
Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Retrieval and Identification of Sugarcane PHT Genes
2.2. Sequence Analysis of PHT Proteins from Sugarcane
2.3. Phylogenetic Analysis of PHT Genes
2.4. Gene Structure Analysis and Identification of Conserved Motifs
2.5. Prediction of Structure, Protein Pocket Sites, and Protein Modeling
2.6. GO and KEGG Analysis
2.7. Chromosomal Organization and Ka/Ks Calculation
2.8. Expression Study of PHT Genes in Sugarcane
2.9. Prediction of Putative cis-Regulatory Elements and Transient Expression Analysis of EaPHT1:2 Promoter
3. Results
3.1. Identification and Characterization of PHT Genes in Sugarcane
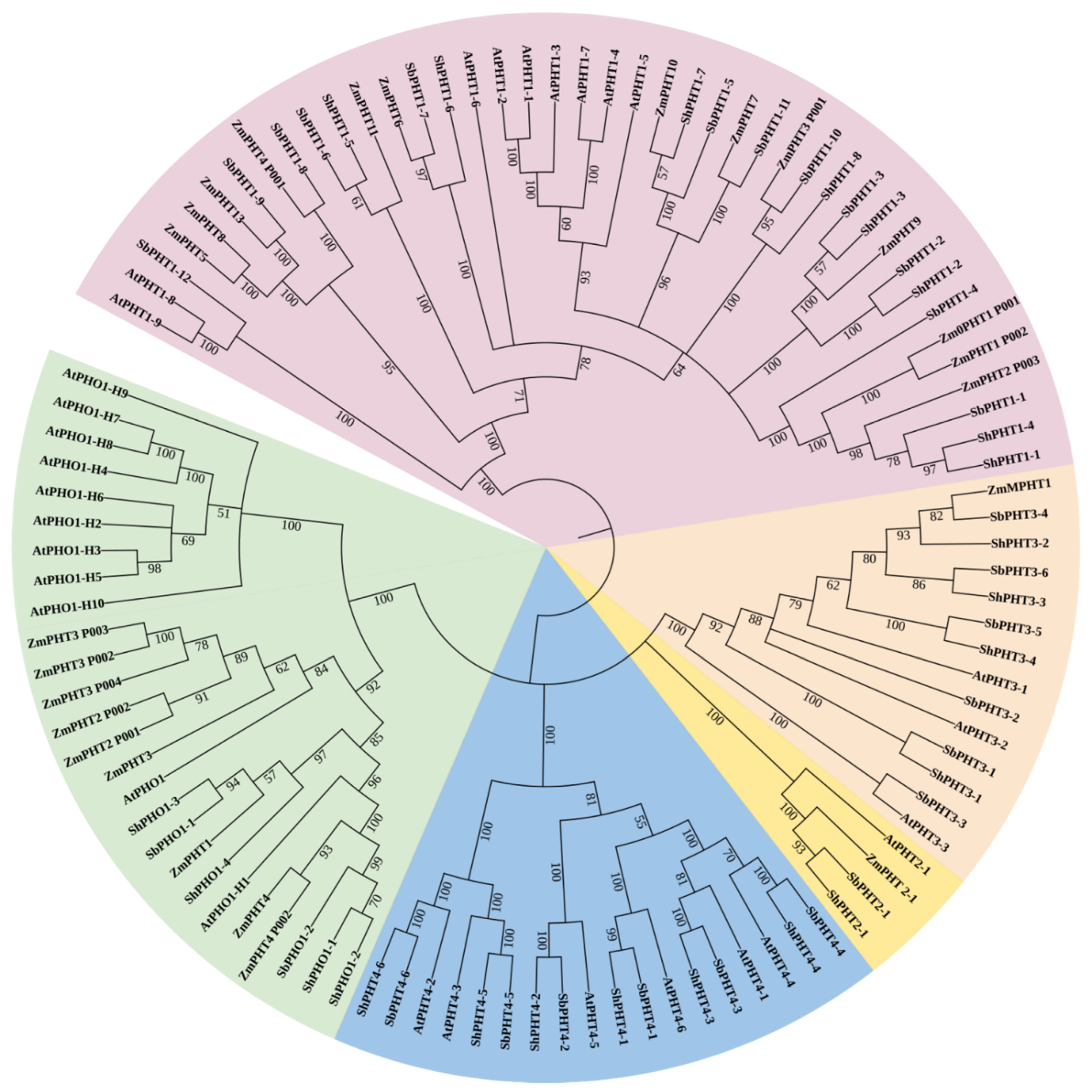
3.2. Phylogenetic Analysis
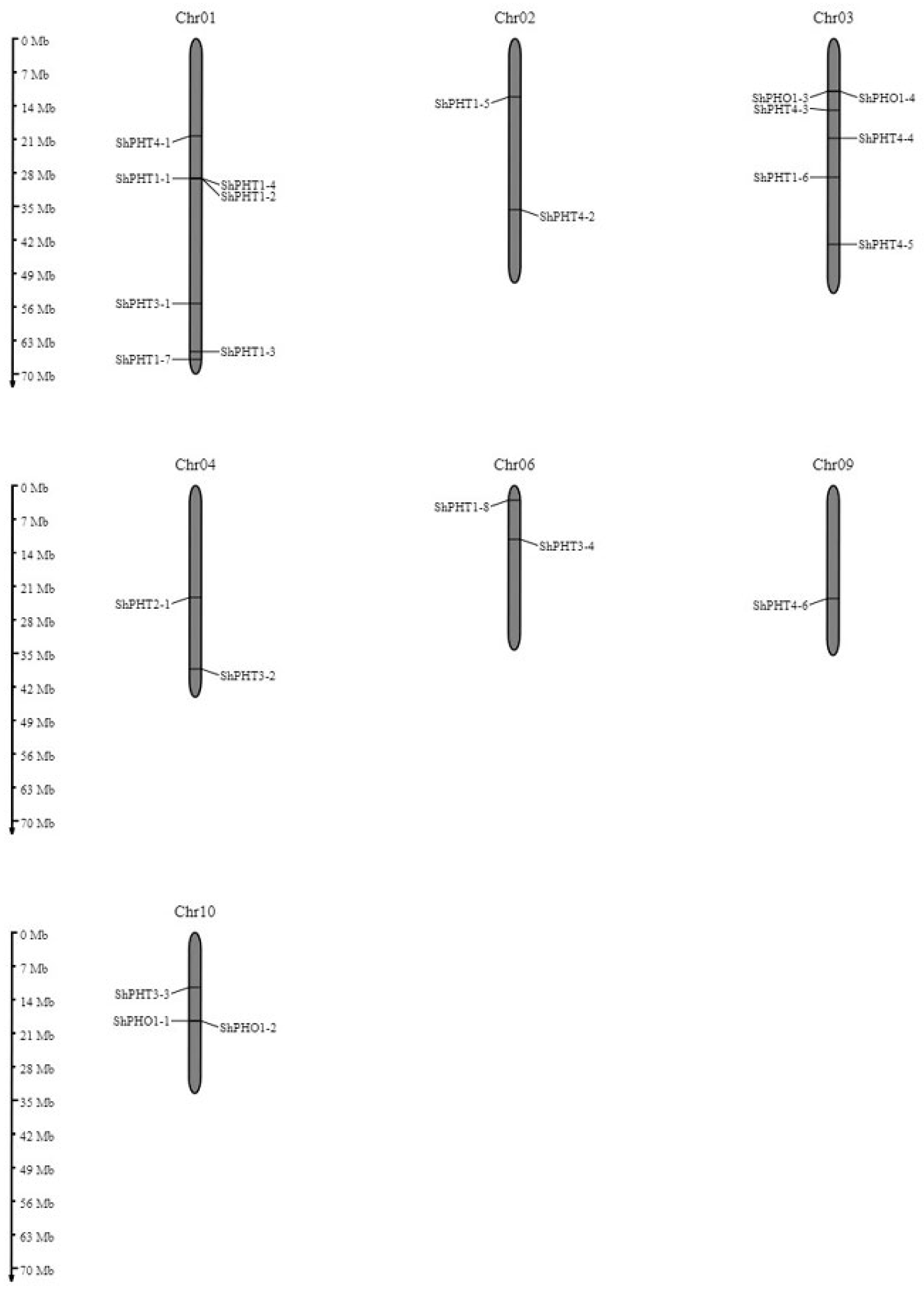
3.3. Chromosomal Localization and Duplication of ShPHT Genes
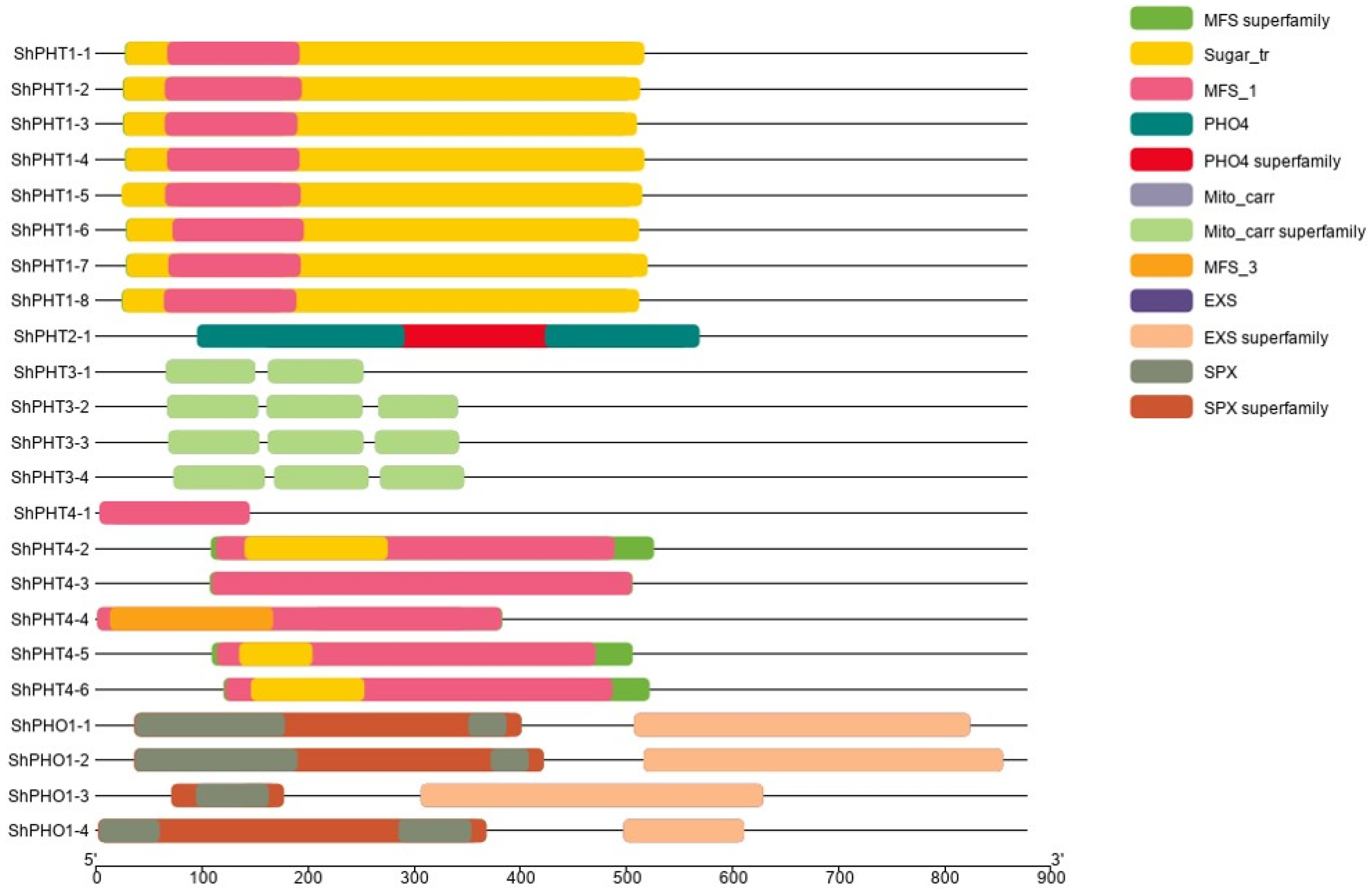
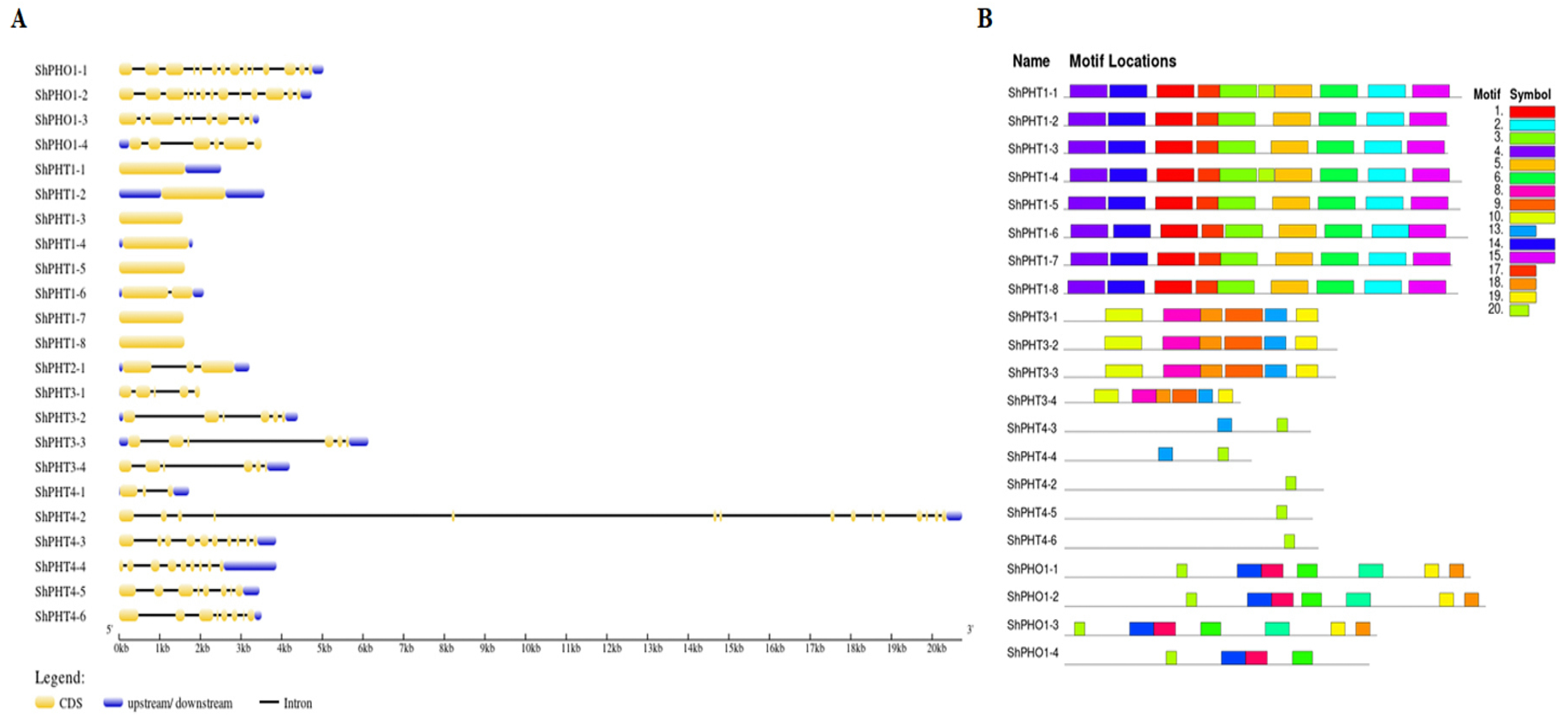
3.4. Gene Structure and Conserved Motif Analysis
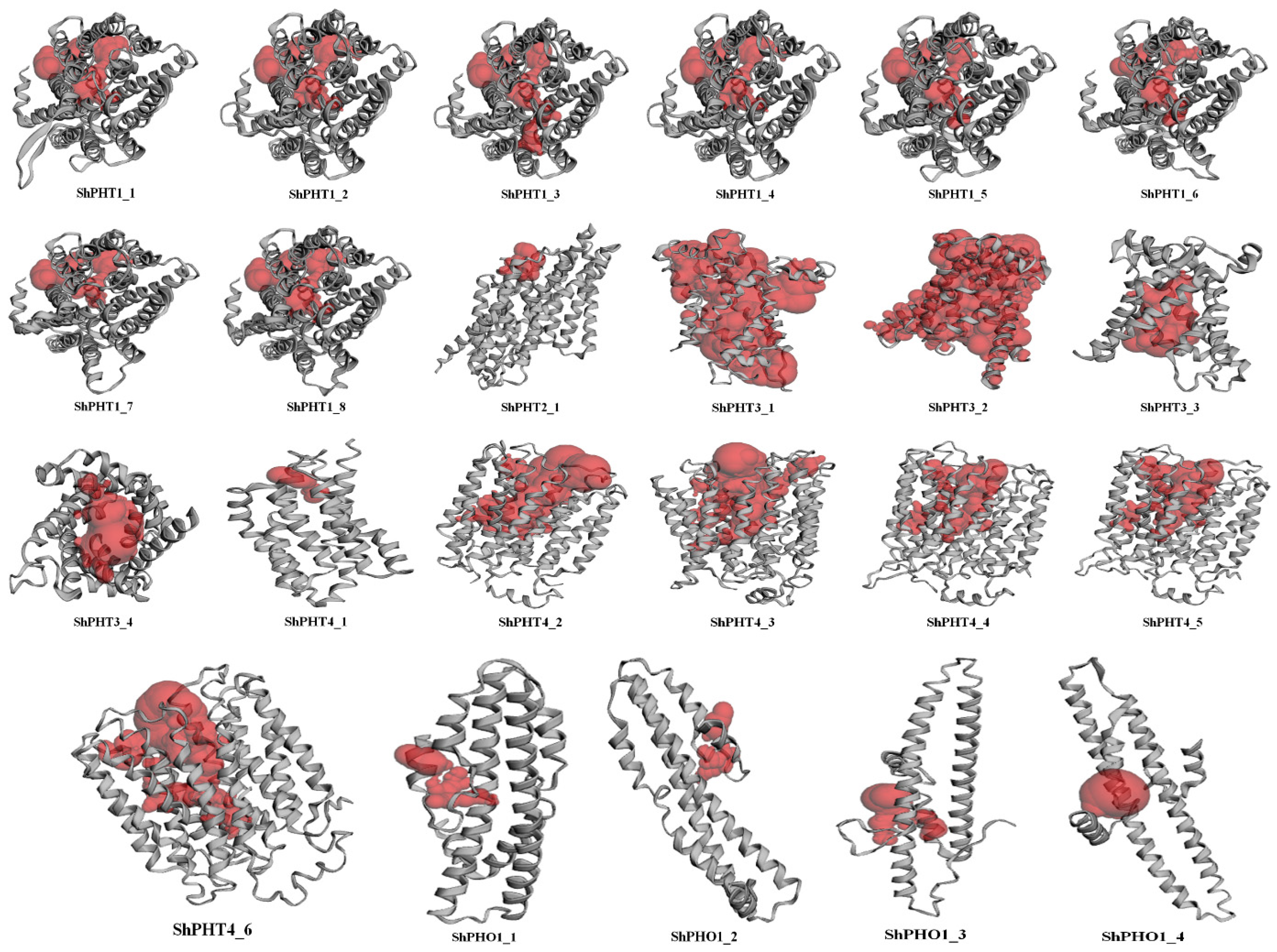
3.5. Structure and Protein Pocket Sites Prediction
3.6. Post-Translational Modifications, Transmembrane Domains (TMD), and Subcellular Localization of ShPHT Proteins
3.7. GO and KEGG Analysis
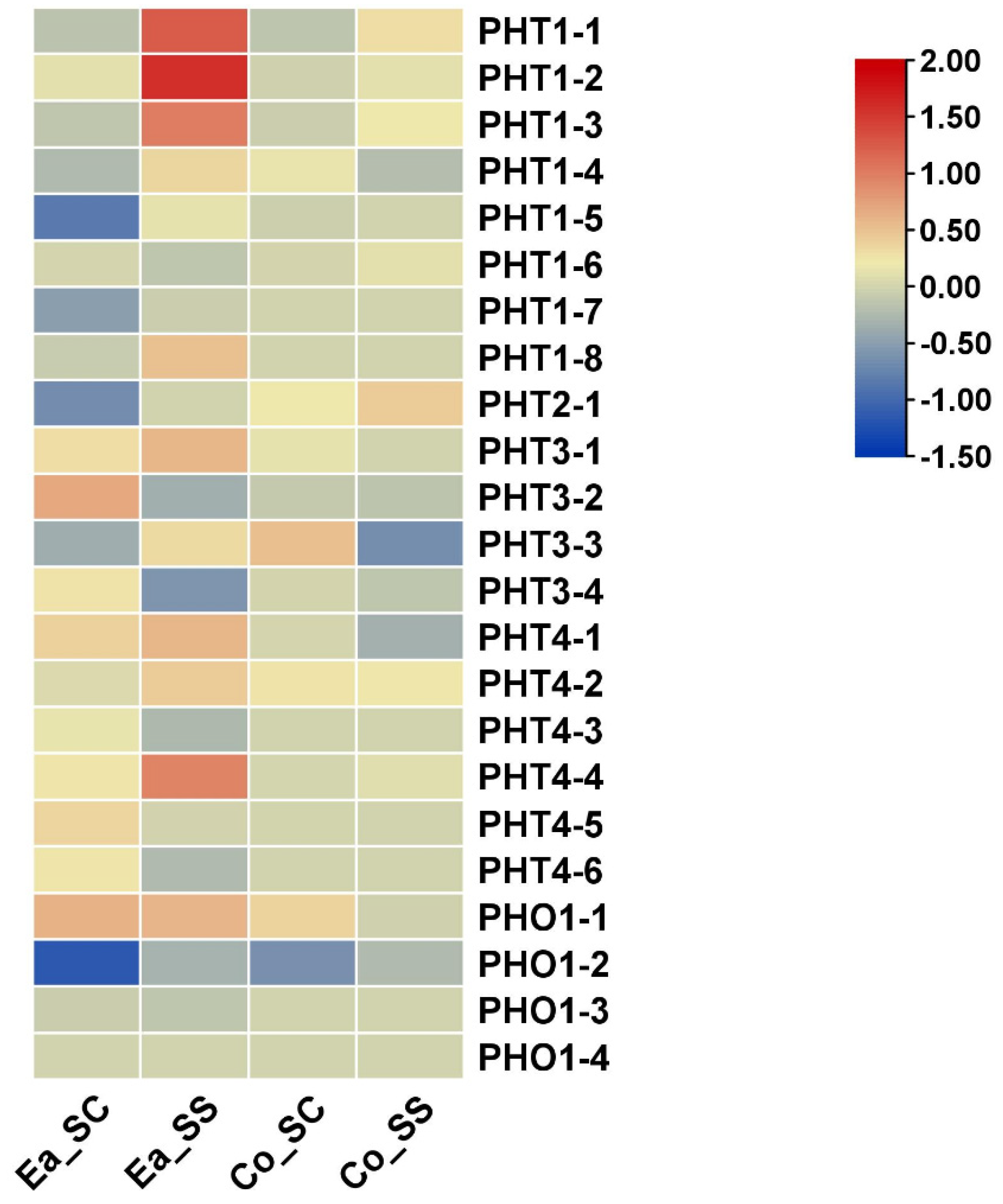
3.8. Expression Analysis of PHT Genes under Salinity Stress
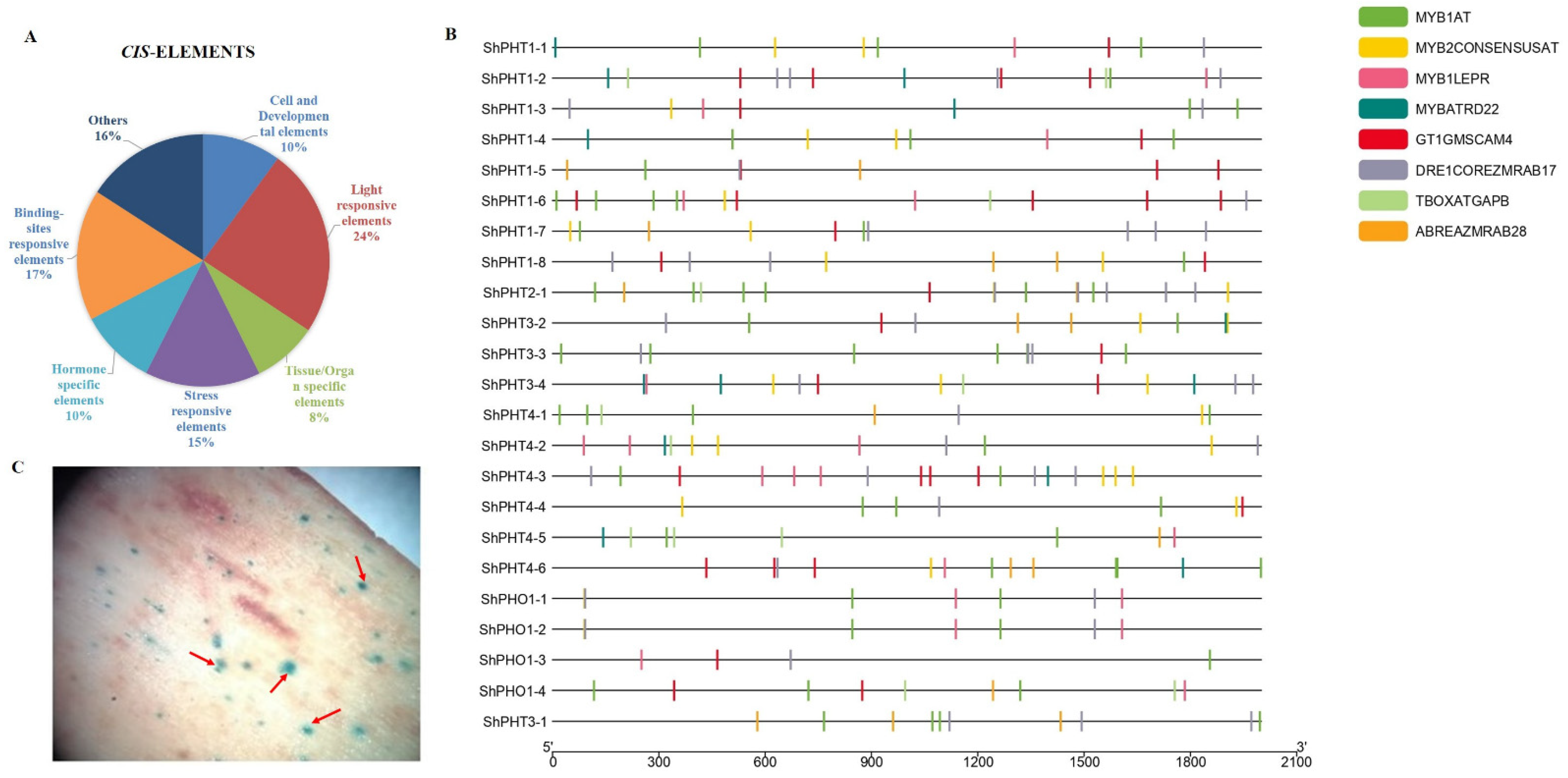
3.9. Putative cis-Regulatory Element Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, W.; Nadeem, M.; Ashiq, W.; Zaeem, M.; Gilani, S.S.M.; Rajabi-Khamseh, S.; Pham, T.H.; Kavanagh, V.; Thomas, R.; Cheema, M. The Effects of Organic and Inorganic Phosphorus Amendments on the Biochemical Attributes and Active Microbial Population of Agriculture Podzols Following Silage Corn Cultivation in Boreal Climate. Sci. Rep. 2019, 9, 17297. [Google Scholar] [CrossRef] [Green Version]
- Bindraban, P.S.; Dimkpa, C.O.; Pandey, R. Exploring Phosphorus Fertilizers and Fertilization Strategies for Improved Human and Environmental Health. Biol. Fertil. Soils 2020, 56, 299–317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liao, H.; Lucas, W.J. Molecular Mechanisms Underlying Phosphate Sensing, Signaling, and Adaptation in Plants. J. Integr. Plant Biol. 2014, 56, 192–220. [Google Scholar] [CrossRef]
- Bechtaoui, N.; Rabiu, M.K.; Raklami, A.; Oufdou, K.; Hafidi, M.; Jemo, M. Phosphate-Dependent Regulation of Growth and Stresses Management in Plants. Front. Plant Sci. 2021, 12, 679916. [Google Scholar] [CrossRef]
- Młodzińska, E.; Zboińska, M. Phosphate Uptake and Allocation—A Closer Look at Arabidopsis thaliana L. and Oryza Sativa L. Front. Plant Sci. 2016, 7, 1198. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, D.; Rezzonico, E.; MacDonald-Comber Petétot, J.; Somerville, C.; Poirier, Y. Identification and Characterization of the Arabidopsis PHO1 Gene Involved in Phosphate Loading to the Xylem. Plant Cell 2002, 14, 889–902. [Google Scholar] [CrossRef] [Green Version]
- Lhamo, D.; Shao, Q.; Tang, R.; Luan, S. Genome-Wide Analysis of the Five Phosphate Transporter Families in Camelina sativa and Their Expressions in Response to Low-P. Int. J. Mol. Sci. 2020, 21, 8365. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Liao, L.; Xu, J.; Liang, X.; Liu, W. Genome-Wide Identification and Functional Characterization of the Phosphate Transporter Gene Family in Sorghum. Biomolecules 2019, 9, 670. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Xu, Y.; Jiang, H.; Jiang, C.; Du, Y.; Gong, C.; Wang, W.; Zhu, S.; Han, G.; Cheng, B. Systematic Identification, Evolution and Expression Analysis of the Zea mays PHT1 Gene Family Reveals Several New Members Involved in Root Colonization by Arbuscular Mycorrhizal Fungi. Int. J. Mol. Sci. 2016, 17, 930. [Google Scholar] [CrossRef] [Green Version]
- Hasanzadeh, S.; Faraji, S.; Abdullah; Heidari, P. Phosphate Transporter Genes: Genome-Wide Identification and Characterization in Camelina sativa. bioRxiv 2021. [Google Scholar] [CrossRef]
- He, M.; He, C.-Q.; Ding, N.-Z. Abiotic Stresses: General Defenses of Land Plants and Chances for Engineering Multistress Tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [Green Version]
- Dey, G.; Banerjee, P.; Sharma, R.K.; Maity, J.P.; Etesami, H.; Shaw, A.K.; Huang, Y.-H.; Huang, H.-B.; Chen, C.-Y. Management of Phosphorus in Salinity-Stressed Agriculture for Sustainable Crop Production by Salt-Tolerant Phosphate-Solubilizing Bacteria—A Review. Agronomy 2021, 11, 1552. [Google Scholar] [CrossRef]
- Baek, D.; Chun, H.J.; Yun, D.-J.; Kim, M.C. Cross-Talk between Phosphate Starvation and Other Environmental Stress Signaling Pathways in Plants. Mol. Cells 2017, 40, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Formann, S.; Hahn, A.; Janke, L.; Stinner, W.; Sträuber, H.; Logroño, W.; Nikolausz, M. Beyond Sugar and Ethanol Production: Value Generation Opportunities Through Sugarcane Residues. Front. Energy Res. 2020, 8, 579577. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Soltangheisi, A.; Withers, P.J.A.; Pavinato, P.S.; Cherubin, M.R.; Rossetto, R.; Do Carmo, J.B.; da Rocha, G.C.; Martinelli, L.A. Improving Phosphorus Sustainability of Sugarcane Production in Brazil. GCB Bioenergy 2019, 11, 1444–1455. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-59259-890-8. [Google Scholar]
- Sonnhammer, E.L.L. A Hidden Markov Model for Predicting Transmembrane Helices in Protein Sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1998, 6, 175–182. [Google Scholar]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of Post-Translational Glycosylation and Phosphorylation of Proteins from the Amino Acid Sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s Conserved Domain Database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The Protein Families Database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools—An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPMA: Significant Improvements in Protein Secondary Structure Prediction by Consensus Prediction from Multiple Alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Dundas, J.; Ouyang, Z.; Tseng, J.; Binkowski, A.; Turpaz, Y.; Liang, J. CASTp: Computed Atlas of Surface Topography of Proteins with Structural and Topographical Mapping of Functionally Annotated Residues. Nucleic Acids Res. 2006, 34, W116–W118. [Google Scholar] [CrossRef] [Green Version]
- Willard, L.; Ranjan, A.; Zhang, H.; Monzavi, H.; Boyko, R.F.; Sykes, B.D.; Wishart, D.S. VADAR: A Web Server for Quantitative Evaluation of Protein Structure Quality. Nucleic Acids Res. 2003, 31, 3316–3319. [Google Scholar] [CrossRef] [Green Version]
- Törönen, P.; Medlar, A.; Holm, L. PANNZER2: A Rapid Functional Annotation Web Server. Nucleic Acids Res. 2018, 46, W84–W88. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust Conversion of Protein Sequence Alignments into the Corresponding Codon Alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.; Conery, J.S. The Evolutionary Fate and Consequences of Duplicate Genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Vignesh, P.; Mahadevaiah, C.; Parimalan, R.; Valarmathi, R.; Dharshini, S.; Nisha, S.; Suresha, G.S.; Swathi, S.; Mahadeva Swamy, H.K.; Sreenivasa, V.; et al. Comparative de Novo Transcriptome Analysis Identifies Salinity Stress Responsive Genes and Metabolic Pathways in Sugarcane and Its Wild Relative Erianthus Arundinaceus [Retzius] Jeswiet. Sci. Rep. 2021, 11, 24514. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Philip, A.; Syamaladevi, D.P.; Chakravarthi, M.; Gopinath, K.; Subramonian, N. 5′ Regulatory Region of Ubiquitin 2 Gene from Porteresia Coarctata Makes Efficient Promoters for Transgene Expression in Monocots and Dicots. Plant Cell Rep. 2013, 32, 1199–1210. [Google Scholar] [CrossRef]
- Rae, A.L.; Cybinski, D.H.; Jarmey, J.M.; Smith, F.W. Characterization of Two Phosphate Transporters from Barley; Evidence for Diverse Function and Kinetic Properties among Members of the Pht1 Family. Plant Mol. Biol. 2003, 53, 27–36. [Google Scholar] [CrossRef]
- Liu, F.; Chang, X.-J.; Ye, Y.; Xie, W.-B.; Wu, P.; Lian, X.-M. Comprehensive Sequence and Whole-Life-Cycle Expression Profile Analysis of the Phosphate Transporter Gene Family in Rice. Mol. Plant 2011, 4, 1105–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Wang, X.; Hu, R.; Wang, Y.; Xiao, C.; Jiang, Y.; Zhang, X.; Zheng, C.; Fu, Y.-F. The Pattern of Phosphate Transporter 1 Genes Evolutionary Divergence in Glycine MaxL. BMC Plant Biol 2013, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Chen, X.; Wang, H.; Liao, D.; Gu, M.; Qu, H.; Sun, S.; Xu, G. Genome-Wide Investigation and Expression Analysis Suggest Diverse Roles and Genetic Redundancy of Pht1 Family Genes in Response to Pi Deficiency in Tomato. BMC Plant Biol. 2014, 14, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceasar, S.A.; Hodge, A.; Baker, A.; Baldwin, S.A. Phosphate Concentration and Arbuscular Mycorrhizal Colonisation Influence the Growth, Yield and Expression of Twelve PHT1 Family Phosphate Transporters in Foxtail Millet (Setaria Italica). PLoS ONE 2014, 9, e108459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Meng, S.; Li, M.; Zhao, Z. Genomic Identification and Expression Analysis of the Phosphate Transporter Gene Family in Poplar. Front. Plant Sci. 2016, 7, 1398. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zhao, S.; Wu, X.; Wang, X.; Nan, Y.; Wang, D.; Chen, Q. Identification and Characterization of Phosphate Transporter Genes in Potato. J. Biotechnol. 2017, 264, 17–28. [Google Scholar] [CrossRef]
- Teng, W.; Zhao, Y.-Y.; Zhao, X.-Q.; He, X.; Ma, W.-Y.; Deng, Y.; Chen, X.-P.; Tong, Y.-P. Genome-Wide Identification, Characterization, and Expression Analysis of PHT1 Phosphate Transporters in Wheat. Front. Plant Sci. 2017, 8, 543. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Li, M.; Shao, Y.; Yu, L.; Ma, F. Comprehensive Genomic Identification and Expression Analysis of the Phosphate Transporter (PHT) Gene Family in Apple. Front. Plant Sci. 2017, 8, 426. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, X.; Zhang, H.; Wang, S.; Ye, X.; Shi, L.; Xu, F.; Ding, G. Molecular Identification of the Phosphate Transporter Family 1 (PHT1) Genes and Their Expression Profiles in Response to Phosphorus Deprivation and Other Abiotic Stresses in Brassica napus. PLoS ONE 2019, 14, e0220374. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhou, J.; Zhou, H.-J.; Wang, M.-M.; Liu, M.-M.; Ke, Y.-Z.; Li, P.-F.; Li, J.-N.; Du, H. Global Survey and Expressions of the Phosphate Transporter Gene Families in Brassica napus and Their Roles in Phosphorus Response. Int. J. Mol. Sci. 2020, 21, 1752. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Li, G.; Sun, Z.; Chen, Y.; Guo, W.; Li, Y.; Chen, Y.; Yang, J.; Hou, H. Identification, Structure Analysis, and Transcript Profiling of Phosphate Transporters under Pi Deficiency in Duckweeds. Int. J. Biol. Macromol. 2021, 188, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Rawoof, A.; Islam, K.; Momo, J.; Ramchiary, N. Identification and Expression Analysis of Phosphate Transporter Genes and Metabolites in Response to Phosphate Stress in Capsicum annuum. Environ. Exp. Bot. 2021, 190, 104597. [Google Scholar] [CrossRef]
- Lv, S.; Wang, D.; Jiang, P.; Jia, W.; Li, Y. Variation of PHT Families Adapts Salt Cress to Phosphate Limitation under Salinity. Plant Cell Environ. 2021, 44, 1549–1564. [Google Scholar] [CrossRef]
- Cao, D.; Liu, Y.; Linlong, M.; Liu, Z.; Li, J.; Beibei, W.; Zhang, X.; Yin, P.; Jin, X.; Huang, J. Genome-Wide Identification and Characterization of Phosphate Transporter Gene Family Members in Tea Plants (Camellia sinensis L. O. Kuntze) under Different Selenite Levels. Plant Physiol. Biochem. 2021, 166, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Walder, F.; Brulé, D.; Koegel, S.; Wiemken, A.; Boller, T.; Courty, P.-E. Plant Phosphorus Acquisition in a Common Mycorrhizal Network: Regulation of Phosphate Transporter Genes of the Pht1 Family in Sorghum and Flax. New Phytol. 2015, 205, 1632–1645. [Google Scholar] [CrossRef] [PubMed]
- Versaw, W.K.; Harrison, M.J. A Chloroplast Phosphate Transporter, PHT2;1, Influences Allocation of Phosphate within the Plant and Phosphate-Starvation Responses. Plant Cell 2002, 14, 1751–1766. [Google Scholar] [CrossRef] [Green Version]
- Mudge, S.R.; Rae, A.L.; Diatloff, E.; Smith, F.W. Expression Analysis Suggests Novel Roles for Members of the Pht1 Family of Phosphate Transporters in Arabidopsis. Plant J. 2002, 31, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A Conserved MYB Transcription Factor Involved in Phosphate Starvation Signaling Both in Vascular Plants and in Unicellular Algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Xu, Q.; Zhang, F.-C.; Chen, Y.; Li, L.-Q.; Wu, W.-H.; Chen, Y.-F. WRKY42 Modulates Phosphate Homeostasis through Regulating Phosphate Translocation and Acquisition in Arabidopsis. Plant Physiol. 2015, 167, 1579–1591. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, K.; Liu, X.; Ruan, H.; Zheng, M.; Yu, Z.; Gai, J.; Yang, S. Transcription Factor GmWRKY46 Enhanced Phosphate Starvation Tolerance and Root Development in Transgenic Plants. Front. Plant Sci. 2021, 12, 700651. [Google Scholar] [CrossRef]
- Yang, T.; Hao, L.; Yao, S.; Zhao, Y.; Lu, W.; Xiao, K. TabHLH1, a BHLH-Type Transcription Factor Gene in Wheat, Improves Plant Tolerance to Pi and N Deprivation via Regulation of Nutrient Transporter Gene Transcription and ROS Homeostasis. Plant Physiol. Biochem. 2016, 104, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Araceli, O.-A.; Alfredo, C.-R.; Javier, M.-M.; Luis, H.-E. A Phosphate Starvation-Driven Bidirectional Promoter as a Potential Tool for Crop Improvement and in Vitro Plant Biotechnology. Plant Biotechnol. J. 2017, 15, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, Y.; Wang, H.; Ma, X.; Yao, W.; Wang, H. Light and Ethylene Coordinately Regulate the Phosphate Starvation Response through Transcriptional Regulation of Phosphate Starvation Response1. Plant Cell 2017, 29, 2269–2284. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Genomic Position (bp) | Strand | CDS (bp) | Protein Length (A.A) | Protein Molecular Weight (kDa) | Theoretical pI | Number of Negatively Amino Acid | Number of Positively Amino Acid | Instability Index | Aliphatic Index | GRAVY | Trans Membrane Domain |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ShPHT1-1 | Sh_251L03_g000070 | Sh01:30199785–30202295 | + | 1626 | 541 | 58.814 | 7.63 | 40 | 41 | 30.26 | 88.82 | 0.349 | 11 |
| ShPHT1-2 | Sh_251L03_g000050 | Sh01:30228262–30231839 | - | 1575 | 524 | 57.087 | 7.13 | 37 | 37 | 31.84 | 90.97 | 0.368 | 11 |
| ShPHT1-3 | Sh_236J19_g000050 | Sh01:67384705–67386273 | - | 1569 | 522 | 57.224 | 8.92 | 36 | 43 | 27.01 | 94.62 | 0.409 | 11 |
| ShPHT1-4 | Sh_229B19_p000040 | Sh01: 67384705–67386273 | - | 1578 | 525 | 57.512 | 8.7 | 38 | 43 | 28.67 | 93.33 | 0.385 | 11 |
| ShPHT1-5 | Sh_216E19_g000030 | Sh02:12496129–12497748 | - | 1620 | 539 | 58.89 | 8.66 | 47 | 52 | 36.71 | 89.98 | 0.294 | 12 |
| ShPHT1-6 | Sh_247G22_g000050 | Sh03:29909878–29911962 | - | 1656 | 551 | 60.285 | 8.6 | 44 | 48 | 33.2 | 93.9 | 0.336 | 12 |
| ShPHT1-7 | Sh_222J11_g000100 | Sh01:69058018–69059604 | + | 1587 | 528 | 56.698 | 8.1 | 33 | 35 | 38.63 | 89.15 | 0.396 | 11 |
| ShPHT1-8 | Sh_222A01_g000030 | Sh06:3102321–3103931 | - | 1611 | 536 | 58.81 | 8.3 | 40 | 43 | 34.94 | 84.38 | 0.289 | 12 |
| ShPHT2-1 | Sh_232L11_g000050 | Sh04:24145076–24148283 | + | 1719 | 572 | 59.251 | 9.42 | 24 | 36 | 32.21 | 106.54 | 0.707 | 13 |
| ShPHT3-1 | Sh_252F06_g000020 | Sh01:57191013–57193003 | - | 1044 | 347 | 37.76 | 8.94 | 26 | 35 | 23.1 | 92.85 | 0.283 | 0 |
| ShPHT3-2 | Sh_208E04_contig-2_g000020 | Sh04:39635010–39639407 | - | 1119 | 372 | 39.135 | 9.23 | 24 | 35 | 38.3 | 88.23 | 0.259 | 0 |
| ShPHT3-3 | Sh_234H01_g000060 | Sh10:11858441–11864575 | + | 1113 | 370 | 39.113 | 9.22 | 23 | 34 | 44.05 | 86.38 | 0.268 | 0 |
| ShPHT3-4 | Sh_247B01_g000110 | Sh06:11674421–11678619 | + | 1104 | 367 | 38.412 | 9.39 | 20 | 34 | 37.81 | 84.99 | 0.292 | 0 |
| ShPHT4-1 | Sh_227B04_g000030 | Sh01:21002885–21004607 | + | 651 | 216 | 23.926 | 9.73 | 12 | 22 | 39.26 | 105.6 | 0.462 | 5 |
| ShPHT4-2 | Sh_241M07_g000080 | Sh02:36755180–36775918 | - | 1623 | 540 | 57.755 | 6.76 | 38 | 37 | 44.48 | 102.94 | 0.377 | 9 |
| ShPHT4-3 | Sh_227N18_g000090 | Sh03:15436854–15440720 | + | 1542 | 513 | 55.383 | 8.42 | 31 | 34 | 45.05 | 97.31 | 0.346 | 9 |
| ShPHT4-4 | Sh_227D07_g000010 | Sh03:21537139–21541012 | - | 1173 | 390 | 42.392 | 9.27 | 17 | 25 | 48.94 | 107.9 | 0.566 | 9 |
| ShPHT4-5 | Sh_220N05_g000170 | Sh03:44310739–44314195 | - | 1554 | 517 | 55.814 | 10.04 | 19 | 38 | 42.72 | 100.43 | 0.475 | 10 |
| ShPHT4-6 | Sh_217I09_g000040 | Sh09:24432214–24435720 | + | 1590 | 529 | 55.44 | 9.65 | 19 | 35 | 37.44 | 97.2 | 0.615 | 11 |
| ShPHO1-1 | Sh_244C06_g000040 | Sh10:19140506–19145537 | - | 2541 | 846 | 96.534 | 8.65 | 90 | 99 | 41.61 | 89.78 | −0.137 | 6 |
| ShPHO1-2 | Sh_226O09_contig-2_g000020 | Sh10:19140506–19145537 | - | 2634 | 877 | 99.766 | 8.44 | 94 | 101 | 42.78 | 87.5 | −0.162 | 5 |
| ShPHO1-3 | Sh_254O18_g000050 | Sh03:11344984–11348425 | - | 1956 | 651 | 73.3 | 9.1 | 54 | 69 | 41.96 | 92.44 | −0.056 | 5 |
| ShPHO1-4 | Sh_253H03_g000040 | Sh03:11346722–11350230 | - | 1908 | 635 | 70.213 | 8.82 | 65 | 74 | 42.67 | 85.64 | −0.154 | 4 |
| Gene Name | Orthologs | Ka | Ks | Ka/Ks | Divergence Time (Mya) |
|---|---|---|---|---|---|
| ShPHT1-1 | ShPHT1-4 | 0.0027 | 0.0552 | 0.0491 | 4.50 × 1012 |
| ShPHT1-2 | ShPHT1-3 | 0.0864 | 1.4853 | 0.0582 | 1.22 × 1014 |
| ShPHT3-2 | ShPHT3-3 | 0.0875 | 0.7968 | 0.1098 | 6.53 × 1013 |
| ShPHT4-3 | ShPHT4-4 | 0.1088 | 2.9394 | 0.037 | 2.41 × 1014 |
| ShPHT4-5 | ShPHT4-6 | 0.4846 | 4.1984 | 0.1154 | 3.44 × 1014 |
| ShPHO1-1 | ShPHO1-2 | 0.0029 | 0.0154 | 0.1904 | 1.26 × 1012 |
| ShPHO1-3 | ShPHO1-4 | 0.0489 | 0.134 | 0.3651 | 1.10 × 1013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murugan, N.; Palanisamy, V.; Channappa, M.; Ramanathan, V.; Ramaswamy, M.; Govindakurup, H.; Chinnaswamy, A. Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress. Sustainability 2022, 14, 15893. https://doi.org/10.3390/su142315893
Murugan N, Palanisamy V, Channappa M, Ramanathan V, Ramaswamy M, Govindakurup H, Chinnaswamy A. Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress. Sustainability. 2022; 14(23):15893. https://doi.org/10.3390/su142315893
Chicago/Turabian StyleMurugan, Naveenarani, Vignesh Palanisamy, Mahadevaiah Channappa, Valarmathi Ramanathan, Manimekalai Ramaswamy, Hemaprabha Govindakurup, and Appunu Chinnaswamy. 2022. "Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress" Sustainability 14, no. 23: 15893. https://doi.org/10.3390/su142315893