
Effects of Chinese Milk Vetch Returning on Soil Properties, Microbial Community, and Rice Yield in Paddy Soil
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Site Characterization
2.2. Experiment Design and Field Management
2.3. Sampling and Laboratory Procedures
2.4. DNA Extraction, PCR Amplification, and MiSeq Sequencing of the Soil
2.5. Statistical Analyses
3. Results
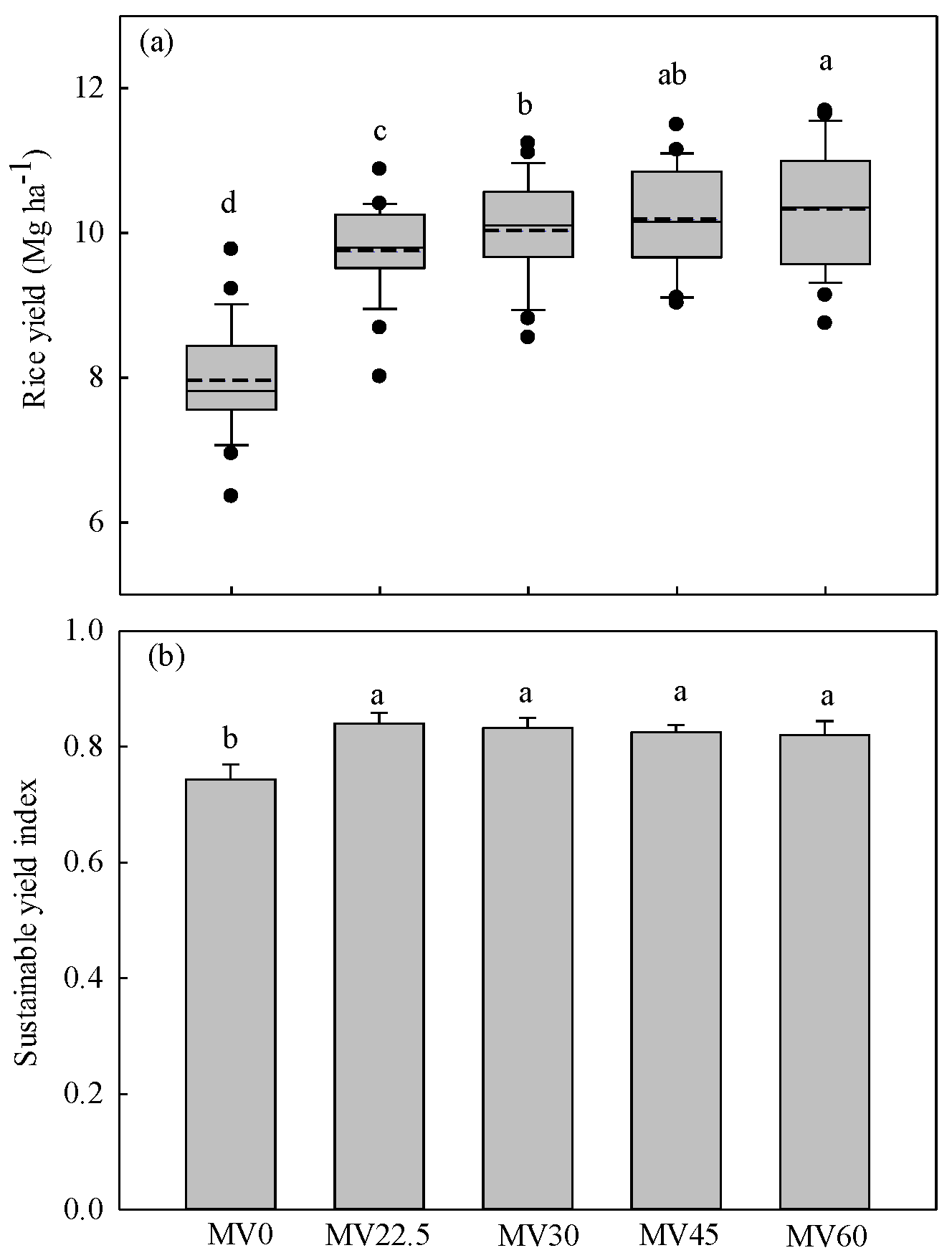
3.1. Rice Yield and Sustainability Yield Index
3.2. Soil Physicochemical Properties
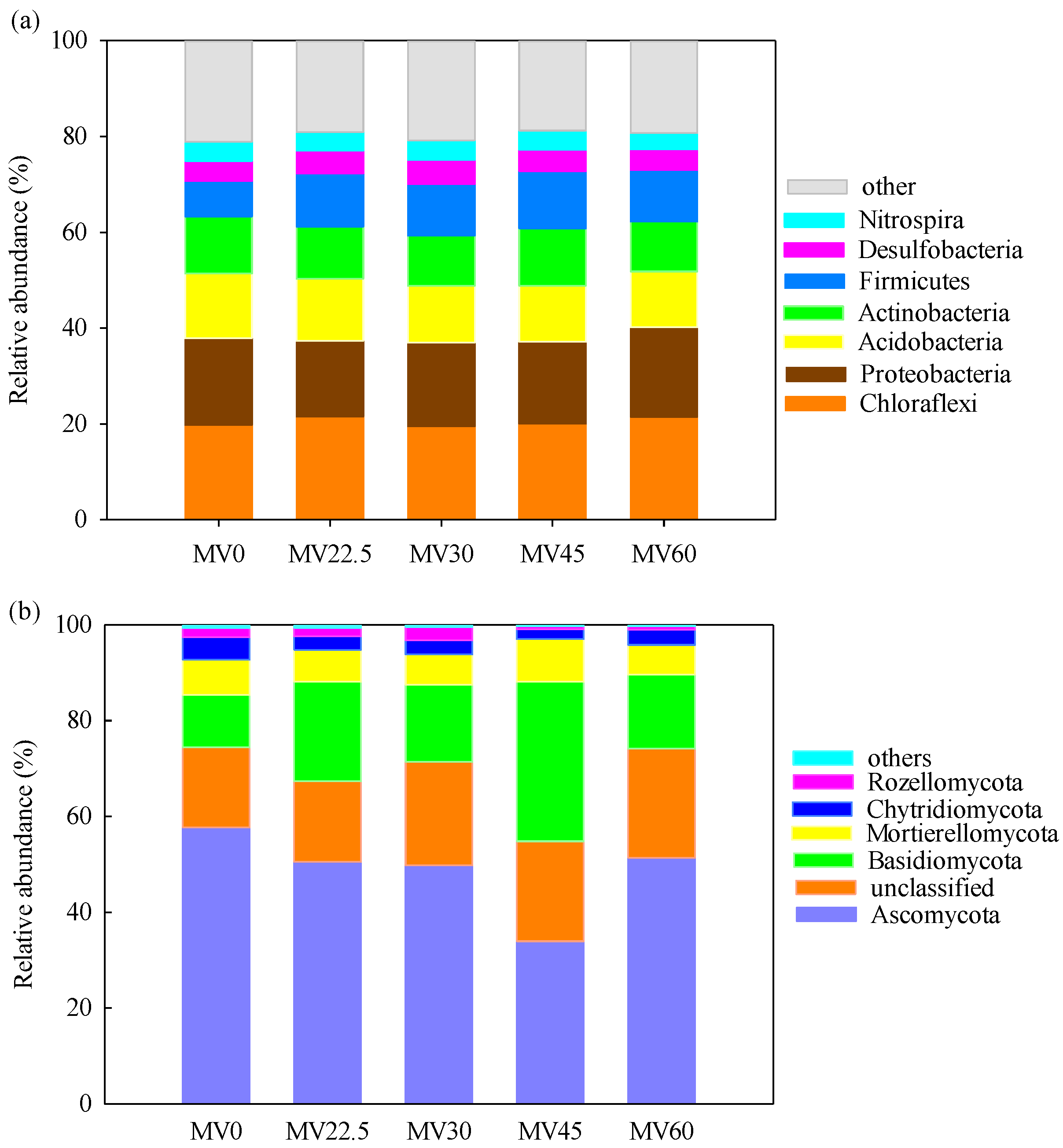
3.3. Soil Microbial Community Structure
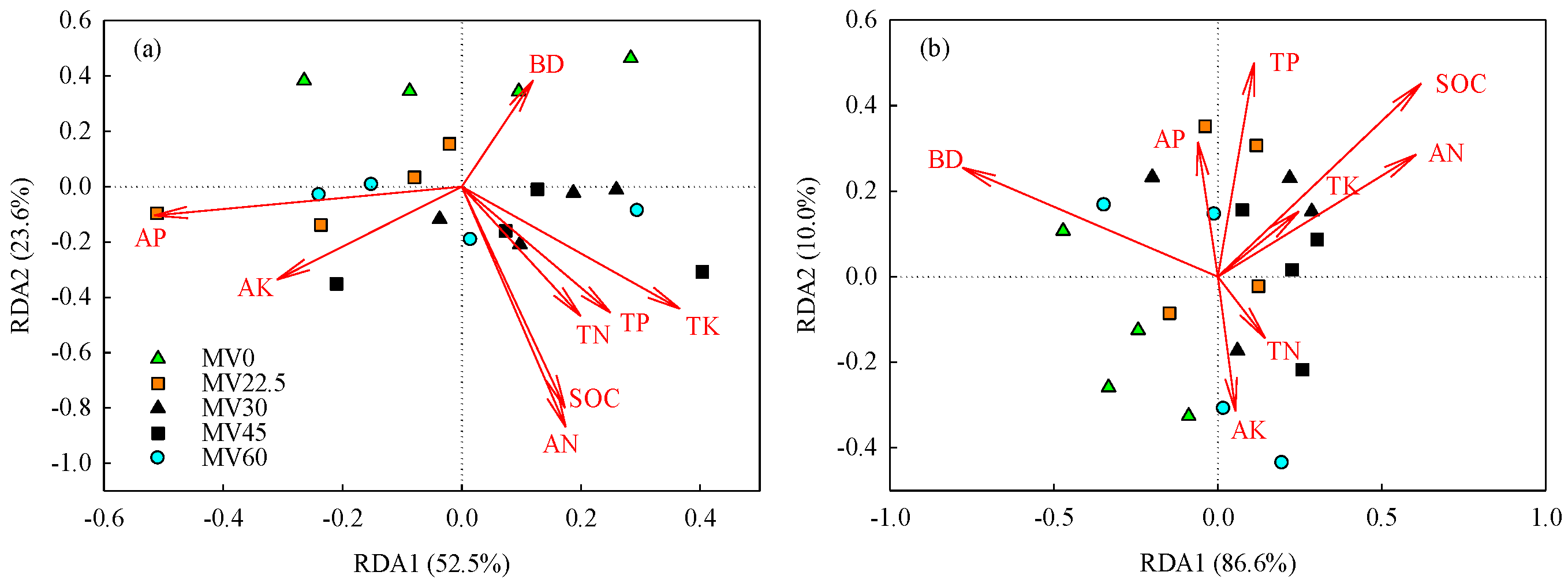
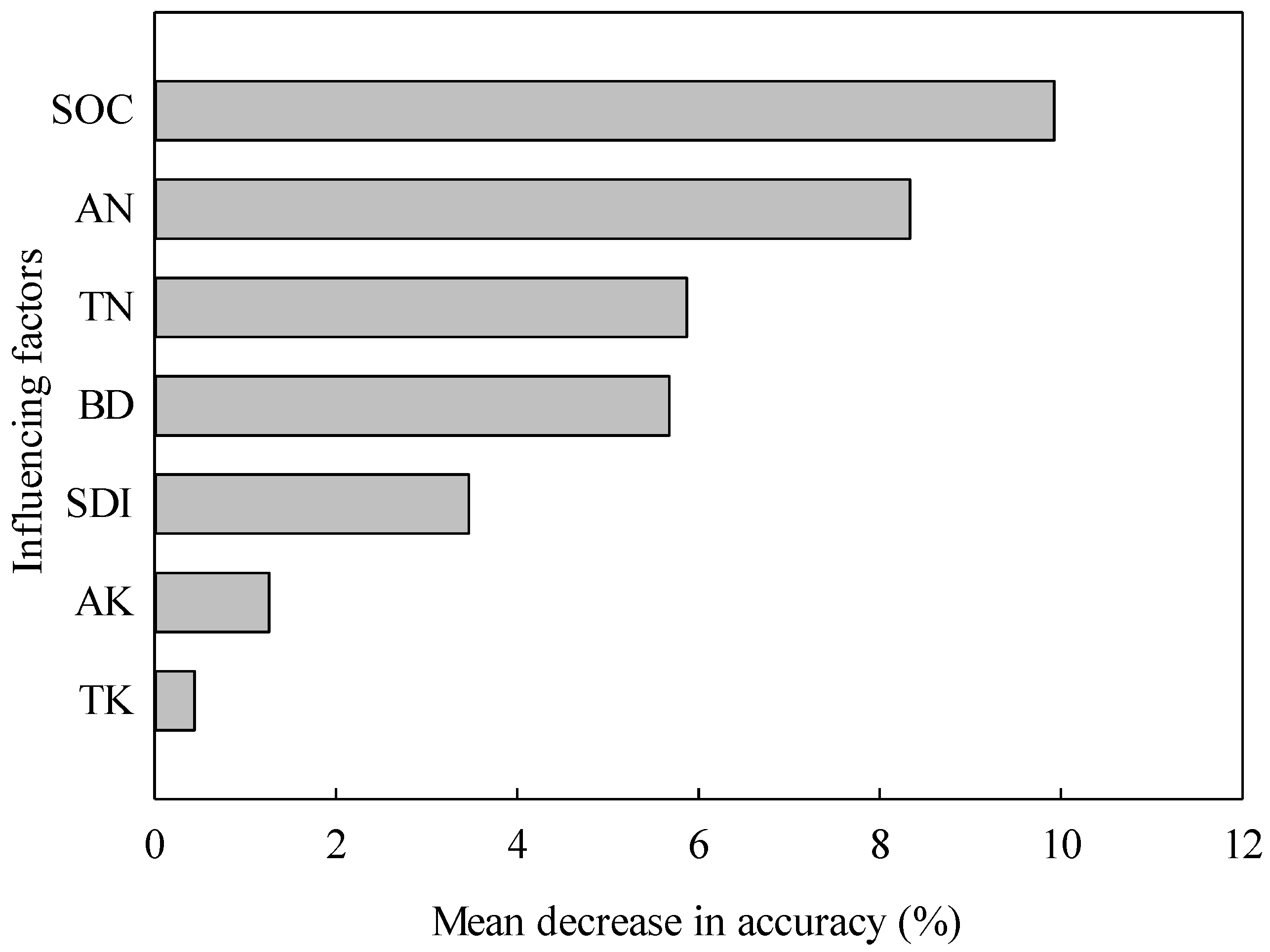
3.4. Relationships between Soil Properties, Microbial Community, and Rice Yield
4. Discussion
4.1. Effect of Varied MV Rates on Soil Physicochemical Properties
4.2. Effect of Varied MV Rates on Soil Microbial Community Structure
4.3. Relationships among Soil Properties, Microbial Community, and Rice Yield
4.4. Management of MV in Rice Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO (Food and Agriculture Organization of the United Nations). FAOSTAT: FAO Statistical Databases. 2019. Available online: http://faostat.fao.org/default.aspx (accessed on 21 December 2021).
- Li, M.; Fu, Q.; Singh, V.P.; Liu, D.; Li, T.; Li, J. Sustainable management of land, water, and fertilizer for rice production considering footprint family assessment in a random environment. J. Clean. Prod. 2020, 258, 120785. [Google Scholar] [CrossRef]
- Ying, H.; Yin, Y.L.; Zheng, H.F.; Wang, Y.C.; Zhang, Q.S.; Xue, Y.F.; Stefanovski, D.; Cui, Z.L.; Dou, Z.X. Newer and select maize, wheat, and rice varieties can help mitigate N footprint while producing more grain. Global Chang. Biol. 2019, 25, 4273–4281. [Google Scholar] [CrossRef] [PubMed]
- Chai, Q.; Nemecek, T.; Liang, C.; Zhao, C.; Yu, A.Z.; Coulter, J.A.; Wang, Y.F.; Hu, F.L.; Wang, L.; Siddique, K.H.M.; et al. Integrated farming with intercropping increases food production while reducing environmental footprint. Proc. Natl. Acad. Sci. USA 2021, 118, 1–12. [Google Scholar] [CrossRef]
- Han, X.; Hu, C.; Chen, Y.; Qiao, Y.; Liu, D.; Fan, J.; Li, S.; Zhang, Z. Crop yield stability and sustainability in a rice-wheat cropping system based on 34-year field experiment. Eur. J. Agron. 2020, 113, 125965. [Google Scholar] [CrossRef]
- Yadav, S.K.; Benbi, D.K. Effect of long-term application of rice straw, farmyard manure and inorganic fertilizer on potassium dynamics in soil. Arch. Agron. Soil Sci. 2018, 65, 374–384. [Google Scholar]
- Baldi, E.; Cavani, L.; Margon, A.; Quartieri, M.; Sorrenti, G.; Marzadori, C.; Toselli, M. Effect of compost application on the dynamics of carbon in a nectarine orchard ecosystem. Sci. Total Environ. 2018, 637, 918–925. [Google Scholar] [CrossRef]
- Qaswar, M.; Liu, Y.R.; Huang, J.; Liu, K.L.; Mudasir, M.; Lv, Z.Z.; Hou, H.Q.; Lan, X.J.; Ji, J.H.; Ahmed, W.; et al. Soil nutrients and heavy metal availability under long-term combined application of swine manure and synthetic fertilizers in acidic paddy soil. J. Soils Sediments 2020, 20, 2093–2106. [Google Scholar] [CrossRef]
- Li, Y.; Bai, N.; Tao, Z.; Mi, X.; He, G.; Wang, Z. Rethinking application of animal manure for wheat production in China. J. Clean. Prod. 2021, 318, 128473. [Google Scholar] [CrossRef]
- Tao, J.; Liu, X.; Liang, Y.; Niu, J.; Xiao, Y.; Gu, Y.; Ma, L.; Meng, D.; Zhang, Y.; Huang, W.; et al. Maize growth responses to soil microbes and soil properties after fertilization with different green manures. Appl. Microbiol. Biot. 2017, 101, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.J.; Cao, W.D.; Gao, J.S.; Huang, J.; Shimizu, K. Effects of long-term application of different green manures on ferric iron reduction in a red paddy soil in southern China. J. Integr. Agric. 2017, 16, 959–966. [Google Scholar] [CrossRef]
- Xie, Z.; Tu, S.; Shah, F.; Xu, C.; Chen, J.; Han, D.; Liu, G.; Li, H.; Muhammad, I.; Cao, W. Substitution of fertilizer-N by green manure improves the sustainability of yield in double-rice cropping system in south China. Field Crop. Res. 2016, 188, 142–149. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, Y.H.; Liao, Y.L.; Zhu, Q.D.; Cheng, H.D.; Nie, X.; Cao, W.D.; Nie, J. Substitution of chemical fertilizer by Chinese milk vetch improves the sustainability of yield and accumulation of soil organic carbon in a double-rice cropping system. J. Integr. Agric. 2019, 18, 2381–2392. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Tiemann, L.K.; Grandy, A.S. Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol. Appl. 2014, 24, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaswar, M.; Huang, J.; Ahmed, W.; Liu, S.; Li, D.; Zhang, L.; Liu, L.; Xu, X.; Han, T.; Du, J.; et al. Substitution of inorganic nitrogen fertilizer with green manure (GM) increased yield stability by improving C input and nitrogen recovery efficiency in rice based cropping system. Agronomy 2019, 9, 609. [Google Scholar] [CrossRef] [Green Version]
- Salangi, F.; Bai, J.; Gao, S.; Yang, L.; Zhou, G.; Cao, W. Improved accumulation capabilities of phosphorus and potassium in green manures and its relationship with soil properties and enzymatic activities. Agronomy 2019, 9, 708. [Google Scholar] [CrossRef] [Green Version]
- Li, F.C.; Wang, Z.H.; Dai, J.; Li, Q.; Wang, X.; Xue, C.; Liu, H.; He, G. Fate of nitrogen from green manure, straw, and fertilizer applied to wheat under different summer fallow management strategies in dryland. Biol. Fertil. Soils 2015, 51, 769–780. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.; Xu, J.; Cao, K.; Wang, J.; Xu, C.; Cao, W. Green manure incorporation with reductions in chemical fertilizer inputs improves rice yield and soil organic matter accumulation. J. Soils Sediments 2020, 20, 2784–2793. [Google Scholar] [CrossRef]
- You, Y.; Ren, J.; Wu, J.; Ma, Z.; Gu, Y.; Wang, Y.; Wang, Z.; Bowatte, S.; Zhou, Q.; Hou, F. Forage taste agents modifying yak grazing decrease soil microbial diversity in alpine meadow. Appl. Soil Ecol. 2021, 168, 104160. [Google Scholar] [CrossRef]
- Wan, P.; Peng, H.; Ji, X.; Chen, X.; Zhou, H. Effect of stand age on soil microbial communities of a plantation Ormosia hosiei forest in southern China. Ecol. Inform. 2021, 62, 101282. [Google Scholar] [CrossRef]
- Zheng, W.; Gong, Q.; Zhao, Z.; Liu, J.; Zhai, B.; Wang, Z.; Li, Z. Changes in the soil bacterial community structure and enzyme activities after intercrop mulch with cover crop for eight years in an orchard. Eur. J. Soil Biol. 2018, 86, 34–41. [Google Scholar] [CrossRef]
- He, H.B.; Li, W.X.; Zhang, Y.W.; Cheng, J.K.; Jia, X.Y.; Li, S.; Yang, H.H.; Chen, B.M.; Xin, G.R. Effects of Italian ryegrass residues as green manure on soil properties and bacterial communities under an Italian ryegrass (Lolium multiflorum L.)-rice (Oryza sativa L.) rotation. Soil Tillage Res. 2020, 196, 104487. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Proshad, R.; Zhang, D.; Uddin, M.; Wu, Y. Presence of cadmium and lead in tobacco and soil with ecological and human health risks in Sichuan province, China. Environ. Sci. Pollut. Res. 2020, 27, 18355–18370. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, D.; Peterson, G.A.; Pratt, P.F. Lithium sodium and potassium. In Methods of Soil Analysis Part II; Chemical and Microbiological, Properties; Pages, A.L., Miller, R.H., Dennis, R.K., Eds.; American Society of Agronomy: Medison, WL, USA, 1982; pp. 225–246. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Government Printing Office: Washington, DC, USA, 1954.
- Liu, Y.M.; Cao, W.Q.; Chen, X.X.; Yu, B.G.; Zou, C.Q. The responses of soil enzyme activities, microbial biomass and microbial community structure to nine years of varied zinc application rates. Sci. Total Environ. 2020, 737, 140245. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.; Wang, Y.; Ye, Y.; Zhao, Y.; Chu, X. Effects of continuous nitrogen fertilizer application on the diversity and composition of rhizosphere soil bacteria. Front. Microbiol. 2020, 11, 1948. [Google Scholar] [CrossRef]
- Chen, J.R.; Qin, W.J.; Chen, X.F.; Cao, W.D.; Qian, G.M.; Liu, J.; Xu, C. Application of Chinese milk vetch affects rice yield and soil productivity in a subtropical double-rice cropping system. J. Integr. Agric. 2020, 19, 2116–2126. [Google Scholar] [CrossRef]
- Jin, K.; White, P.J.; Whalley, W.R.; Shen, J.; Shi, L. Shaping an optimal soil by root-soil interaction. Trends Plant Sci. 2017, 22, 823–829. [Google Scholar] [CrossRef]
- Singh, K.; Choudhary, O.P.; Singh, H.P.; Singh, A.; Mishra, S.K. Sub-soiling improves productivity and economic returns of cotton-wheat cropping system. Soil Tillage Res. 2019, 189, 131–139. [Google Scholar] [CrossRef]
- Adekiya, A.O.; Agbede, T.M.; Aboyeji, C.M.; Oaniran, A.F.; Aremu, C.; Ejue, W.S.; Iranloye, Y.M.; Adegbite, K. Poultry and green manures effects on soil properties, and sorghum performance, and quality. Commun. Soil Sci. Plan. 2022, 53, 463–477. [Google Scholar] [CrossRef]
- Wang, L.; Sun, X.; Li, S.; Zhang, T.; Zhang, W.; Zhai, P. Application of organic amendments to a coastal saline soil in north China: Effects on soil physical and chemical properties and tree growth. PLoS ONE 2014, 9, e89185. [Google Scholar] [CrossRef] [Green Version]
- Xiu, L.; Zhang, W.; Sun, Y.; Wu, D.; Meng, J.; Chen, W. Effects of biochar and straw returning on the key cultivation limitations of albic soil and soybean growth over 2 years. Catena 2019, 173, 481–493. [Google Scholar] [CrossRef]
- Sun, Y.N.; Huang, S.; Yu, X.C.; Zhang, W.J. Stability and saturation of soil organic carbon in rice fields: Evidence from a long-term fertilization experiment in subtropical China. J. Soils Sediments 2013, 13, 1327–1334. [Google Scholar] [CrossRef]
- Li, Z.Q.; Li, D.D.; Ma, L.; Yu, Y.Y.; Zhao, B.Z.; Zhang, J.B. Effects of straw management and nitrogen application rate on soil organic matter fractions and microbial properties in North China Plain. J. Soils Sediments 2019, 19, 618–628. [Google Scholar] [CrossRef]
- Bedadaa, W.; Karltuna, E.; Lemenihb, M.; Tolerab, M. Long-term addition of compost and N P fertilizer increases crop yield and improves soil quality in experiments on small holder farms. Agric. Ecosyst. Environ. 2014, 195, 193–201. [Google Scholar] [CrossRef]
- Yang, L.; Bai, J.S.; Liu, J.; Zeng, N.H.; Cao, W.D. Green manuring effect on changes of soil nitrogen fractions, maize growth, and nutrient uptake. Agronomy 2018, 8, 261. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of carbon reserves is improved by controlled soil-drying during grain filling of wheat. Crop Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Lu, D.; Yue, S.; Lu, F.; Cui, Z.; Liu, Z.; Zou, C.; Chen, X. Integrated crop-N system management to establish high wheat yield population. Field Crop. Res. 2016, 191, 66–74. [Google Scholar] [CrossRef]
- Rodríguez-Berbel, N.; Ortega, R.; Lucas-Borja, M.E.; Sol´e-Benet, A.; Miralles, I. Long-term effects of two organic amendments on bacterial communities of calcareous mediterranean soils degraded by mining. J. Environ. Manag. 2020, 271, 110920. [Google Scholar] [CrossRef]
- Fu, H.; Li, H.; Yin, P.; Mei, H.; Li, J.; Zhou, P.; Wang, Y.; Ma, Q.; Jeyaraj, A.; Kuberan, T.; et al. Integrated Application of Rapeseed Cake and Green Manure Enhances Soil Nutrients and Microbial Communities in Tea Garden Soil. Sustainability 2021, 13, 2967. [Google Scholar] [CrossRef]
- Zhang, Y.; Hao, X.; Alexander, T.W.; Thomas, B.W.; Shi, X.; Lupwayi, N.Z. Long-term and legacy effects of manure application on soil microbial community composition. Biol. Fertil. Soils 2018, 54, 269–283. [Google Scholar] [CrossRef]
- Pan, J.; Shang, Y.; Zhang, W.J.; Chen, X.; Cui, Z. Improving soil quality for higher grain yields in Chinese wheat and maize production. Land Degrad. Dev. 2020, 31, 1125–1137. [Google Scholar] [CrossRef]
- Ai, C.; Zhang, S.; Zhang, X.; Guo, D.; Zhou, W.; Huang, S. Distinct responses of soil bacterial and fungal communities to changes in fertilization regime and crop rotation. Geoderma 2018, 319, 156–166. [Google Scholar] [CrossRef]
- Liang, H.; Wang, X.; Yan, J.; Luo, L. Characterizing the Intra-Vineyard Variation of Soil Bacterial and Fungal Communities. Front. Microbiol. 2019, 10, 1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Liu, X.; Yang, W.; Yang, X.; Li, W.; Xia, Q.; Li, J.; Gao, Z.; Yang, Z. Rhizosphere soil properties, microbial community, and enzyme activities: Short-term responses to partial substitution of chemical fertilizer with organic manure. J. Environ. Manag. 2021, 299, 113650. [Google Scholar] [CrossRef]
- Caban, J.R.; Kuppusamy, S.; Kim, J.H.; Yoon, Y.E.; Kim, S.Y.; Lee, Y.B. Green manure amendment enhances microbial activity and diversity in antibiotic-contaminated soil. Appl. Soil Ecol. 2018, 129, 72–76. [Google Scholar] [CrossRef]
- Gao, S.; Cao, W.; Zhou, G.; Rees, R.M. Bacterial communities in paddy soils changed by milk vetch as green manure: A study conducted across six provinces in South China. Pedosphere 2021, 31, 521–530. [Google Scholar] [CrossRef]
- Yuan, J.; Zhao, M.; Li, R.; Huang, Q.; Rensing, C.; Raza, W.; Shen, Q. Antibacterial compounds-macrolactin alters the soil bacterial community and abundance of the gene encoding PKS. Front. Microbiol. 2016, 7, 1904. [Google Scholar] [CrossRef] [Green Version]
- Pang, G.; Cai, F.; Li, R.X.; Zhao, Z.; Li, R.; Gu, X.L.; Shen, Q.R.; Chen, W. Trichoderma-enriched organic fertilizer can mitigate microbiome degeneration of monocropped soil to maintain better plant growth. Plant Soil 2017, 416, 181–192. [Google Scholar] [CrossRef]
- Kong, J.; He, Z.; Chen, L.; Yang, R.; Du, J. Efficiency of biochar, nitrogen addition, and microbial agent amendments in remediation of soil properties and microbial community in Qilian Mountains mine soils. Ecol. Evol. 2021, 11, 9318–9331. [Google Scholar] [CrossRef]
- Ding, T.; Yan, Z.; Zhang, W.; Duan, T. Green manure crops affected soil chemical properties and fungal diversity and community of apple orchard in the Loess Plateau of China. J. Soil Sci. Plant Nut. 2021, 21, 1089–1102. [Google Scholar] [CrossRef]
- Wang, J.T.; Zheng, Y.M.; Hu, H.W.; Zhang, L.M.; Li, J.; He, J.Z. Soil pH determines the alpha diversity but not beta diversity of soil fungal community along altitude in a typical Tibetan forest ecosystem. J. Soils Sediments 2015, 15, 1224–1232. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Zhou, B.; Zhao, B.; Ma, M.; Guan, D.; Li, J.; Chen, S.; Cao, F.; Shen, D.; et al. Thirty-four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast china. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]
- Wang, J.; Liu, G.; Zhang, C.; Wang, G.; Cui, Y. Higher temporal turnover of soil fungi than bacteria during long-term secondary succession in a semiarid abandoned farmland. Soil Tillage Res. 2019, 194, 104305. [Google Scholar] [CrossRef]
- Zhou, G.; Gao, S.; Lu, Y.; Liao, Y.; Nie, J.; Cao, W. Co-incorporation of green manure and rice straw improves rice production, soil chemical, biochemical and microbiological properties in a typical paddy field in southern China. Soil Tillage Res. 2020, 197, 104499. [Google Scholar] [CrossRef]
- Wang, C.; Li, H.; Sun, X.; Cai, T. Responses of soil microbial biomass and enzyme activities to natural restoration of reclaimed temperate marshes after abandonment. Front. Environ. Sci. 2021, 9, 701610. [Google Scholar] [CrossRef]
- Xiang, X.; Liu, J.; Zhang, J.; Li, D.; Kuzyakov, Y. Divergence in fungal abundance and community structure between soils under long-term mineral and organic fertilization. Soil Tillage Res. 2020, 196, 104491. [Google Scholar] [CrossRef]
- He, X.; Xie, H.; Gao, D.; Khashi, U.; Rahman, M.; Zhou, X.; Wu, F. Biochar and intercropping with potato–onion enhanced the growth and yield advantages of tomato by regulating the soil properties, nutrient uptake, and soil microbial community. Front. Microbiol. 2021, 12, 695447. [Google Scholar] [CrossRef]
- Chen, L.; Xu, H.; Sun, J.; Baoyin, T. The short-term impacts of soil disturbance on soil microbial community in a degraded Leymus chinensis steppe, North China. Soil Tillage Res. 2021, 213, 105112. [Google Scholar] [CrossRef]
- Wang, Z.T.; Li, T.; Li, Y.Z.; Zhao, D.Q.; Han, J.; Liu, Y.; Liao, Y.C. Relationship between the microbial community and catabolic diversity in response to conservation tillage. Soil Tillage Res. 2020, 196, 104431. [Google Scholar] [CrossRef]
- Liu, J.; Jia, X.; Yan, W.; Zhong, Y.; Shangguan, Z. Changes in soil microbial community structure during long-term secondary succession. Land Degrad. Dev. 2020, 31, 1151–1166. [Google Scholar] [CrossRef]
- Gao, S.J.; Zhang, R.G.; Cao, W.D.; Fan, Y.Y.; Gao, J.S.; Jing, H.; Bai, J.S.; Zeng, N.H.; Chang, D.N.; Katsu-Yoshi, S.; et al. Long-term rice-rice-green manure rotation changing the microbial communities in typical red paddy soil in South China. J. Integr. Agric. 2015, 14, 2512–2520. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BD (g cm−3) | SOC (g kg−1) | TN (g kg−1) | TP (g kg−1) | TK (g kg−1) | AN (mg kg−1) | AP (mg kg−1) | AK (mg kg−1) | |
|---|---|---|---|---|---|---|---|---|
| MV0 | 1.24 ± 0.04 a | 10.6 ± 0.25 c | 1.07 ± 0.06 c | 0.21 ± 0.03 a | 7.04 ± 0.49 c | 46.0 ± 5.13 c | 6.23 ± 0.76 a | 50.56 ± 2.14 b |
| MV22.5 | 1.22 ± 0.03 ab | 12.0 ± 0.94 b | 1.08 ± 0.08 c | 0.23 ± 0.02 a | 7.44 ± 0.52 bc | 62.5 ± 9.06 b | 6.74 ± 0.24 a | 53.08 ± 2.99 b |
| MV30 | 1.20 ± 0.04 ab | 13.0 ± 0.94 ab | 1.18 ± 0.05 b | 0.23 ± 0.03 a | 7.36 ± 0.08 bc | 86.0 ± 11.1 a | 6.09 ± 0.64 a | 50.98 ± 3.13 b |
| MV45 | 1.16 ± 0.04 b | 13.1 ± 0.20 a | 1.31 ± 0.07 a | 0.25 ± 0.04 a | 8.13 ± 0.59 a | 88.8 ± 7.18 a | 6.39 ± 0.81 a | 52.82 ± 4.82 b |
| MV60 | 1.18 ± 0.05 b | 12.9 ± 0.57 ab | 1.37 ± 0.09 a | 0.22 ± 0.04 a | 7.93 ± 0.39 ab | 77.7 ± 5.59 b | 6.61 ± 0.39 a | 59.28 ± 4.54 a |
| ANOVA | ns | *** | *** | ns | * | *** | ns | * |
| Treatment | Bacteria | Fungi | ||
|---|---|---|---|---|
| Chao Richness | Shannon Diversity | Chao Richness | Shannon Diversity | |
| MV0 | 4518.9 ± 318.3 a | 7.02 ± 0.09 a | 1257.7 ± 192.8 a | 4.53 ± 0.62 a |
| MV22.5 | 4329.6 ± 96.8 a | 6.93 ± 0.02 ab | 1236.6 ± 141.7 a | 4.42 ± 0.67 a |
| MV30 | 4149.3 ± 316.8 a | 6.85 ± 0.13 ab | 1236.0 ± 419.1 a | 4.54 ± 0.41 a |
| MV45 | 4125.7 ± 338.5 a | 6.80 ± 0.16 b | 1206.3 ± 124.2 a | 4.19 ± 0.80 a |
| MV60 | 4170.8 ± 361.6 a | 6.86 ± 0.09 ab | 1030.5 ± 180.8 a | 4.13 ± 0.43 a |
| ANOVA | ns | ns | ns | ns |
| Yield | SOC | TN | TP | TK | AN | AP | AK | BD | Chao (B) | Shannon (B) | Chao (F) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOC | 0.69 ** | |||||||||||
| TN | 0.75 ** | 0.39 | ||||||||||
| TP | 0.29 | 0.16 | 0.12 | |||||||||
| TK | 0.57 ** | 0.32 | 0.62 ** | 0.04 | ||||||||
| AN | 0.71 ** | 0.62 ** | 0.51 * | 0.23 | 0.39 | |||||||
| AP | −0.02 | 0.20 | 0.21 | 0.02 | 0.22 | −0.02 | ||||||
| AK | 0.47 * | 0.10 | 0.30 | −0.07 | 0.08 | 0.18 | −0.25 | |||||
| BD | −0.47 * | −0.61 ** | −0.44 | −0.08 | −0.34 | −0.31 | −0.18 | −0.02 | ||||
| Chao (B) | −0.32 | −0.33 | −0.44 | 0.15 | −0.51 * | −0.28 | −0.04 | 0.19 | 0.46 * | |||
| Shannon (B) | −0.50 * | −0.44 | −0.48 * | 0.03 | −0.58 ** | −0.41 | −0.01 | 0.14 | 0.52 * | 0.87 ** | ||
| Chao (F) | −0.17 | −0.04 | −0.37 | 0.20 | −0.27 | 0.10 | −0.25 | −0.02 | 0.12 | 0.35 | 0.29 | |
| Shannon (F) | −0.30 | −0.31 | −0.43 | 0.18 | −0.16 | −0.03 | −0.31 | −0.11 | 0.10 | 0.15 | 0.08 | 0.56 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zheng, C.; Zhang, L.; Zhang, C.; Lv, Y.; Nie, L.; Zhang, X.; Li, B.; Cao, W.; Li, M.; et al. Effects of Chinese Milk Vetch Returning on Soil Properties, Microbial Community, and Rice Yield in Paddy Soil. Sustainability 2022, 14, 16065. https://doi.org/10.3390/su142316065
Zhang J, Zheng C, Zhang L, Zhang C, Lv Y, Nie L, Zhang X, Li B, Cao W, Li M, et al. Effects of Chinese Milk Vetch Returning on Soil Properties, Microbial Community, and Rice Yield in Paddy Soil. Sustainability. 2022; 14(23):16065. https://doi.org/10.3390/su142316065
Chicago/Turabian StyleZhang, Jishi, Chunfeng Zheng, Lin Zhang, Chenglan Zhang, Yuhu Lv, Liangpeng Nie, Xiangning Zhang, Benyin Li, Weidong Cao, Min Li, and et al. 2022. "Effects of Chinese Milk Vetch Returning on Soil Properties, Microbial Community, and Rice Yield in Paddy Soil" Sustainability 14, no. 23: 16065. https://doi.org/10.3390/su142316065