
Changes in the Vegetation, Soil Seed Bank and Soil Properties at Bait Sites in a Protected Area of the Central European Lower Montane Zone
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
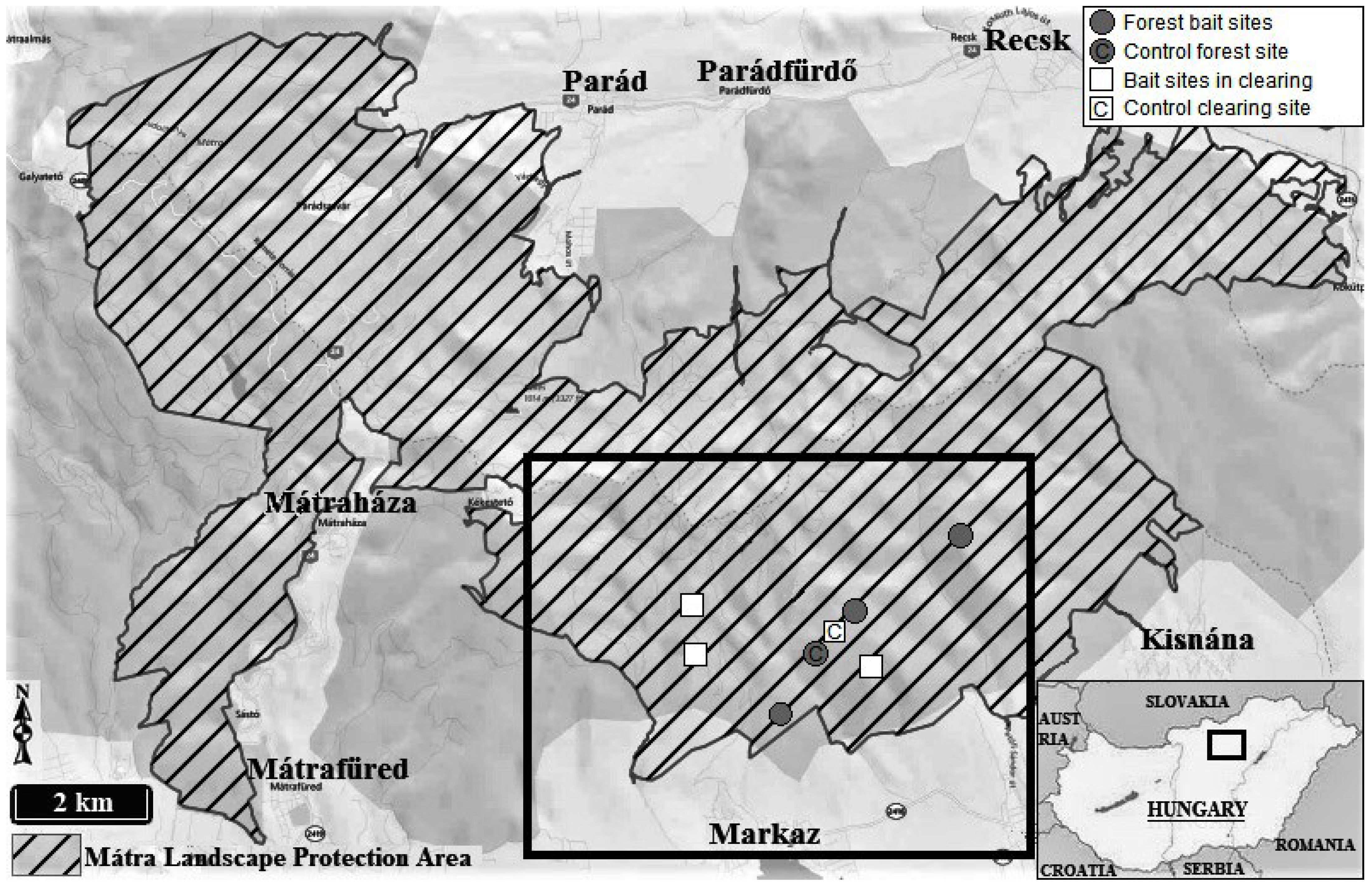
2.1. Study Area
2.2. Sampling Setup
2.3. Collection of Soil Seed Bank and Vegetation Data
2.4. Soil Sampling
2.5. Data Processing
3. Results
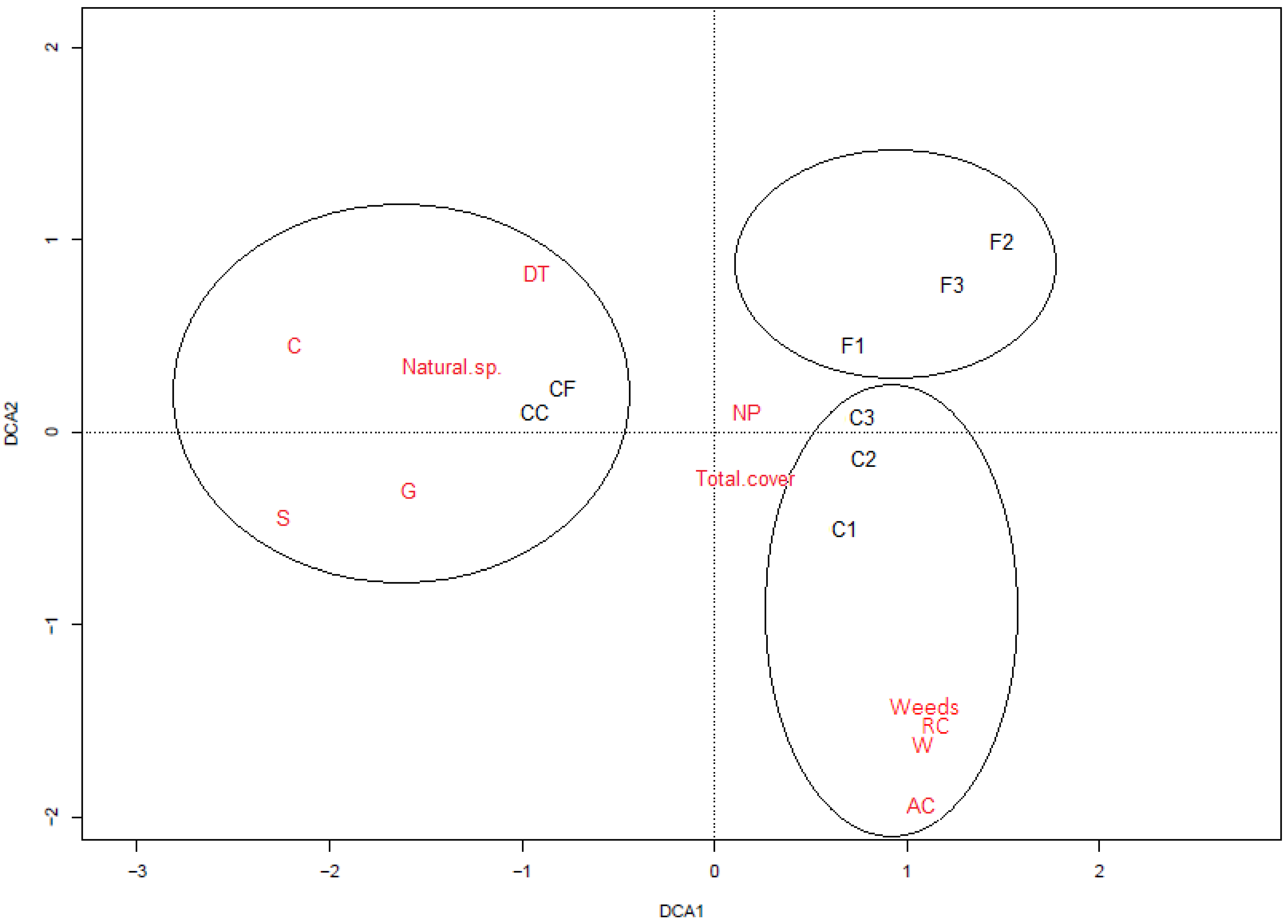
3.1. Vegetation Composition
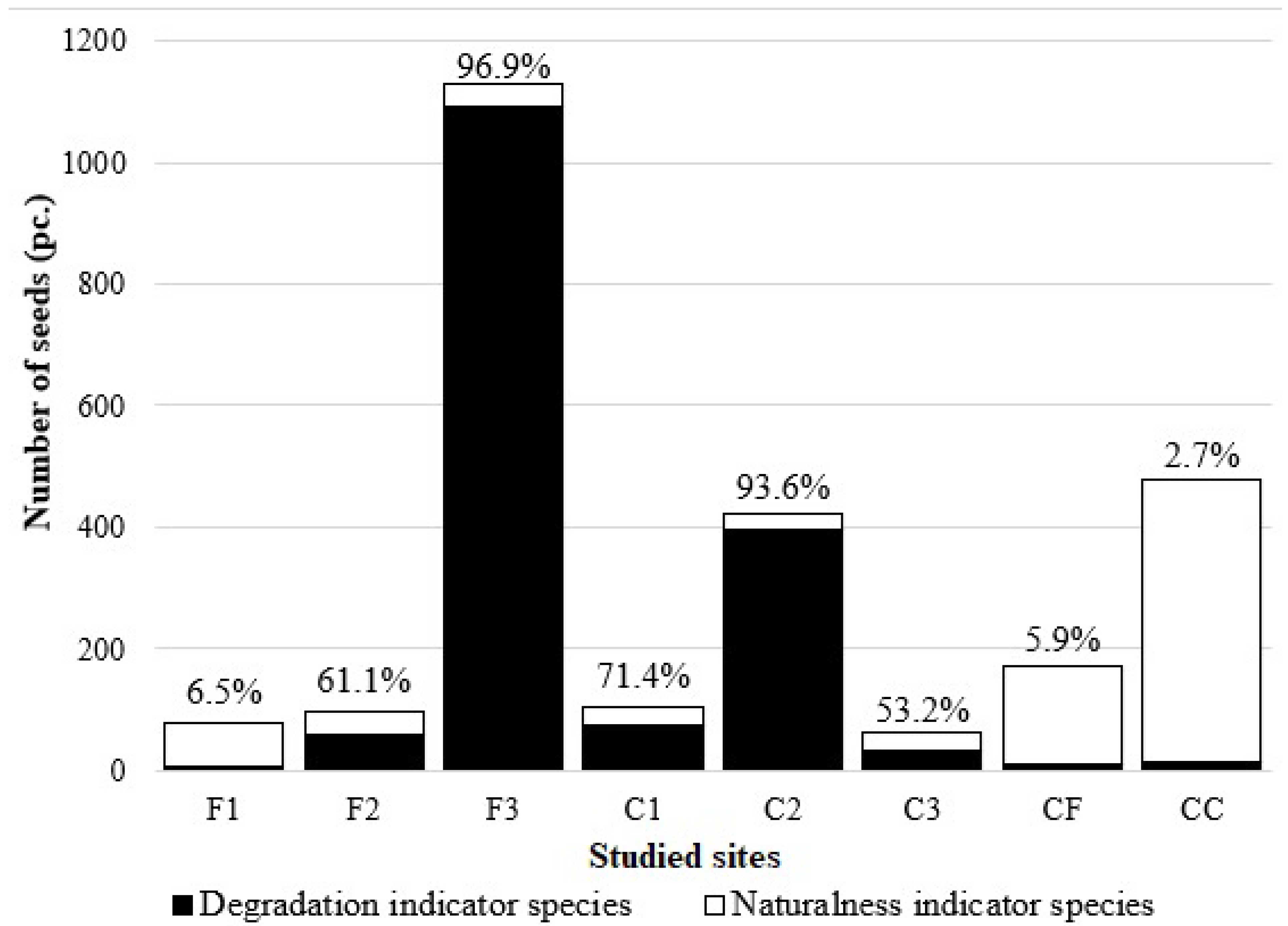
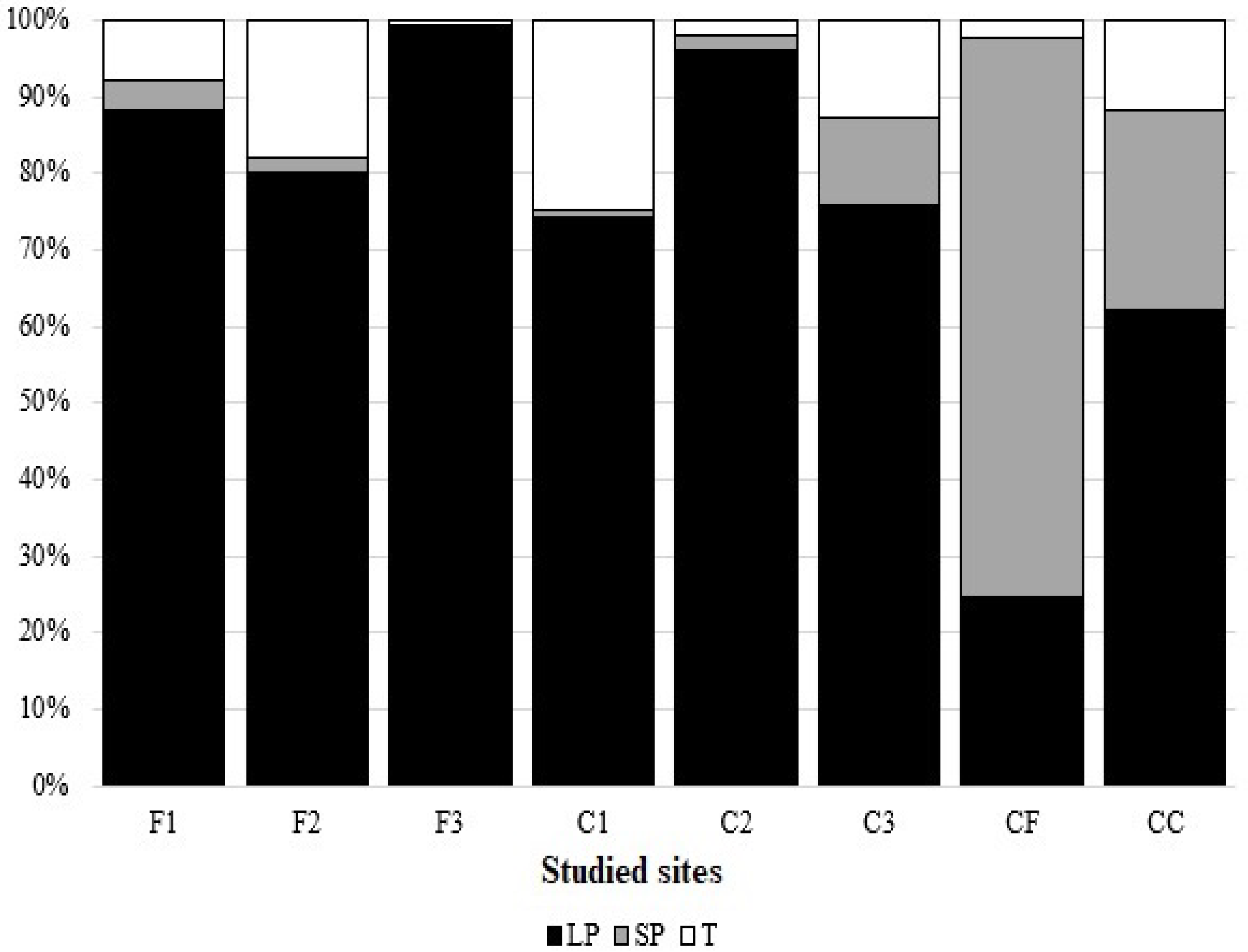
3.2. Seed Bank Composition and Density
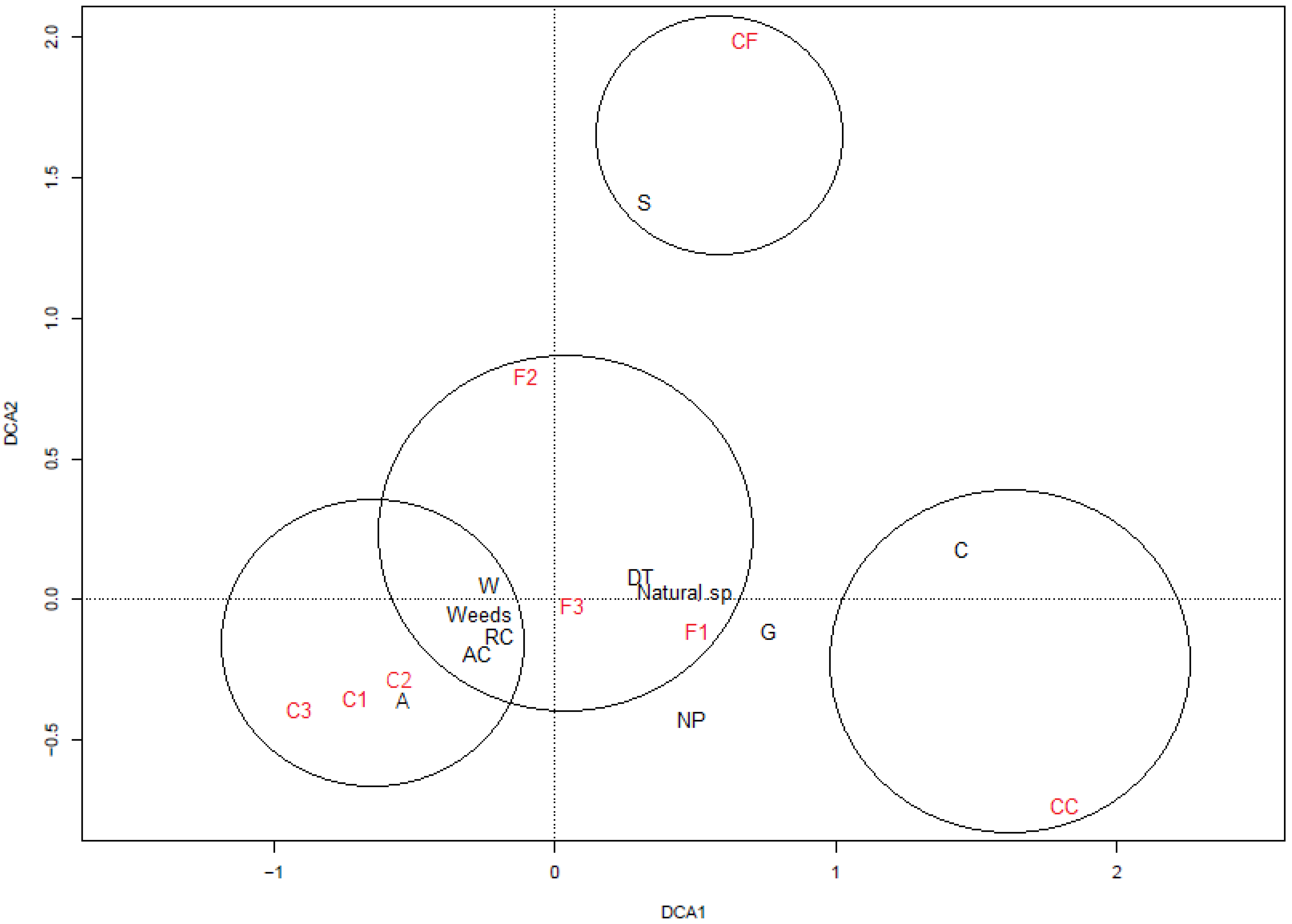
3.3. Relationship between Vegetation and Seed Bank
3.4. Seed Bank Persistence
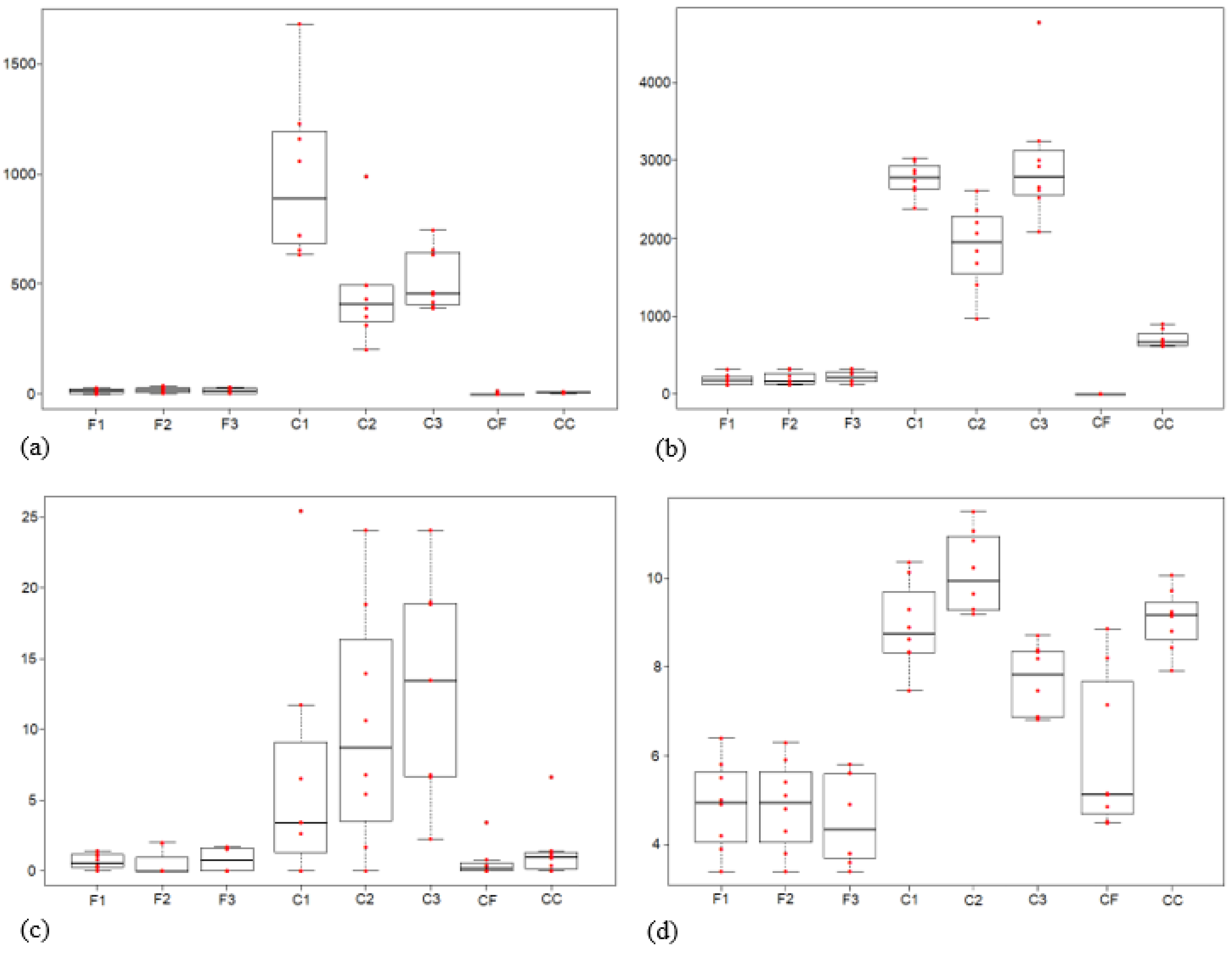
3.5. Soil Characteristics
4. Discussion
4.1. Vegetation Composition
4.2. Seed Bank Composition and Density
4.3. Relationship between Vegetation and Seed Bank
4.4. Seed Bank Persistence
4.5. Soil Characteristics
5. Conclusions and Management Implications
Novelty and Innovation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, J.M.; Gerhardt, P.; Steyaert, S.M.J.G.; Hochbichler, E.; Hacklander, K. Diversionary feeding can reduce red deer habitat selection pressure on vulnerable forest stands, but is not a panacea for red deer damage. For. Ecol. Manag. 2018, 407, 166–173. [Google Scholar] [CrossRef]
- Rinella, M.J.; Dean, R.; Vavra, M.; Parks, C.G. Vegetation responses to supplemental winter feeding of elk in western Wyoming. W. N. Am. Nat. 2012, 72, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Selva, N.; Berezowska-Cnota, T.; Elguero-Claramunt, I. Unforeseen Effects of Supplementary Feeding: Ungulate Baiting Sites as Hotspots for Ground-Nest Predation. PLoS ONE 2014, 9, e90740. [Google Scholar] [CrossRef]
- Heltai, M.; Sonkoly, K. The role and opportunities of feeding in game management (Review). Anim. Welf. Ethol. Hous. Syst. 2009, 5, 1–22. [Google Scholar]
- Gervilla, C.; Rita, J.; Cursach, J. Contaminant seeds in imported crop seed lots: A non-negligible human-mediated pathway for introduction of plant species to islands. Weed Res. 2019, 59, 245–253. [Google Scholar] [CrossRef]
- Rusvai, K.; Saláta, D.; Falvai, D.; Czóbel, S. Assesment of weed invasion at bait sites in a Central European lower montane zone. Perspect. Plant Ecol. Evol. Syst. 2022, 55, 125667. [Google Scholar] [CrossRef]
- Gioria, M.; Pyšek, P. The legacy of plant invasions: Changes in the soil seed bank of invaded plant communities. Bioscience 2015, 66, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Galloway, A.D.; Holmes, P.M.; Gaertner, M.; Esler, K.J. The impact of pine plantations on fynbos above-ground vegetation and soil seed bank composition. S. Afr. J. Bot. 2017, 113, 300–307. [Google Scholar] [CrossRef]
- Gioria, M.; Osborne, B.A. Assessing the impact of plant invasions on soil seed bank communities: Use of univariate and multivariate statistical approaches. J. Veg. Sci. 2009, 20, 547–556. [Google Scholar] [CrossRef]
- Gooden, B.; French, K. Impacts of alien grass invasion in coastal seed banks vary amongst native growth forms and dispersal strategies. Biol. Conserv. 2014, 171, 114–126. [Google Scholar] [CrossRef] [Green Version]
- Gioria, M.; Osborne, B.A. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci. 2014, 5, 501. [Google Scholar] [CrossRef] [PubMed]
- Venable, D.L.; Brown, J.S. The selective interactions of dispersal, dormancy, and seed size as adaptations for reducing risk in variable environments. Am. Nat. 1988, 131, 360–384. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Dairel, M.; Fidelis, A. The presence of invasive grasses affects the soil seed bank composition and dynamics of both invaded and non-invaded areas of open savannas. J. Environ. Manag. 2020, 276, 111291. [Google Scholar] [CrossRef]
- Simberloff, D.; von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Cadenasso, M.L.; Pickett, S.T.A. Effect of edge structure on the flux of species into forest interiors. Conserv. Biol. 2001, 15, 91–97. [Google Scholar] [CrossRef]
- Turner, P.J.; Scott, J.K.; Spafford, H. The ecological barriers to the recovery of bridal creeper ((Asparagus asparagoides L.) Druce) infested sites: Impacts on vegetation and the potential increase in other exotic species. Austral. Ecol. 2008, 33, 713–722. [Google Scholar] [CrossRef]
- Zhang, C.; Willis, C.G.; Ma, Z.; Ma, M.; Csontos, P.; Baskin, C.C.; Baskin, J.; Li, J.; Zhou, H.; Zhao, X.; et al. Direct and indirect effects of long-term fertilization on the stability of the persistent seed bank. Plant Soil 2019, 438, 239–250. [Google Scholar] [CrossRef]
- Lehoczky, E.; Reisinger, P.; Kőmíves, T. Loss of nutrients caused by excessive weediness at the early stage of maize vegetation period. Commun. Plant Soil Anal. 2005, 36, 415–422. [Google Scholar] [CrossRef]
- Bölöni, J.; Molnár, Z.; Biró, M.; Horváth, F. Distribution of the (semi-) natural habitats in Hungary II. Woodlands and shrublands. Acta Bot. Hung. 2008, 50, 107–148. [Google Scholar] [CrossRef]
- Katona, K.; Szemethy, L.; Csányi, S. Forest management practices and forest sensitivity to game damage in Hungary. Hung. Agric. Res. 2011, 1, 12–16. [Google Scholar]
- Manninger, M.; Edelényi, M.; Pödör, Z.; Jereb, L. The effect of temperature and precipitation on growth of beech (Fagus sylvatica L.) in Mátra Mountains, Hungary. In Applied Forestry Research in the 21st Century; Book of Abstracts; Forestry and Game Management Research Institute: Jíloviště, Czech Republic, 2011; p. 22. [Google Scholar]
- Juhász, O.; Fürjes-Mikó, Á.; Tenyér, A.; Somogyi, A.Á.; Aguilon, D.J.; Kiss, P.J.; Bátori, Z.; Maák, I. Consequences of Climate Change-Induced Habitat Conversions on Red Wood Ants in a Central European Mountain: A Case Study. Animals 2020, 10, 1677. [Google Scholar] [CrossRef] [PubMed]
- Bereczki, K.; Ódor, P.; Csóka, G.; Mag, Z.; Báldi, A. Effects of forest heterogeneity on the efficiency of caterpillar control service provided by birds in temperate oak forests. For Ecol. Manag. 2014, 327, 96–105. [Google Scholar] [CrossRef]
- Dövényi, Z. (Ed.) Mátra-Vidék. In Magyarország Kistájainak Katasztere; MTA Földrajztudományi Kutatóintézet: Budapest, Hungary, 2010; pp. 713–736. [Google Scholar]
- Standovár, T. Comparative Study of Vegetation and Soil Pattern in a Mountain Meadow (Mátra, Hungary) II. Soil Pattern and Its Overlap with Vegetation Pattern. Abstr. Bot. 1986, 10, 291–315. Available online: http://www.jstor.org/stable/43519136 (accessed on 8 October 2022).
- Milner, J.M.; Bonenfant, C.; Mysterud, A.; Gaillard, J.M.; Csányi, S.; Stenseth, N. Temporal and spatial development of red deer harvesting in Europe: Biological and cultural factors. J. Appl. Ecol. 2006, 43, 721–734. [Google Scholar] [CrossRef]
- Csányi, S.; Lehoczki, R. Ungulates and their management in Hungary. In European Ungulates and Their Management in the 21st Century; Apollonio, M., Andersen, R., Putman, R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 291–318. [Google Scholar]
- Durbecq, A.; Jaunatre, R.; Buisson, E.; Cluchier, A.; Bischoff, A. Identifying reference communities in ecological restoration: The use of environmental conditions driving vegetation composition. Restor. Ecol. 2020, 28, 1445–1453. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Harris, J.A. Restoration ecology: Repairing the earth’s ecosystems in the new millennium. Restor. Ecol. 2001, 9, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Matus, G.; Papp, M.; Tóthmérész, B. Impact of management on vegetation dynamics and seed bank formation of inland dune grassland in Hungary. Flora 2005, 200, 296–306. [Google Scholar] [CrossRef]
- Jacquemyn, H.; van Mechelen, C.; Brys, R.; Honnay, O. Management effects on the vegetation and soil seed bank of calcareous grasslands: An 11-year experiment. Biol. Conserv. 2011, 144, 416–422. [Google Scholar] [CrossRef]
- Török, P.; Kelemen, A.; Valkó, O.; Miglécz, T.; Tóth, K.; Tóth, E.; Sonkoly, J.; Kiss, R.; Csecserits, A.; Rédei, T.; et al. Succession in soil seed banks and its implications for restoration of calcareous sand grasslands. Restor. Ecol. 2017, 26, 134–140. [Google Scholar] [CrossRef]
- Buzás, I. Talaj-és agrokémiai vizsgálati módszerkönyv 2. A talajok fizikai-kémiai és kémiai vizsgálati módszerei. Agrokem. Talajtan 1989, 38, 504–505. (In Hungarian) [Google Scholar]
- Egner, J.; Riehm, H.; Domingo, W. Untersuchungen über die che-mische Bodenanalyse als Grundlage für die Beurteilung des Nährst-offzustandes der Böden II. Chemische Extraktionsmethoden zur Phosphor- und Kaliumbestimmung. K. Lantbr. Ann. 1960, 26, 199–215. (In German) [Google Scholar]
- MSZ-08-0210-1977; Methods for the Determination of the Organic Carbon Content of the Soil. Department of Agriculture and Tourism: Budapest, Hungary, 2021.
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian Flora. Acta Bot. Hung. 1995, 39, 97–181. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1991, 18, 1–248. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley: Chichester, NY, USA, 1979. [Google Scholar]
- Thompson, K.; Bakker, J.P.; Bekker, R.M. The Soil Seed Banks of North West Europe: Methodology, Density and Longevity; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Rejmánek, M.; Richardson, D.M.; Pyšek, P. Plant Invasions and Invasibility of Plant Communities. In Vegetation Ecology, 2nd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 387–424. [Google Scholar] [CrossRef]
- Martin, P.H.; Canham, C.D.; Marks, P.L. Why forests appear resistant to exotic plant invasions: Intentional introductions, stand dynamics, and the role of shade tolerance. Front. Ecol. Environ. 2009, 7, 142–149. [Google Scholar] [CrossRef]
- Bourgeois, B.; Munoz, F.; Fried, G.; Mahaut, L.; Armengot, L.; Denelle, P.; Storkey, J.; Gaba, S.; Violle, C. What makes a weed a weed? A large-scale evaluation of arable weeds through a functional lens. Am. J. Bot. 2019, 106, 90–100. [Google Scholar] [CrossRef]
- Tóth, K.; Lukács, B.A.; Radócz, S.; Simon, E.A. Magbank szerepe a szikes gyepek diverzitásának fenntartásában a Hortobágyi Nemzeti Park területén. Bot. Közlemények 2015, 102, 141–157. (In Hungarian) [Google Scholar] [CrossRef]
- Kiss, R.; Valkó, O.; Tóthmérész, B.; Török, P. Seed bank research in Central-European grasslands—An overview. In Seed Banks: Types, Roles and Research; Chapter 1; Murphy, J., Ed.; Nova Science: Hauppauge, NY, USA, 2016; pp. 1–34. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds, 2nd ed; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Heinken, T.; Schmidt, M.; von Oheimb, G.; Kriebitzsch, W.-U.; Ellenberg, H. Soil seed banks near rubbing trees indicate dispersal of plant species into forests by wild boar. Basic Appl. Ecol. 2006, 7, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.; Johansson, M.; Hughes, F.; Hayes, A.; Richards, K.; Nilsson, C. Interactive effects of soil moisture, vegetation canopy, plant litter and seed addition on plant diversity in a wetland community. J. Ecol. 2003, 91, 976–986. [Google Scholar] [CrossRef]
- Bossuyt, B.; Honnay, O. Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J. Veg. Sci. 2008, 19, 875–884. [Google Scholar] [CrossRef]
- Miaojun, M.; Xianhui, Z.; Guozhen, D. Role of soil seed bank along a disturbance gradient in an alpine meadow on the Tibet plateau. Flora 2010, 205, 128–134. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; McDonell, M.J. Seed bank dynamics intemperate deciduous forest. In Ecology of Soil Seed Banks; Leck, M.A., Parker, V.T., Simpson, R.L., Eds.; Academic Press: London, UK, 1989; pp. 123–147. [Google Scholar]
- Csontos, P. A természetes magbank, valamint a hazai flóramagökológiai vizsgálatának új eredményei. Some new results improving the knowledge of the natural soil seed banks of the Hungarian flora. Kanitzia 2010, 17, 77–110. (In Hungarian) [Google Scholar]
- Valkó, O.; Török, P.; Tóthmérész, B.; Matus, G. Restoration potential in seed banks of acidic fen and dry-mesophilous meadows: Can restoration be based on local seed banks? Restor. Ecol. 2011, 19, 9–15. [Google Scholar] [CrossRef]
- Lacko-Bartosova, M.; Minar, M.; Vranovska, Z.; Strasser, D. Weed seed bank in ecological and integrated farming system. Rostl. Vyrob. 2000, 46, 319–324. [Google Scholar]
- Magyar, I.E. Gyógynövényes fűmagkeverék gyomosodási vizsgálata a telepedési idő és a talaj magbank hatására. Magy. Gyomkutatás Technol. 2005, 6, 37–51. (In Hungarian) [Google Scholar]
- Simić, M.; Spasojević, I.; Brankov, M.; Dragicevic, V. Weeds Seed Bank Richness in Maize Field: Effects of Crop Rotation and Herbicides. In Proceedings of the 5th International Scientific Agricultural Symposium Agrosym 2014, Jahorina, Bosnia and Herzegovina, 25–28 September 2014; pp. 501–507. Available online: https://www.cabdirect.org/cabdirect/abstract/20153437535 (accessed on 8 October 2022).
- Koncz, G.; Papp, M.; Török, P.; Kotroczó, Z.; Krakomperger, Z.; Matus, G.; Tóthmérész, B. The role of seed bank in the dynamics of understorey in an oak forest in Hungary. Acta Biol. Hung. 2010, 61, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Bossuyt, B.; Hermy, M. Influence of land use history on seed banks in European temperate forest ecosystems: A review. Ecography 2001, 24, 225–238. [Google Scholar] [CrossRef]
- Halassy, M. Possible role of the seed bank in the restoration of open sand grasslands in old fields. Commun. Ecol. 2001, 2, 101–108. [Google Scholar] [CrossRef]
- Tóth, K.; Hüse, B. Soil seed banks in loess grasslands and their role in grassland recovery. Appl. Ecol. Environ. Res. 2014, 12, 537–547. [Google Scholar] [CrossRef]
- Handlová, V.; Münzbergová, Z. Seed banks of managed and degraded grasslands in the Krkonoše Mts., Czech Republic. Folia Geobot. 2006, 41, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Kemény, G.; Nagy, Z.; Tuba, Z. Seed bank dynamics in a semiarid sandy grassland in Hungary. Ekol. Bratisl. 2005, 24, 1–13. [Google Scholar]
- Thompson, K.; Hodkinson, D.J. Seed mass, habitat and life history: A re-analysis of Salisbury. New Phytol. 1998, 138, 163–166. [Google Scholar] [CrossRef]
- Tierney, G.L.; Fahey, T.J. Soil seed bank dynamics of pin cherry in a northern hardwood forest, New Hampshire, USA. Can. J. For. Res. 1998, 28, 1471–1480. [Google Scholar] [CrossRef]
- Thompson, K. The functional ecology of seed banks. In Seed Ecology; Fenner, M., Ed.; Chapman & Hall: London, UK, 1985; pp. 231–258. [Google Scholar]
- Ádám, R.; Ódor, P.; Bidló, A.; Somay, L.; Bölöni, J. The effect of light, soil pH and stand heterogeneity on understory species composition of dry oak forests in the North Hungarian Mountains. Community Ecol. 2018, 19, 259–271. [Google Scholar] [CrossRef]
- Lengyel, A.; Purger, D.; Csiky, J. Classification of mesic grasslands and their transitions of South Transdanubia (Hungary). Acta Bot. Croat. 2012, 71, 31–50. [Google Scholar] [CrossRef]
- Turunen, M.; Oksanen, P.; Vuojala-Magga, T.; Markkula, I.; Sutinen, M.-L.; Hyvönen, J. Impacts of winter feeding of reindeer on vegetation and soil in the sub-Arctic: Insights from a feeding experiment. Polar Res. 2013, 32, 18610. [Google Scholar] [CrossRef]
- Skuodiené, R.; Repšiené, R.; Karcauskiene, D.; Siaudinis, G. Assessment of the weed incidence and weed seed bank of crops under different pedological traits. Appl. Ecol. Environ. Res. 2018, 16, 1131–1142. [Google Scholar] [CrossRef]
- Weidenhamer, J.D.; Callaway, R.M. Direct and Indirect Effects of Invasive Plants on Soil Chemistry and Ecosystem Function. J. Chem. Ecol. 2010, 36, 59–69. [Google Scholar] [CrossRef]
- Barrios-Garcia, M.N.; Ballari, S.A. Impact of wild boar (Sus scrofa) in its introduced and native range: A review. Biol. Invasions 2012, 14, 2283–2300. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Aparicio, L.G.; Haider, S.; Heger, T.; Lortie, C.J.; Pyšek, P.; Strayer, D.L. Support for major hypotheses in invasion biology is uneven and declining. NeoBiota 2012, 14, 1–20. [Google Scholar] [CrossRef]
- Ruprecht, E.; Szabó, A. Grass litter is a natural seed trap in long-term undisturbed grassland. J. Veg. Sci. 2012, 23, 495–504. [Google Scholar] [CrossRef]
- Lewis, J. Longevity of crop and weed seeds: Survival after 20 years in soil. Weed Res. 1973, 13, 179–191. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Point | Type | GPS Coordinates |
|---|---|---|
| F1 | forest bait site | 47°50′32.8″ N 20°03′54.7″ E |
| F2 | forest bait site | 47°51′57.6″ N 20°06′03.5″ E |
| F3 | forest bait site | 47°51′20.9″ N 20°04′49.4″ E |
| C1 | bait site in clearing | 47°51′24.7″ N 20°02′50.8″ E |
| C2 | bait site in clearing | 47°51′00.3″ N 20°02′52.6″ E |
| C3 | bait site in clearing | 47°50′55.0″ N 20°04′58.9″ E |
| CF | control forest site | 47°51′14.4″ N 20°04′49.8″ E |
| CC | control clearing site | 47°51′16.3″ N 20°04′50.3″ E |
| F1 | F2 | F3 | C1 | C2 | C3 | CF | CC | Forest vs. Clearing | Baits vs. Controls | |
|---|---|---|---|---|---|---|---|---|---|---|
| Vegetation cover (%) | 5 | 2 | 5 | 95 | 50 | 30 | 36.5 | 100.0 | * | ns. |
| Total cover (%) | 15 | 3 | 6 | 184 | 54 | 43 | 140.6 | 271.0 | ns. | * |
| Cover of natural species | 9 | 0 | 2 | 28 | 11 | 14 | 138.1 | 269.1 | ns. | *** |
| Cover of weeds | 6 | 3 | 4 | 156 | 43 | 29 | 2.5 | 2 | ns. | ns. |
| Weed% | 40.0 | 100.0 | 66.7 | 84.8 | 79.6 | 67.4 | 1.8 | 0.7 | ns. | ** |
| Total sp. no. (pc.) | 11 | 3 | 3 | 34 | 13 | 14 | 32 | 44 | ns. | * |
| Number of natural species | 6 | 0 | 1 | 12 | 2 | 6 | 31 | 42 | ns. | *** |
| Number of weeds | 5 | 3 | 2 | 22 | 11 | 8 | 1 | 2 | ns. | ns. |
| Weed% | 45.5 | 100.0 | 66.7 | 64.7 | 84.6 | 57.1 | 3.1 | 4.5 | ns. | ** |
| F1 | F2 | F3 | C1 | C2 | C3 | CF | CC | Forest vs. Clearing | Baits vs. Controls | |
|---|---|---|---|---|---|---|---|---|---|---|
| Bare soil surface (%) | 85 | 93 | 80 | 5 | 50 | 40 | 59.35 | 0,00 | * | ns. |
| Stones (%) | 10 | 5 | 15 | 0 | 0 | 30 | 4.15 | 0.00 | ns. | ns. |
| Soil pH (H2O) | 5.3 | 5.7 | 5.3 | 6.5 | 6.0 | 6.8 | 5.6 | 5.7 | * | ns. |
| Soil pH (KCl) | 4.5 | 4.8 | 4.5 | 5.5 | 5.2 | 5.8 | 4.7 | 4.9 | * | ns. |
| Salinity (%) | 0.09 | 0.07 | 0.07 | 0.08 | 0.07 | 0.09 | 0.05 | 0.06 | ns. | * |
| SOC (%) | 4.9 | 4.9 | 4.6 | 8.9 | 10.1 | 7.7 | 6.0 | 9.1 | ** | ns. |
| Soil humidity | 47.1 | 49.0 | 49.0 | 44.9 | 42.3 | 50.3 | 39.4 | 48.1 | ns. | ns. |
| Phosphate (µg/g) | 14.7 | 17.8 | 15.9 | 980.5 | 457.0 | 517.9 | 1.9 | 7.6 | * | ns. |
| Potassium (µg/mL) | 183.0 | 196.7 | 220.1 | 2763.2 | 1888.1 | 2976.0 | 0.0 | 702.7 | ** | ns. |
| NO3-N (mg kg−1) | 0.7 | 0.5 | 0.8 | 6.6 | 10.2 | 13.1 | 0.6 | 1.4 | ** | ns. |
| NH4-N (mg kg−1) | 16.4 | 18.6 | 19.3 | 16.5 | 18.9 | 18.7 | 16.2 | 18.6 | ns. | ns. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusvai, K.; Wichmann, B.; Saláta, D.; Grónás, V.; Skutai, J.; Czóbel, S. Changes in the Vegetation, Soil Seed Bank and Soil Properties at Bait Sites in a Protected Area of the Central European Lower Montane Zone. Sustainability 2022, 14, 13134. https://doi.org/10.3390/su142013134
Rusvai K, Wichmann B, Saláta D, Grónás V, Skutai J, Czóbel S. Changes in the Vegetation, Soil Seed Bank and Soil Properties at Bait Sites in a Protected Area of the Central European Lower Montane Zone. Sustainability. 2022; 14(20):13134. https://doi.org/10.3390/su142013134
Chicago/Turabian StyleRusvai, Katalin, Barnabás Wichmann, Dénes Saláta, Viktor Grónás, Julianna Skutai, and Szilárd Czóbel. 2022. "Changes in the Vegetation, Soil Seed Bank and Soil Properties at Bait Sites in a Protected Area of the Central European Lower Montane Zone" Sustainability 14, no. 20: 13134. https://doi.org/10.3390/su142013134