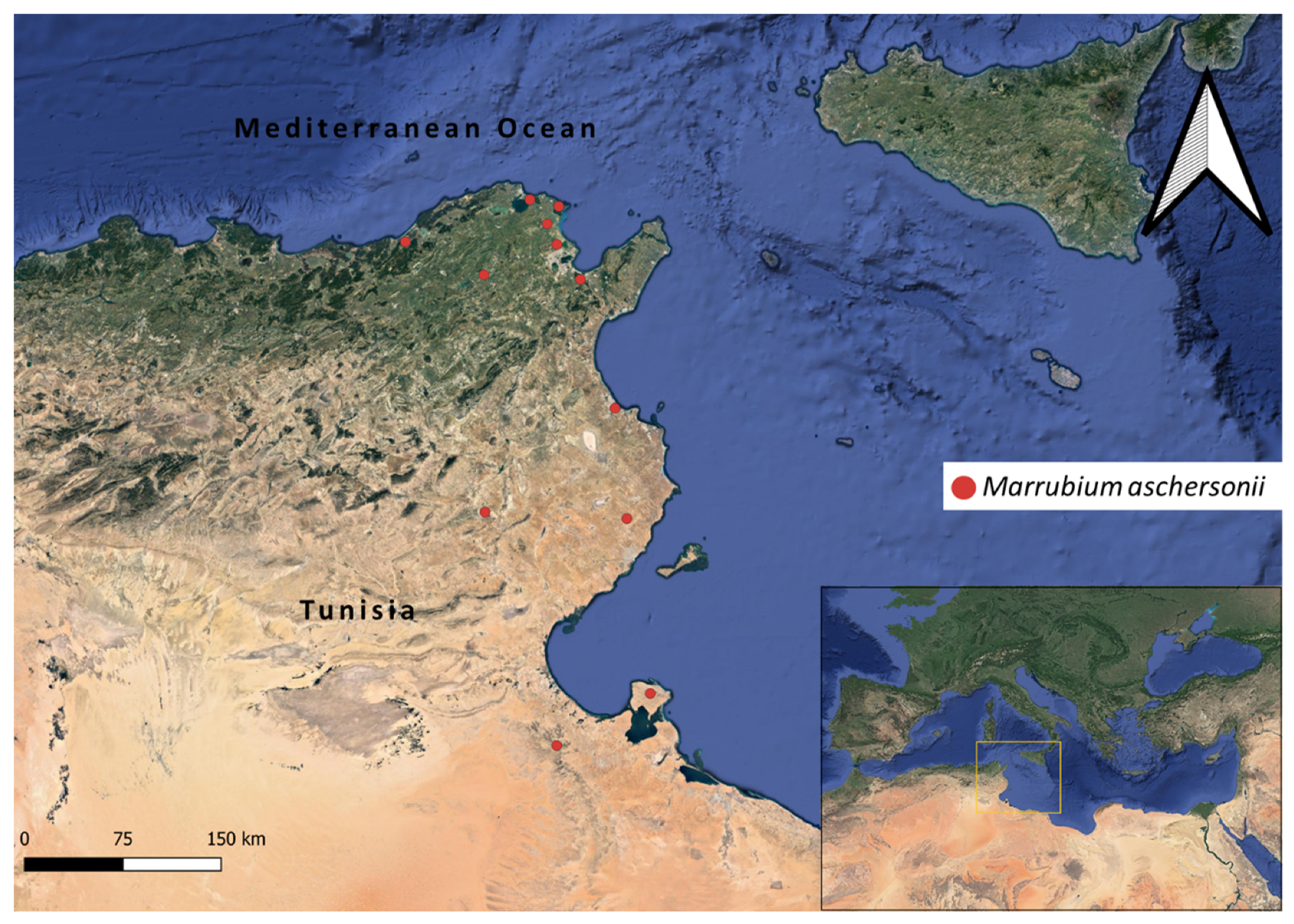
2.2. Distribution Mapping and GIS Ecological Profiling
- (a)
- WorldClim version 2.1, containing minimum, maximum, and average temperatures (°C) as well as precipitation values (mm) and data for 19 bioclimatic variables for every month derived from 1970–2000, with a raster resolution of 1 km2, and
- (b)
- Marrubium aschersonii distribution points raster file (Figure 1).
2.3. Seed Collection and Storage
2.4. Germination Test
2.5. Molecular Markers and PCR Procedures
2.6. Pilot Cultivation of Seedlings, Transplanting, and Fertilization Treatments
2.7. Morphological and Physiological Measurements of Seedlings
2.8. Plant Tissue Analyses of Seedlings
2.9. Statistical Analysis
3. Results
3.1. Overview of the Potential of Marrubium aschersonii in Economic Sectors
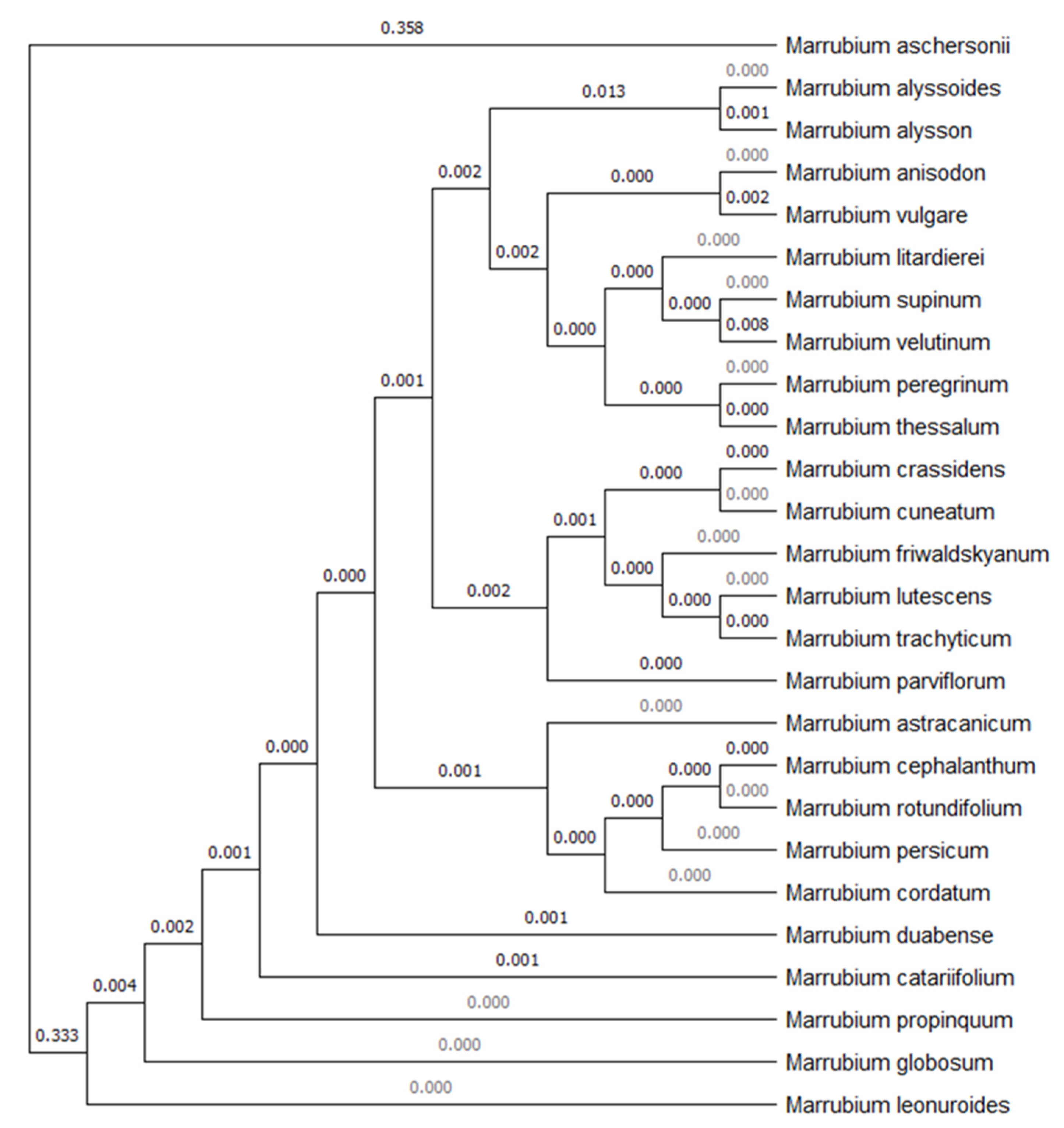
3.2. Molecular Characterization of M. aschersonii and Annotation in GenBank
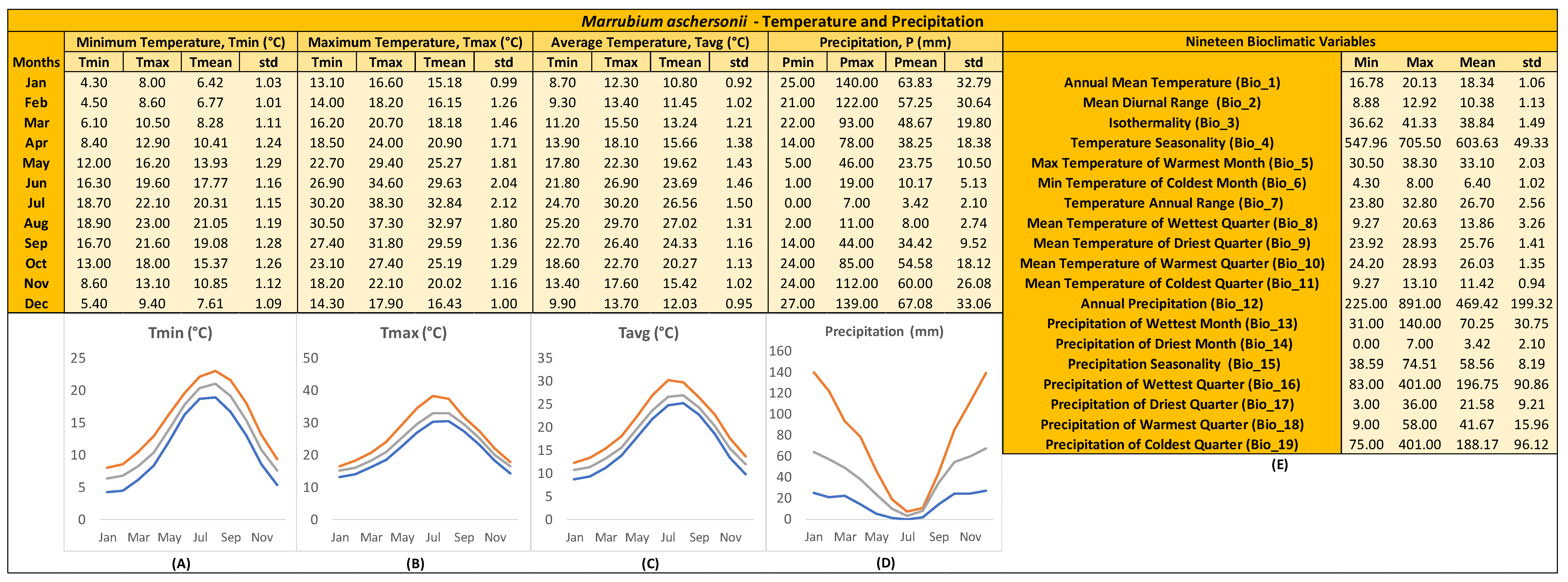
3.3. Ecological Profiling of Marrubium aschersonii
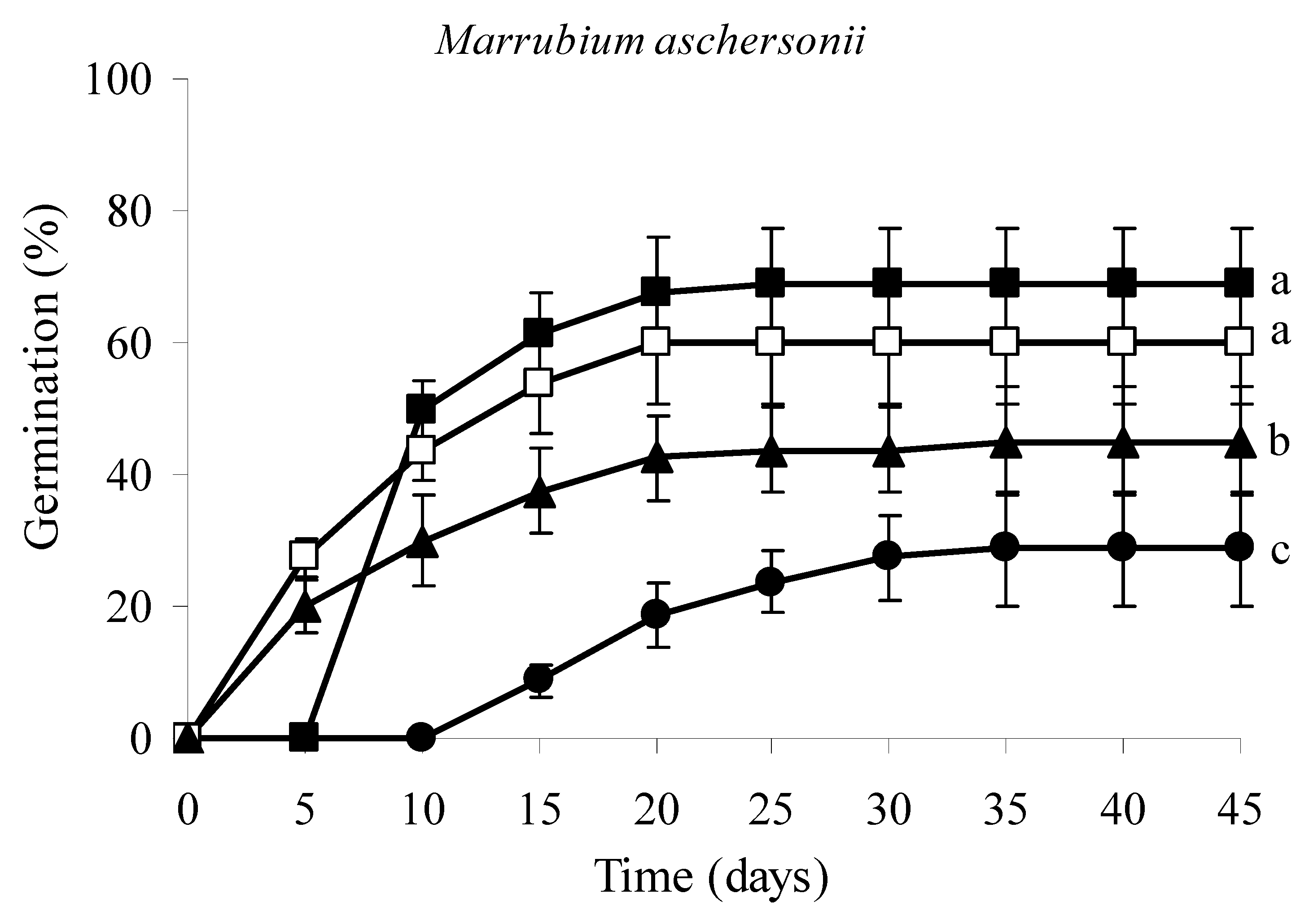
3.4. Seed Germination Tests
3.5. Seedling Growth in Pilot Cultivation
3.6. Macro- and Micronutrient Content
4. Discussion
4.1. Molecular Authentication (DNA Barcoding) of M. aschersonii
4.2. GIS-Facilitated Seed Germination
4.3. M. aschersonii Seedling Growth and Response in Fertilization
4.4. Re-Evaluation of Feasibility and Readiness Timescale for the Sustainable Exploitation of M. aschersonii
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyre-Silva, C.; Cechinel-Filho, V. A review of the chemical and pharmacological aspects of the genus Marrubium. Curr. Pharm. Des. 2010, 16, 3503–3518. [Google Scholar] [CrossRef] [PubMed]
- Siadati, S.; Salmaki, Y.; Saeidi, M.S.; Weigend, M. Untangling the generic boundaries in tribe Marrubieae (Lamiaceae: Lamioideae) using nuclear and plastid DNA sequences. Taxon 2018, 67, 770–783. [Google Scholar] [CrossRef]
- Wink, M.; Kaufmann, M. Phylogenetic relationships between some members of the Subfamily Lamioideae (Family Labiatae) inferred from nucleotide sequences of the rbcL gene. Bot. Acta. 1995, 109, 139–148. [Google Scholar] [CrossRef]
- Elsherbeny, A.E. DNA barcoding of some medicinal plants, family Labiatae. Egypt. J. Exp. Biol. 2016, 12, 175–180. [Google Scholar] [CrossRef]
- Scheen, A.C.; Bendiksby, M.; Ryding, O.; Mathiesen, C.; Albert, V.A.; Lindqvist, C. Molecular phylogenetics, character evolution, and suprageneric classification of Lamioideae (Lamiaceae). Ann. Missouri Bot. Gard. 2010, 97, 191–217. [Google Scholar] [CrossRef]
- Roy, T.; Lindqvist, C. New insights into evolutionary relationships within the subfamily Lamioideae (Lamiaceae) based on pentatricopeptide repeat (PPR) nuclear DNA sequences. Amer. J. Bot. 2015, 102, 1721–1735. [Google Scholar] [CrossRef] [Green Version]
- Marzouk, R.I.; El-Darier, S.M.; Nour, I.H.; Kamal, S.A. Numerical taxonomic study of Marrubium L. (Lamiaceae) in Egypt. Catrina 2015, 13, 25–35. Available online: https://cat.journals.ekb.eg/article_18376.html (accessed on 30 December 2021).
- Salehi, N.; Kharazian, N.; Shiran, B. Genetic diversity of Marrubium species from Zagros region (Iran), using inter simple sequence repeat molecular marker. J. Sci. Islamic Repub. Iran. 2018, 29, 7–19. [Google Scholar] [CrossRef]
- Sanna, G.; Farci, P.; Busonera, B.; Murgia, G.; La Colla, P.; Giliberti, G. Antiviral properties from plants of the Mediterranean flora. Nat. Prod. Res. 2015, 29, 2065–2070. [Google Scholar] [CrossRef]
- Hashemi, M.; Kharazian, N. Identification of flavonoids from Marrubium and Ballota species (Lamiaceae) and determination of chemotaxonomic markers using High Performance Liquid Chromatography Mass Spectrometer. J. Sci. Islam Repub. Iran 2021, 32, 305–320. [Google Scholar] [CrossRef]
- Zaabat, N.; Hay, A.E.; Michalet, S.; Darbour, N.; Bayet, C.; Skandrani, I.; Chekir-Ghedira, L.; Akkal, S.; Dijoux-Franca, M.G. Antioxidant and antigenotoxic properties of compounds isolated from Marrubium deserti de Noé. Food Chem. Toxicol. 2011, 49, 3328–3335. [Google Scholar] [CrossRef] [PubMed]
- Hamdaoui, B.; Wannes, W.A.; Marrakchi, M.; Brahim, N.B.; Marzouk, B. Essential oil composition of two Tunisian Horehound species: Marrubium vulgare L. and Marrubium aschersonii Magnus. J. Essent. Oil Bear. Plants 2013, 16, 608–612. [Google Scholar] [CrossRef]
- Hammami, S.; Li, Z.; Huang, M.; El Mokni, R.; Dhaouadi, H.; Yin, S. New bioactive labdane diterpenoids from Marrubium aschersonii. Nat. Prod. Res. 2016, 30, 2142–2148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Zou, Y.H.; Zhao, J.J.; Chen, Y.; Bao, J.M.; Tang, G.H. Three new diterpenoids from Marrubium aschersonii. Phytochem. Lett. 2016, 16, 241–244. [Google Scholar] [CrossRef]
- De Jesus, R.A.P.; Cechinel-Filho, V.; Oliveira, A.E.; Schlemper, V. Analysis of the antinociceptive properties of marrubiin isolated from Marrubium vulgare. Phytomedicine 2000, 7, 111–115. [Google Scholar] [CrossRef]
- Sahpaz, S.; Garbacki, N.; Tits, M.; Bailleul, F. Isolation and pharmacological activity of phenylpropanoid esters from Marrubium vulgare. J. Ethnopharmacol. 2002, 79, 389–392. [Google Scholar] [CrossRef]
- Stulzer, H.K.; Tagliari, M.P.; Zampirolo, J.A.; Cechinel-Filho, V.; Schlemper, V. Antioedematogenic effect of marrubiin obtained from Marrubium vulgare. J. Ethnopharmacol. 2006, 108, 379–384. [Google Scholar] [CrossRef]
- Wiesner, J. Assessment Report on Marrubium vulgare L., Herba. EMA/HMPC/604273/2012, Committee on Herbal Medicinal Products (HMPC), European Medicines Agency. 2013. Available online: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-marrubium-vulgare-l-herba-first-version_en.pdf (accessed on 29 December 2021).
- Pottier-Alapetite, G. Angiosperrnes Dicotylédones: Apétales-dialypétales-gamopétales. Première et deuxième partie.-Ouvrage publié par le Ministère de l’Enseignement Supérieur et de la Recherche Scientifique et le Ministère de l’Agriculture-Tunisie. Flore Tunisie 1981, 1190. [Google Scholar]
- Grigoriadou, K.; Krigas, N.; Lazari, D.; Maloupa, E. Chapter 4-Sustainable use of Mediterranean medicinal-aromatic plants. In Feed Additives; Florou-Paneri, P., Christaki, E., Giannenas, I., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2020; pp. 57–74. ISBN 9780128147009. [Google Scholar] [CrossRef]
- Dadach, M. Recherche des Conditions Optimales de la Germination des Graines de Quelques Labiées du Mont de Tessala (Ouest Algérien) et Perspectives de Conservation. Ph.D. Thesis, Université Djillali Liabes de Sidi Bel Abbes, Sidi Bel Abbes, Algérie, October 2016. [Google Scholar]
- Belmehdi, O.; El Harsal, A.; Benmoussi, M.; Laghmouchi, Y.; Skali Senhaji, N.; Abrini, J. Effect of light, temperature, salt stress and pH on seed germination of medicinal plant Origanum elongatum (Bonnet) Emb. & Maire. Biocatal. Agric. Biotechnol. 2018, 16, 126–131. [Google Scholar] [CrossRef]
- Flores-Enríquez, V.; Castillo, G.; Collazo-Ortega, M. Experimental seed germination for ex situ conservation of Mexican Podostemaceae. Bot. Sci. 2019, 97, 413–422. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Sarropoulou, V.; Krigas, N.; Maloupa, E.; Tsoktouridis, G. GIS-facilitated effective propagation protocols of the Endangered local endemic of Crete Carlina diae (Rech. f.) Meusel and A. Kástner (Asteraceae): Serving ex-situ conservation needs and its future sustainable exploitation as an ornamental. Plants 2020, 9, 1465. [Google Scholar] [CrossRef] [PubMed]
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M. Exploring the potential of neglected local endemic plants of three Mediterranean regions in the ornamental sector: Value chain feasibility and readiness timescale for their sustainable exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Bourgou, S.; Ben, I.H.J.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; et al. Medicinal-cosmetic potential of the local endemic plants of Crete (Greece), Northern Morocco and Tunisia: Priorities for conservation and sustainable exploitation of neglected and underutilized phytogenetic resources. Biology 2021, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Fanourakis, D.; Paschalidis, K.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Liapaki, E.; Juini, M.; Ipsilantis, I.; Maloupa, E.; et al. Pilot cultivation of the local endemic Cretan marjoram Origanum microphyllum (Benth.) Vogel (Lamiaceae): Effect of fertilizers on growth and herbal quality features. Agronomy 2022, 12, 94. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; El Haissoufi, M.; Pipinis, E.; Kostas, S.; Libiad, M.; Khabbach, A.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; et al. GIS-facilitated seed germination and multifaceted evaluation of the endangered Abies marocana Trab. (Pinaceae): Enabling conservation and sustainable exploitation. Plants 2021, 10, 2606. [Google Scholar] [CrossRef] [PubMed]
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Anestis, I.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Greveniotis, V.; Tsiripidis, I.; et al. Agro-alimentary potential of the neglected and underutilized local endemic plants of Crete (Greece), Rif-Mediterranean coast of Morocco and Tunisia: Perspectives and challenges. Plants 2021, 10, 1770. [Google Scholar] [CrossRef]
- Maloupa, E.; Karapatzak, E.; Ganopoulos, I.; Karydas, A.; Papanastasi, K.; Kyrkas, D.; Yfanti, P.; Nikisianis, N.; Zahariadis, A.; Kosma, I.S.; et al. Molecular Authentication, Phytochemical evaluation and asexual propagation of wild-growing Rosa canina L. (Rosaceae) genotypes of Northern Greece for sustainable exploitation. Plants 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Paschalidis, K.; Fanourakis, D.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Kalogiannakis, K.; Debouba, F.J.; Ipsilantis, I.; Tsoktouridis, G.; et al. Pilot cultivation of the vulnerable Cretan endemic Verbascum Arcturus L. (Scrophulariaceae): Effect of fertilization on growth and quality features. Sustainability 2021, 13, 14030. [Google Scholar] [CrossRef]
- Macdonald, B. Practical Woody Plant Propagation for Nursery Growers; Timber Press Inc.: Portland, OR, USA, 2006; p. 669. [Google Scholar]
- Kostopoulou, P.; Radoglou, K.; Dini-Papanastasi, O.; Spyroglou, G. Enhancing planting stock quality of Italian cypress (Cupressus sempervirens L.) by pre-cultivation in mini-plugs. Ecol. Eng. 2010, 36, 912–919. [Google Scholar] [CrossRef]
- O’Reill, C.; Arnott, J.T.; Owens, J.N. Effects of photoperiod and moisture availability on shoot growth, seedling morphology, and cuticle and epicuticular wax features of container-grown western hemlock seedlings. Can. J. For. Res. 1989, 19, 122–131. [Google Scholar] [CrossRef]
- Radoglou, K.M. Effect of environmental stress on planting stock and field performance. In Proceedings of the National Forestry Conference: Planting Stock of Woody Species, Thessaloniki, Greece, 28–29 January 1999; pp. 15–26. [Google Scholar]
- Buendia Velazquez, M.V.; Lopez Lopez, M.A.; Cetina Alcala, V.M.; Diakite, L. Substrates and nutrient addition rates affect morphology and physiology of Pinus leiophylla seedlings in the nursery stage. iForest 2016, 10, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.; Planelles, R.; Artero, F.; Valverde, R.; Jacobs, D.F.; Segura, M. Field performance of Pinus halepensis planted in Mediterranean arid conditions: Relative influence of seedling morphology and mineral nutrition. New For. 2009, 37, 313–331. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Puertolas, J.; Penuelas, J.L.; Planelles, R. Effect of nitrozen fertilization in the nursery on the drought and frost resistance of Mediterranean forest species. For. Syst. 2005, 14, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Krigas, N.; Mouflis, G.; Grigoriadou, K. Conservation of important plants from the Ionian Islands at the Balkan Botanic Garden of Kroussia, N Greece: Using GIS to link the in situ collection data with plant propagation and ex situ cultivation. Biodivers. Conserv. 2010, 19, 3583–3603. [Google Scholar] [CrossRef]
- Krigas, N.; Papadimitriou, K.; Mazaris, A.D. GIS and Ex-Situ Plant Conservation; Alam, B.M., Ed.; Application of Geographic Information Systems, InTechopen.com: Rijeka, Croatia, 2012; Available online: https://www.intechopen.com/chapters/40525 (accessed on 29 December 2021).
- Cuénod, A. Flore Analytique et Synoptique de la Tunisie (Cryptogames Vasculaires, Gymnospermes et Monocotylédones); Office de l’Expérimentation et de la Vulgarisation Agricoles: Tunisie, Tunis, 1954. [Google Scholar]
- Pottier-Alapetite, G. Flore de la Tunisie (Angiospermes-Dicotylédones); Publications Scientifiques Tunisiennes, Ministère de l’Enseignement supérieur et de la Recherche Scientifique, Ministère de l’Agriculture: Tunis, Tunisie, 1981. [Google Scholar]
- Tsoktouridis, G.; Tsiamis, G.; Koutinas, N.; Mantell, S. Molecular detection of bacteria in plant tissues using universal 16S ribosomal DNA degenerated primers. Biotechnol. Biotechol. Equip. 2014, 28, 583–591. [Google Scholar] [CrossRef]
- Tsoktouridis, G.; Krigas, Ν.; Sarropoulou, V.; Kampouropoulou, S.; Papanastasi, K.; Grigoriadou, Κ.; Menexes, G.; Maloupa, E. Micropropagation and molecular characterization of Thymus sibthorpii Benth. (Lamiaceae), an aromatic-medicinal thyme with ornamental value and conservation concern. In Vitro Cell. Dev. Biol. 2019, 55, 647–658. [Google Scholar] [CrossRef]
- Van de Peer, Y. Phylogenetic inference based on distance methods. In The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing; Lemey, P., Salemi, M., Vandamme, A.M., Eds.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytologist 2002, 153, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Cessna, S.; Demmig-Adams, B.; Adams, W.W., III. Exploring photosynthesis and plant stress using inexpensive chlorophyll fluorometers. J. Nat. Resourc. Life Sci. Educ. 2010, 39, 22–30. [Google Scholar] [CrossRef]
- Allen, S.E.; Grimshaw, H.M.; Rowland, A.P. Chemical analysis. In Methods in Plant Ecology; Moore, P.D., Chapman, S.B., Eds.; Blackwell Scientific Publication: Oxford, UK, 1986; pp. 285–344. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis Part 2 Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Bremmer, J.M.; Mulvaney, C.S. Nitrogen-total. In Methods of Soil Analysis. Part 2: Chemical and Microbial Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Klockars, A.; Sax, G. Multiple Comparisons; Sage Publications: Newbury Park, CA, USA, 1986; p. 87. [Google Scholar]
- Snedecor, G.W.; Cochran, W.C. Statistical Methods, 7th ed.; The Iowa State University Press: Ames, IA, USA, 1980; p. 507. [Google Scholar]
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Sharrock, S.; Krigas, N. Ex-situ conservation of single-country endemic plants of Tunisia and northern Morocco (Mediterranean coast and Rif region) in seed banks and botanic gardens worldwide. Kew Bull. 2020, 75, 46. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; Fazekas, A.J.; et al. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Chaudhary, V.; Sirohi, U.; Sharma, V.R.; Naresh, R.K. Application of molecular markers and their utility in genetic studies of floricultural crops: A review. Inter. J. Agric. Environ. Biotechnol. 2019, 12, 229–247. [Google Scholar] [CrossRef]
- Theodoridis, S.; Stefanaki, A.; Tezcan, M.; Aki, C.; Kokkini, S.; Vlachonasios, K.E. DNA barcoding in native plants of the Labiatae (Lamiaceae) family from Chios Island (Greece) and the adjacent Cesme-Karaburun Peninsula (Turkey). Mol. Ecol. Resour. 2012, 12, 620–633. [Google Scholar] [CrossRef] [PubMed]
- Chase, W.; Cowan, M.R.; Hollingsworth, M.; van den Berg, P.C.; Madriñán, S.; Petersen, G.; Seberg, O.; Jorgsensen, T.; Cameron, K.; Carine, M.; et al. A proposal for a standardized protocol to barcode all land plants. Taxon 2007, 56, 295–299. [Google Scholar] [CrossRef]
- Löhne, C.; Borsch, T. Phylogenetic utility and molecular evolution of the petD group II intron in basal angio-sperms. Mol. Biol. Evol. 2005, 22, 317–332. [Google Scholar] [CrossRef]
- Barber, J.C.; Francisco-Ortega, J.; Santos-Guerra, A.; Turner, K.G.; Jansen, R.K. Origin of Macaronesian sideritis L. (Lamioideae: Lamiaceae) inferred from nuclear and chloroplast sequence datasets. Mol. Phylogenet. Evol. 2002, 23, 293–306. [Google Scholar] [CrossRef]
- Salmaki, Y.; Heubl, G.; Weigend, M. Towards a new classification of tribe Stachydeae (Lamiaceae): Naming clades using molecular evidence. Bot. J. Linn. Soc. 2019, 190, 345–358. [Google Scholar] [CrossRef]
- Javaid, M.M.; Florentine, S.K.; Ali, H.H.; Chauhan, B.S. Environmental factors affecting the germination and emergence of white horehound (Marrubium vulgare L.): A weed of arid-zone areas. Rangel. J. 2018, 40, 47–54. [Google Scholar] [CrossRef]
- Thanos, C.A.; Kadis, C.C.; Skarou, F. Ecophysiology of germination in the aromatic plants thyme, savory and oregano (Labiatae). Seed Sci. Res. 1995, 5, 161–170. [Google Scholar] [CrossRef]
- Krichen, K.; Mariem, H.B.; Chaieb, M. Ecophysiological requirements on seed germination of a Mediterranean perennial grass (Stipa tenacissima L.) under controlled temperatures and water stress. S. Afr. J. Bot. 2014, 94, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Zou, T.X.; Dai, T.B.; Jiang, D. Effects of nitrogen and potassium application levels on flag leaf photosynthetic characteristics after anthesis in winter wheat. Acta Agron. Sin. 2007, 33, 1667–1673. [Google Scholar]
- Kunzova, E.; Hejcman, M. Yield development of winter wheat over 50 years of FYM, N, P and K fertilizer application on black earth soil in the Czech Republic. Field Crops Res. 2009, 111, 226–234. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, J.; Chen, L.; Ji, A.; Zhao, H.; Guo, P. Effects of fertilizer levels and plant density on chlorophyll contents, its fluorescence and grain yield of Setaria italica. Intern. J. Agric. Biol. 2018, 20, 737–744. [Google Scholar] [CrossRef]
- Zhang, Y. Response characteristics of plant growth and leaf photochemical activity of sugar beet seedlings to different nitrogen application leaves. J. Nuclear Agric. Sci. 2013, 27, 1391–1400. [Google Scholar] [CrossRef]
- Xiong, Z.R.C.; Zhongxiao, Y.; Xiahong, L.; Jing, L.; Minijia, H. Effects of nitrogen and density interaction on grain yield and nitrogen use efficiency of Sorghum. Crops 2018, 5, 110–115. [Google Scholar] [CrossRef]
- Xiao, W.X.; Xie, F.T.; Zhang, H.J.; Wang, H.Y.; Wang, H. Effect of fertilizer and planting density on photosynthetic characteristics and yield of super-high- yielding soybean cultivar. Chin. J. Oil Crop Sci. 2009, 31, 190–195. [Google Scholar]
- Najm, A.; Haj, M.R.; Hadi, S.; Fazeli, F.; Darzi, M.; Rahi, A. Effect of integrated management of nitrogen fertilizer and cattle manure on the leaf chlorophyll, yield, and tuber glycoalkaloids of agria potato. Commun. Soil Sci. Plant Anal. 2012, 43, 912–923. [Google Scholar] [CrossRef]
- Kuo, Y.J. Effects of fertilizer type on chlorophyll content and plant biomass in common Bermuda grass. Afr. J. Agric. Res. 2015, 10, 3997–4000. [Google Scholar] [CrossRef] [Green Version]
- Jafar, S.; Mehdi, T.M. Reducing nitrogen fertilizers with chlorophyll meter for determining health production. J. Env. Sci. Tech. 2020, 22, 103–111. [Google Scholar] [CrossRef]
- Špulák, O.; Martincová, J.; Vitamvas, J.; Kunes, I. Effect of fertilization on chlorophyll activity, content of photosynthetically active pigments and nutrients in Carpathian birch leaves. Austrian J. For. Sci. 2014, 131, 23–44. Available online: https://www.researchgate.net/publication/269630072_Effect_of_fertilization_on_chlorophyll_activity_content_of_photosynthetically_active_pigments_and_nutrients_in_Carpathian_birch_leaves (accessed on 30 December 2021).
- Fiorentini, M.; Zenobi, S.; Giorgini, E.; Basili, D.; Conti, C.; Pro, C.; Monaci, E.; Orsini, R. Nitrogen and chlorophyll status determination in durum wheat as influenced by fertilization and soil management: Preliminary results. PLoS ONE 2019, 14, e0225126. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio R Fd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Organic Matter (%) | Soluble Salts (mS/cm) | CaCO3 (%) | Mechanical Composition | ||
|---|---|---|---|---|---|---|
| Sand (%) | Silt (%) | Clay (%) | ||||
| 8.12 | 0.36 | 0.35 | 5.50 | 56.00 | 28.00 | 16.00 |
| Macronutrient concentrations (ppm) | ||||||
| N-No3 | P | K | Mg | Ca | ||
| 8.00 | 8.00 | 104.00 | 842.00 | >2000 | ||
| Micronutrient concentrations (ppm) | ||||||
| Fe | Zn | Mn | Cu | |||
| 4.7 | 2.00 | 7.06 | 0.77 | |||
| Marrubium Species | matK | rbcL | trnH/psbA | 18S–26S | petB/petD | rpoC1 |
|---|---|---|---|---|---|---|
| M. vulgare | 9, 9, 9, 9, and 12 (5 accessions) | 6, 6, 18, 18, and 8X7 bp (12 accessions) | 7, 12 | − | − | − |
| M. crassidens | 4 | 3 and a deletion of 25 | − | − | − | |
| M. incanum | 5 | − | − | − | ||
| M. peregrinum | 9 | 5 | 7 | − | − | − |
| M. alysson | 2 | − | − | − | ||
| M. thessalum | 9 | − | − | − |
| Characteristics | Control | INM | ChF |
|---|---|---|---|
| Shoot height (cm) | 35.75 ± 7.55 a | 43.06 ± 5.37 a | 37.69 ± 5.94 a |
| Root collar diameter (mm) | 4.03 ± 0.39 b | 4.57 ± 0.40 a | 4.54 ± 0.26 a |
| Number of leaves | 39.25 ± 7.36 c | 70.12 ± 5.19 a | 59.37± 6.09 b |
| Leaf length (cm) | 3.12 ± 0.42 a | 3.04 ± 0.40 a | 3.14 ± 0.73 a |
| Leaf width (cm) | 2.66 ± 0.35 a | 2.50 ± 0.29 a | 2.71 ± 0.49 a |
| Root dry biomass (g) | 1.10 ± 0.19 a | 1.00 ± 0.12 a | 0.69 ± 0.16 b |
| Above ground dry biomass (g) | 3.24 ± 0.49 b | 4.29 ± 0.60 a | 4.05 ± 0.73 ab |
| Photosynthetic rate (μmol m−2 s−1) | 3.48 ± 0.28 c | 4.15 ± 0.49 b | 7.18 ± 0.40 a |
| Chlorophyll content | 19.16 ± 2.35 b | 26.49 ± 1.69 a | 24.93 ± 2.31 a |
| Chlorophyll fluorescence (Fv/Fm) | 0.719 ± 0.028 a | 0.741 ± 0.017 a | 0.725 ± 0.013 a |
| Treatment | N (%) | P (mg/gr) | K (mg/gr) | Ca (mg/gr) | Mg (mg/gr) | Na (mg/gr) |
|---|---|---|---|---|---|---|
| Control | 1.84 c | 2.40 a | 18.87 c | 9.54 c | 5.09 a | 3.74 a |
| Chemical fertilizer | 2.68 b | 2.38 a | 19.81 b | 8.70 b | 4.65 b | 3.00 c |
| Integrated nutrient management | 3.03 a | 2.38 a | 20.85 a | 10.11 a | 5.51 a | 3.48 b |
| Treatment | Fe (ppm) | Mn (ppm) | Zn (ppm) | Cu (ppm) |
|---|---|---|---|---|
| Control | 485.69 a | 90.21 a | 51.31 a | 7.08 b |
| Chemical fertilizer | 224.73 b | 82.89 a | 31.27 c | 3.53 b |
| Integrated nutrient management | 254.60 b | 55.90 b | 37.50 b | 244.86 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pipinis, E.; Hatzilazarou, S.; Kostas, S.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; et al. Facilitating Conservation and Bridging Gaps for the Sustainable Exploitation of the Tunisian Local Endemic Plant Marrubium aschersonii (Lamiaceae). Sustainability 2022, 14, 1637. https://doi.org/10.3390/su14031637
Pipinis E, Hatzilazarou S, Kostas S, Bourgou S, Megdiche-Ksouri W, Ghrabi-Gammar Z, Libiad M, Khabbach A, El Haissoufi M, Lamchouri F, et al. Facilitating Conservation and Bridging Gaps for the Sustainable Exploitation of the Tunisian Local Endemic Plant Marrubium aschersonii (Lamiaceae). Sustainability. 2022; 14(3):1637. https://doi.org/10.3390/su14031637
Chicago/Turabian StylePipinis, Elias, Stefanos Hatzilazarou, Stefanos Kostas, Soumaya Bourgou, Wided Megdiche-Ksouri, Zeineb Ghrabi-Gammar, Mohamed Libiad, Abdelmajid Khabbach, Mohamed El Haissoufi, Fatima Lamchouri, and et al. 2022. "Facilitating Conservation and Bridging Gaps for the Sustainable Exploitation of the Tunisian Local Endemic Plant Marrubium aschersonii (Lamiaceae)" Sustainability 14, no. 3: 1637. https://doi.org/10.3390/su14031637