


Vermicomposting as a Sustainable Option for Managing Biomass of the Invasive Tree Acacia dealbata Link

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Vermicomposting Setup
2.3. Data Collection
2.4. Physico-Chemical Analyses
2.5. Microbiological Analyses
2.6. Phytotoxicity Assay
2.7. Statistical Analyses
3. Results
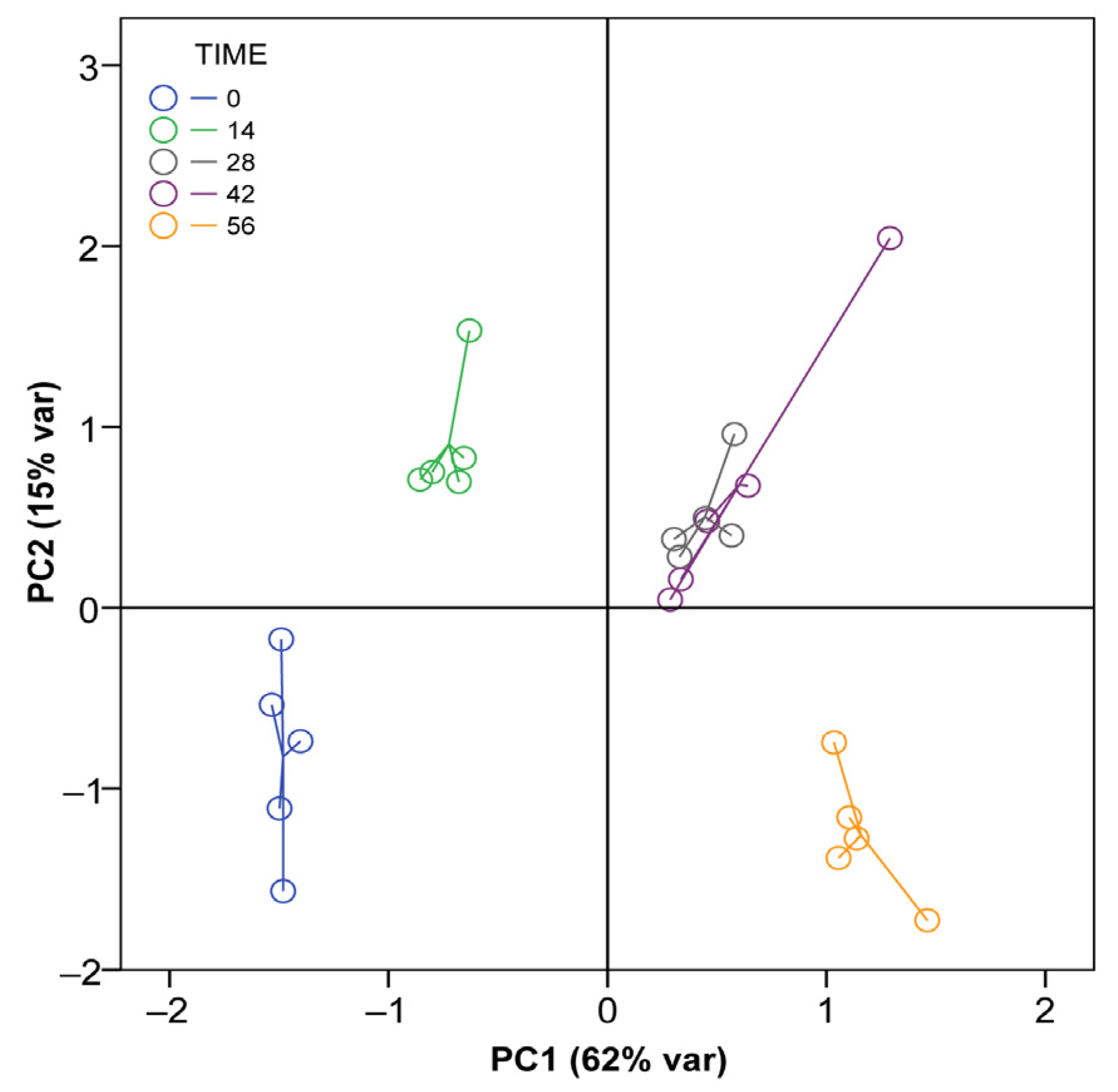
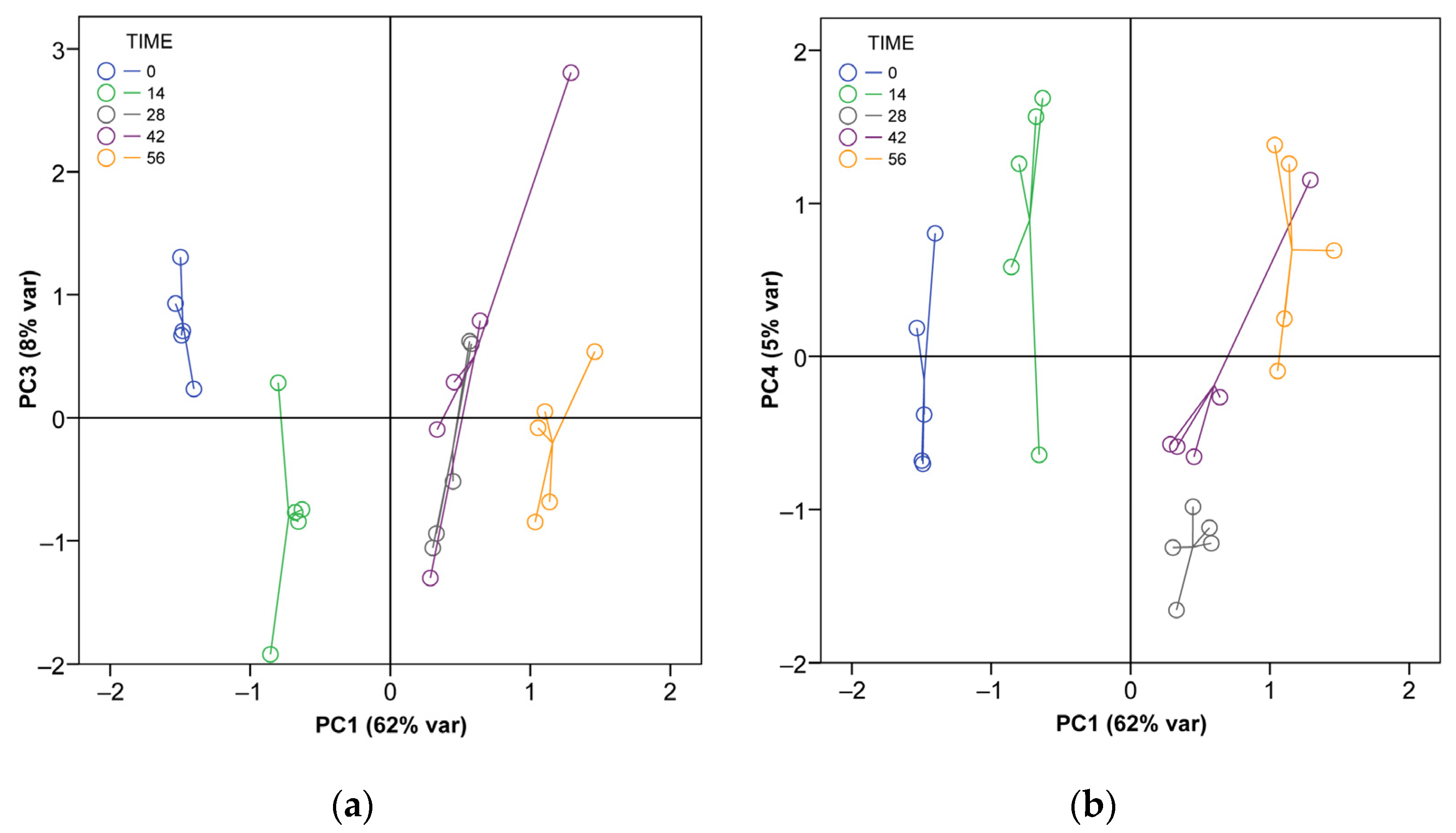
3.1. Changes in the Properties of A. dealbata Vermicompost during Vermicomposting
3.2. Changes in Earthworm Population
3.3. Phytotoxic Effects before and after Vermicomposting
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable (Unit) | Time (Days) | ||||
|---|---|---|---|---|---|
| 0 | 14 | 28 | 42 | 56 | |
| Moisture (%) | 64.23 | 72.08 | 78.63 | 80.7 | 79.56 |
| pH | 5.14 | 7.26 | 7.15 | 6.63 | 6.42 |
| Electrical Conductivity (mS cm−2) | 752.4 | 370.8 | 259 | 263.6 | 211.8 |
| Organic Matter (%) | 96.64 | 96.38 | 92.89 | 93.41 | 89.12 |
| Total C (g kg−1) | 506.2 | 516.3 | 489.2 | 493.6 | 471.1 |
| Total N (g kg−1) | 25.5 | 33.3 | 40.8 | 41.3 | 39.2 |
| C:N | 19.9 | 15.49 | 11.99 | 11.96 | 12.01 |
| Total B (mg kg−1) | 62.39 | 43.56 | 37.22 | 28.19 | 15.71 |
| Total Ca (g kg−1) | 4.03 | 5.84 | 8.24 | 8.29 | 7.35 |
| Total Cl (g kg−1) | 6.86 | 4.22 | 0.89 | 0.98 | 0.56 |
| Total Cu (mg kg−1) | 2.7 | 3.6 | 12.3 | 13.6 | 27.8 |
| Total Fe (g kg−1) | 0.11 | 0.12 | 0.57 | 0.57 | 1.07 |
| Total K (g kg−1) | 1.82 | 1.96 | 1.78 | 2.23 | 1.38 |
| Total Mg(g kg−1) | 0.92 | 1.16 | 1.74 | 1.78 | 1.34 |
| Total Mn (mg kg−1) | 89.3 | 119.1 | 216.8 | 205.3 | 229.9 |
| Total Mo (mg kg−1) | 0.8 | 0.9 | 0.71 | 0.92 | 1.14 |
| Total P (g kg−1) | 0.61 | 0.76 | 1.23 | 1.39 | 1.42 |
| Total S (g kg−1) | 0.26 | 0.36 | 0.74 | 0.95 | 0.94 |
| Total Zn (mg kg−1) | 11.5 | 15.0 | 85.8 | 122.0 | 269.6 |
| Basal respiration (μg CO2 g−1 dw h−1) | 701.5 | 594.5 | 391.8 | 320.1 | 127.6 |
| Microbial biomass C (g kg−1) | 9.55 | 24.08 | 12.44 | 16.12 | 10.81 |
| Microbial biomass N (g kg−1) | 4.26 | 5.72 | 5.60 | 5.37 | 5.84 |
| Lignin (g kg−1) | 388.4 | 519.2 | 518.5 | 544.9 | 530.7 |
| Cellulose (g kg−1) | 217.1 | 219.9 | 209.1 | 192.1 | 188.1 |
| Hemicellulose (g kg−1) | 133.1 | 94.7 | 71.6 | 71.3 | 57.4 |

References
- Flora of Australia. Available online: https://profiles.ala.org.au/opus/foa/profile/Acacia dealbata (accessed on 29 July 2022).
- May, B.W.; Attiwill, P.M. Nitrogen-fixation by Acacia dealbata and changes in soil properties 5 years after mechanical disturbance or slash-burning following timber harvest. For. Ecol. Manag. 2003, 181, 339–355. [Google Scholar] [CrossRef]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 environmental weeds for classical biological control in Europe: A review of opportunities, regulations and other barriers to adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Gouws, A.J.; Shackleton, C.M. Abundance and correlates of the Acacia dealbata invasion in the northern Eastern Cape, South Africa. For. Ecol. Manag. 2019, 432, 455–466. [Google Scholar] [CrossRef]
- Vieites-Blanco, C.; González-Prieto, S.J. Invasiveness, ecological impact and control of acacias in southwestern Europe—A review. Web Ecol. 2020, 20, 33–51. [Google Scholar] [CrossRef]
- Lorenzo, P.; González, L.; Reigosa, M.J. The genus Acacia as invader: The characteristic case of Acacia dealbata Link in Europe. Ann. For. Sci. 2010, 67, 101. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, N.; Becerra, J.; Guedes, L.M.; Villaseñor-Parada, C.; González, L.; Hernández, V. Allelopathic effect of the invasive Acacia dealbata Link (Fabaceae) on two native plant species in south-central Chile. Gayana Bot. 2015, 72, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Acacia dealbata (acacia bernier). CABI Invasive Species Compendium. Species Datasheet. Available online: https://www.cabi.org/isc/datasheet/2207 (accessed on 19 September 2022).
- Lorenzo, P.; Pazos-Malvido, E.; Rubido-Bará, M.; Reigosa, M.J.; González, L. Invasion by the leguminous tree Acacia dealbata (Mimosaceae) reduces the native understorey plant species in different communities. Aust. J. Bot. 2012, 60, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Reigosa, M.J.; Carballeira, A. Phenological phase influences allelopathic potential of water leachates of Acacia dealbata obtained using simulated rain. J. Allelochem. Interact. 2016, 2, 9–15. [Google Scholar]
- Aguilera, N.; Becerra, J.; Villaseñor-Parada, C.; Lorenzo, P.; González, L.; Hernández, V. Effects and identification of chemical compounds released from the invasive Acacia dealbata Link. Chem. Ecol. 2015, 31, 479–493. [Google Scholar] [CrossRef]
- Lorenzo, P.; Rodríguez, J.; González, L.; Rodríguez-Echeverría, S. Changes in microhabitat, but not allelopathy, affect plant establishment after Acacia dealbata invasion. J. Plant Ecol. 2017, 10, 610–617. [Google Scholar]
- Acacia dealbata: Catálogo Español de Especies Exóticas Invasoras. Available online: https://www.miteco.gob.es/es/biodiversidad/temas/conservacion-de-especies/acacia_dealbata_2013_tcm30-69798.pdf (accessed on 20 September 2022).
- Spanish Government. Ley 42/2007, de 13 de diciembre, del Patrimonio Natural y de la Biodiversidad. Available online: https://www.boe.es/buscar/pdf/2007/BOE-A-2007-21490-consolidado.pdf (accessed on 20 September 2022).
- Moran, V.C.; Hoffmann, J.H. Conservation of the fynbos biome in the Cape Floral Region: The role of biological control in the management of invasive alien trees. BioControl 2012, 57, 139–149. [Google Scholar] [CrossRef]
- Flora Invasora de Galicia, Xunta de Galicia. Available online: https://cmatv.xunta.gal/seccion-feitos-vitais?content=Direccion_Xeral_Conservacion_Natureza/Biodiversidade/seccion.html&sub=Informacion_especies/&ui=Direccion_Xeral_Conservacion_Natureza/Dinamico/Especies/Invasoras/Flora/especie_0003.html (accessed on 25 August 2022).
- Nunes, L.J.R.; Raposo, M.A.M.; Meireles, C.I.R.; Pinto Gomes, C.J.; Almeida Ribeiro, N.M.C. Control of Invasive Forest Species through the Creation of a Value Chain: Acacia dealbata Biomass Recovery. Environments 2020, 7, 39. [Google Scholar] [CrossRef]
- López-Hortas, L.; Rodríguez-González, I.; Díaz-Reinoso, B.; Torres, M.D.; Moure, A.; Domínguez, H. Tools for a multiproduct biorefinery of Acacia dealbata biomass. Ind. Crop. Prod. 2021, 169, 113655. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Gómez-Brandón, M. Vermicomposting: Earthworms Enhance the Work of Microbes. In Microbes at Work: From Wastes to Resources; Insam, H., Franke-Whittle, I., Goberna, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 93–114. [Google Scholar]
- Reynolds, J.W. Earthworms of the world. Global. Divers. 1994, 4, 11–16. [Google Scholar]
- Domínguez, J. Earthworms and vermicomposting. In Earthworms—The Ecological Engineers of Soil; Ray, S., Ed.; Intech Open Science: Rijeka, Croatia, 2018; pp. 63–77. [Google Scholar]
- Domínguez, J. State of the art and new perspectives on vermicomposting research. In Earthworm Ecology, 2nd ed.; Edwards, C.A., Ed.; CRC Press LLC: Boca Raton, FL, USA, 2004; pp. 401–424. [Google Scholar]
- Domínguez, J.; Martínez-Cordeiro, H.; Álvarez-Casas, M.; Lores, M. Vermicomposting grape marc yields high quality organic biofertiliser and bioactive polyphenols. Waste Manag. Res. 2014, 32, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Hernandez, J.C.; Domínguez, J. Vermicompost derived from spent coffee grounds: Assessing the potential for enzymatic bioremediation. In Handbook of Coffee Processing By-Products Sustainable Applications; Galanakis, C.M., Ed.; Academic Press: London, UK, 2017; pp. 369–398. [Google Scholar]
- Domínguez, J.; Gómez-Brandón, M.; Martínez-Cordeiro, H.; Lores, M. Bioconversion of Scotch broom into a high-quality organic fertiliser: Vermicomposting as a sustainable option. Waste Manag. Res. 2018, 36, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Goering, H.K.; van Soest, P.J. Forage Fiber Analysis. Agricultural Handbook No. 379; Agricultural Research Service, United States Department of Agriculture: Washington, DC, USA, 1970. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method for measuring microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Klein, D.R. The introduction, increase, and crash of reindeer on St. Matthew Island. J. Wildl. Manag. 1968, 32, 350–367. [Google Scholar] [CrossRef]
- Ganesh, P.S.; Gajalakshmi, S.; Abbasi, S.A. Vermicomposting of the leaf litter of acacia (Acacia auriculiformis): Possible roles of reactor geometry, polyphenols, and lignin. Bioresour. Technol. 2009, 100, 1819–1827. [Google Scholar] [CrossRef]
- Zhou, B.; Chen, Y.; Zhang, C.; Li, J.; Tang, H.; Liu, J.; Dai, J.; Tang, J. Earthworm biomass and population structure are negatively associated with changes in organic residue nitrogen concentration during vermicomposting. Pedosphere 2021, 31, 433–439. [Google Scholar] [CrossRef]
- Fornes, F.; Mendoza-Hernández, D.; García-de-la-Fuente, R.; Abad, M.; Belda, R.M. Composting versus vermicomposting: A comparative study of organic matter evolution through straight and combined processes. Bioresour. Technol. 2012, 118, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Gong, X.; Sun, X.; Li, S.; Yu, X. Comparison of chemical and microbiological changes during the aerobic composting and vermicomposting of green waste. PLoS ONE 2018, 13, e0207494. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, J.; Aira, M.; Kolbe, A.R.; Gómez-Brandón, M.; Pérez-Losada, M. Changes in the composition and function of bacterial communities during vermicomposting may explain beneficial properties of vermicompost. Sci. Rep. 2019, 9, 9657. [Google Scholar] [CrossRef] [Green Version]
- Ndegwa, P.M.; Thompson, S.A.; Das, K.C. Effects of stocking density and feeding rate on vermicomposting of biosolids. Bioresour. Technol. 2000, 71, 5–12. [Google Scholar] [CrossRef]
- Brito, L.M.; Mourao, I.; Coutinho, J.; Smith, S. Composting for management and resource recovery of invasive Acacia species. Waste Manag. Res. 2013, 31, 1125–1132. [Google Scholar] [CrossRef]
- European Union. Consolidated Text: Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and(EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003 (Text with EEA Relevance) Text with EEA Relevance. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A02019R1009-20220716 (accessed on 29 August 2022).
- Mañas, P.; de Heras, J.L. Phytotoxicity test applied to sewage sludge using Lactuca sativa L. and Lepidium sativum L. seeds. Int. J. Environ. Sci. Technol. 2018, 15, 273–280. [Google Scholar] [CrossRef]
- Paré, T.; Gregorich, E.G.; Dinel, H. Effects of stockpiled and composted manures on germination and initial growth of cress (Lepidium sativum). Biol. Agric. Hortic. 1997, 14, 1–11. [Google Scholar] [CrossRef]
- Contreras-Ramos, S.M.; Escamilla-Silva, E.M.; Dendooven, L. Vermicomposting of biosolids with cow manure and oat straw. Biol. Fertil. Soils 2005, 41, 190–198. [Google Scholar] [CrossRef]
- Majlessi, M.; Eslami, A.; Saleh, H.N.; Mirshafieean, S.; Babaii, S. Vermicomposting of food waste: Assessing the stability and maturity. J. Environ. Health Sci. Eng. 2012, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Paradelo, R.; Moldes, A.B.; Rodríguez, M.; Barral, M.T. Relationship between heavy metals and phytotoxicity in composts. Cienc. Tecnol. Aliment. 2008, 6, 143–151. [Google Scholar] [CrossRef]



| PC1 | PC2 | PC3 | PC4 | |
|---|---|---|---|---|
| Explained variation (%) | 61.5 | 15.3 | 8.1 | 5.4 |
| Eigenvalue | 12.9 | 3.2 | 1.7 | 1.1 |
| Variable loadings | ||||
| pH | 0.47 | 0.64 | −0.45 | 0.03 |
| Electrical Conductivity | −0.89 | −0.25 | 0.31 | −0.00 |
| Moisture content | 0.90 | 0.23 | −0.14 | −0.14 |
| Organic Matter content | −0.87 | 0.36 | −0.00 | −0.00 |
| N total concentration | 0.89 | 0.31 | −0.18 | −0.20 |
| C total concentration | −0.78 | 0.54 | −0.04 | 0.14 |
| B total concentration | −0.83 | 0.14 | 0.28 | −0.13 |
| Ca total concentration | 0.83 | 0.45 | 0.25 | −0.07 |
| Cl total concentration | −0.95 | −0.13 | 0.16 | 0.12 |
| Cu total concentration | 0.90 | −0.37 | 0.06 | 0.15 |
| Fe total concentration | 0.93 | −0.34 | 0.07 | 0.05 |
| K total concentration | −0.07 | 0.73 | 0.60 | 0.15 |
| Mg total concentration | 0.71 | 0.55 | 0.35 | −0.22 |
| Mn total concentration | 0.95 | 0.12 | 0.15 | −0.10 |
| Mo total concentration | 0.42 | −0.20 | 0.34 | 0.74 |
| P total concentration | 0.94 | 0.13 | 0.29 | −0.00 |
| S total concentration | 0.95 | 0.08 | 0.25 | −0.02 |
| Zn total concentration | 0.86 | −0.39 | 0.05 | 0.23 |
| Basal Respiration | −0.92 | 0.24 | 0.12 | −0.02 |
| Microbial biomass C | −0.07 | 0.73 | −0.23 | 0.51 |
| Microbial biomass N | 0.24 | 0.16 | −0.58 | 0.24 |
| F(4,20) | p-Val | Day 0 | Day 14 | Day 28 | Day 42 | Day 56 | |
|---|---|---|---|---|---|---|---|
| Earthworm density (individuals m−2) | 254.9 | 7.5 × 10−17 | 345 d ± 23 | 434 c ± 40 | 877 a ± 34 | 889 a ± 27 | 665 b ± 46 |
| Earthworm biomass (g m−2) | 493.3 | 1.13 × 10−19 | 99.4 e ± 10 | 134.2 d ± 15.6 | 371.9 b ± 18.9 | 404.2 a ± 15.2 | 208.2 c ± 5.8 |
| F(4,10) | p-Val | C | AD5 | AD10 | VC5 | VC10 | |
|---|---|---|---|---|---|---|---|
| Germination percentage (%) | 4.63 | 0.023 | 96.7 a ± 2.9 | 81.7 b ± 5.8 | 78.3 b ± 5.8 | 88.3 a,b ± 7.6 | 93.3 a,b ± 7.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintela-Sabarís, C.; Mendes, L.A.; Domínguez, J. Vermicomposting as a Sustainable Option for Managing Biomass of the Invasive Tree Acacia dealbata Link. Sustainability 2022, 14, 13828. https://doi.org/10.3390/su142113828
Quintela-Sabarís C, Mendes LA, Domínguez J. Vermicomposting as a Sustainable Option for Managing Biomass of the Invasive Tree Acacia dealbata Link. Sustainability. 2022; 14(21):13828. https://doi.org/10.3390/su142113828
Chicago/Turabian StyleQuintela-Sabarís, Celestino, Luís A. Mendes, and Jorge Domínguez. 2022. "Vermicomposting as a Sustainable Option for Managing Biomass of the Invasive Tree Acacia dealbata Link" Sustainability 14, no. 21: 13828. https://doi.org/10.3390/su142113828