Regression Models to Estimate Accumulation Capability of Six Metals by Two Macrophytes, Typha domingensis and Typha elephantina, Grown in an Arid Climate in the Mountainous Region of Taif, Saudi Arabia

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Design
2.3. Sediment Sampling and Analysis
2.4. Quality Assurance and Control
2.5. Data Analysis
3. Results
3.1. Sediment Analysis
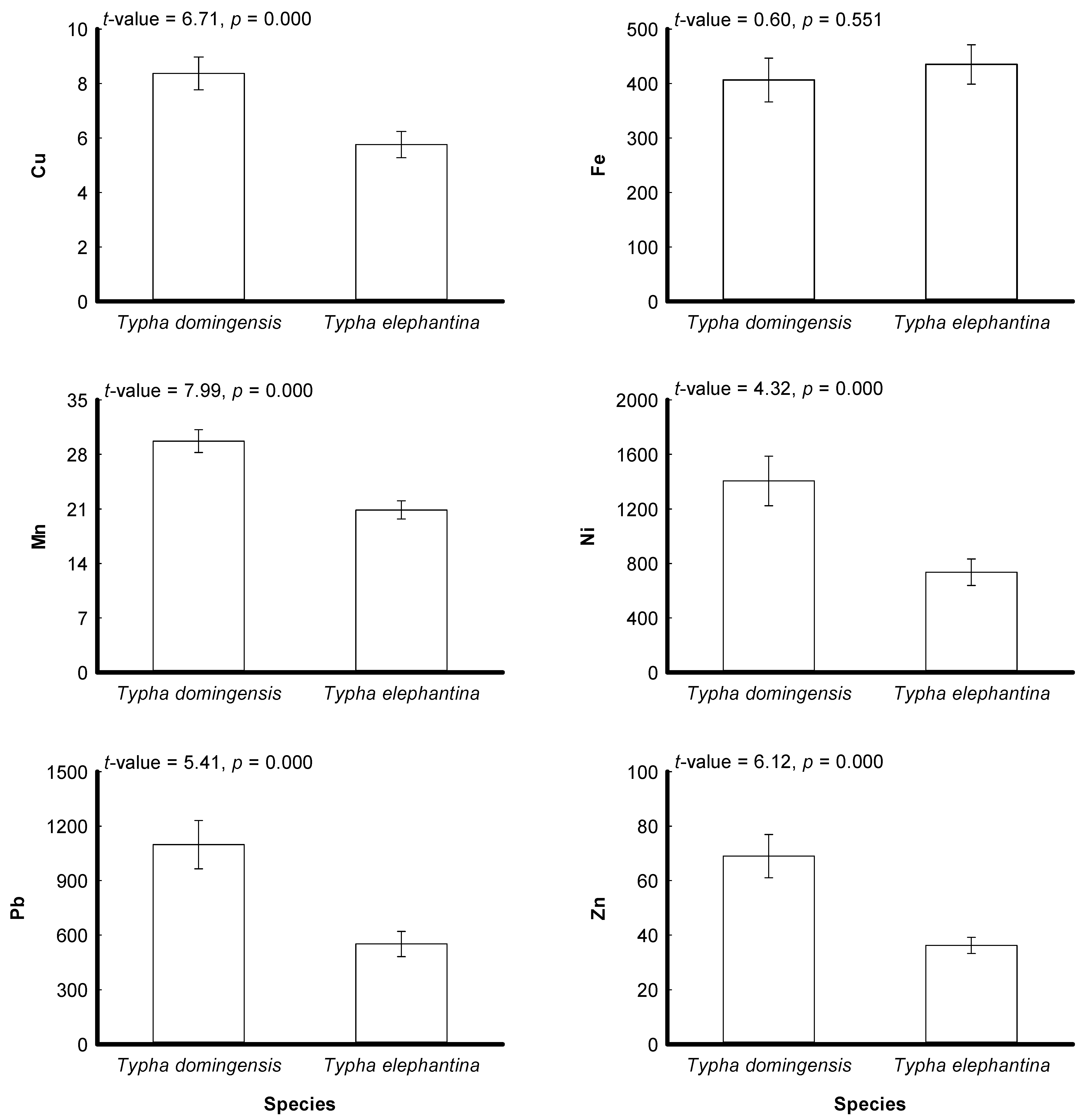
3.2. Plant Metals
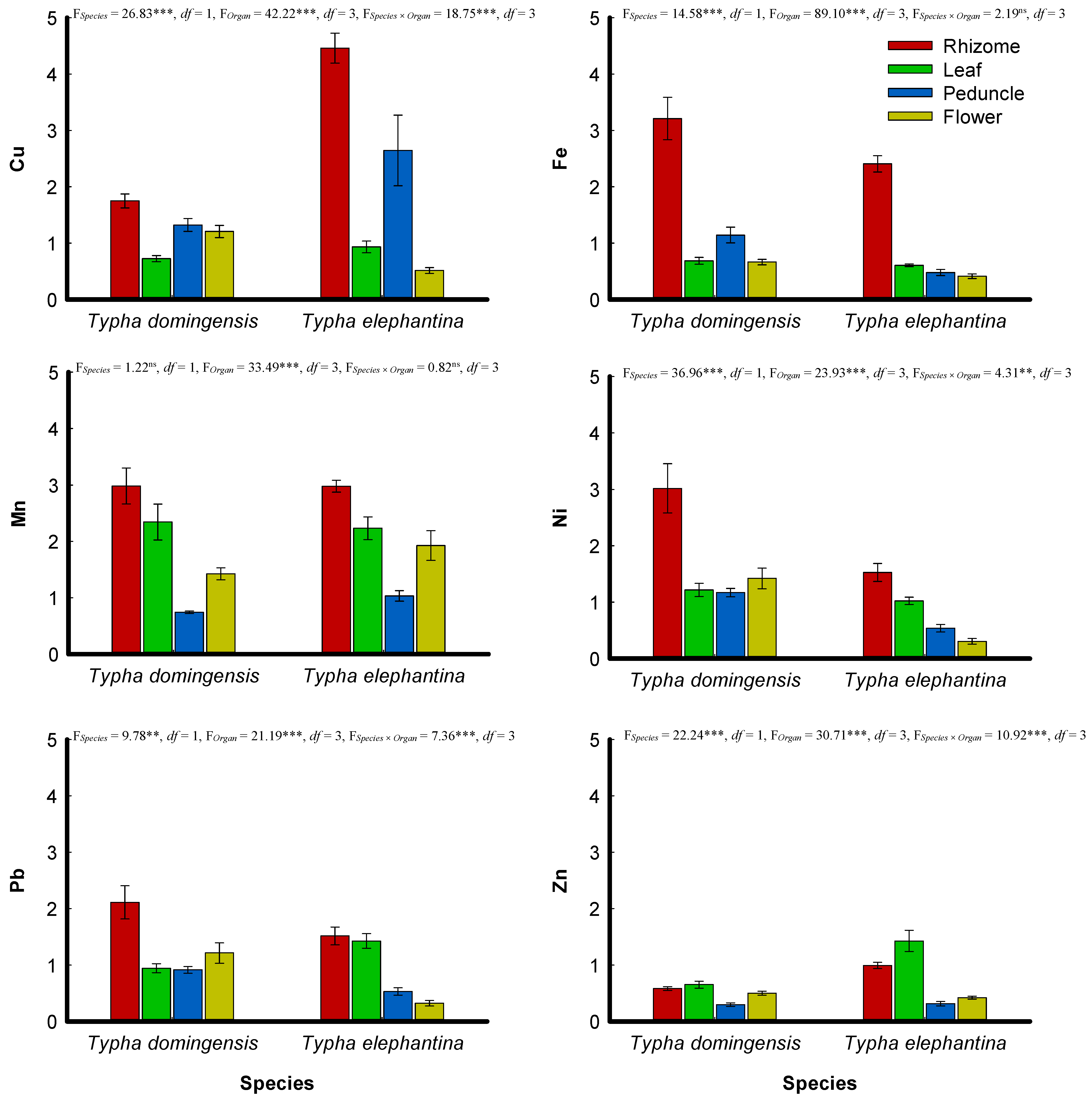
3.3. Bioaccumulation and Translocation of Metals
3.4. Plant–Sediment Correlations
3.5. Prediction Models
4. Discussion
4.1. Sediment Analysis
4.2. Plant Metals
4.3. Bioaccumulation and Translocation of Metals
4.4. Plant–Sediment Correlations
4.5. Prediction Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethical Statement
References
- Eid, E.M.; Galal, T.M.; Sewelam, N.A.; Talha, N.I.; Abdallah, S.M. Phytoremediation of heavy metals by four aquatic macrophytes and their potential use as contamination indicators: A comparative assessment. Environ. Sci. Pollut. Res. 2020, 27, 12138–12151. [Google Scholar] [CrossRef]
- Parveen, N.; Zaidi, S.; Danish, M. Support vector regression model for predicting the sorption capacity of lead (II). Perspect. Sci. 2016, 8, 629–631. [Google Scholar] [CrossRef] [Green Version]
- Gall, J.E.; Boyd, R.S.; Rajakaruna, N. Transfer of heavy metals through terrestrial food webs: A review. Environ. Monit. Assess. 2015, 187, 201. [Google Scholar] [CrossRef] [Green Version]
- Eid, E.M.; Shaltout, K.H.; Alamri, S.A.M.; Alrumman, S.A.; Sewelam, N.; Taher, M.A.; Hashem, M.; Mostafa, Y.S.; Ahmed, M.T. Prediction models founded on soil characteristics for the estimated uptake of nine metals by okra plant, Abelmoschus esculentus (L.) Moench., cultivated in agricultural soils modified with varying sewage sludge concentrations. Sustainability 2021, 13, 12356. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Knasmuller, S.; Gottmann, E.; Steinkellner, H.; Fomin, A.; Pickl, C.; Paschke, A.; God, R.; Kundi, M. Detection of genotoxic effects of heavy metal contaminated soils with plant bioassay. Mutat. Res.-Genet. Toxicol. Environ. Mutagenesis 1998, 420, 37–48. [Google Scholar] [CrossRef]
- Gupta, S.; Satpati, S.; Nayek, S.; Garai, D. Effect of wastewater irrigation on vegetables in relation to bioaccumulation of heavy metals and biochemical changes. Environ. Monit. Assess. 2009, 165, 169–177. [Google Scholar] [CrossRef]
- Bonanno, G.; Vymazal, J. Compartmentalization of potentially hazardous elements in macrophytes: Insights into capacity and efficiency of accumulation. J. Geochem. Explor. 2017, 181, 22–30. [Google Scholar] [CrossRef]
- Rezania, S.; Taib, S.M.; Din, M.F.M.; Dahalan, F.A.; Kamyab, H. Comprehensive review on phytotechnology: Heavy metals removal by diverse aquatic plants species from wastewater. J. Hazard. Mater. 2016, 318, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Vymazal, J.; Kröpfelová, L.; Švehla, J.; Chrastný, V.; Štíchová, J. Trace elements in Phragmites australis growing in constructed wetlands for treatment of municipal wastewater. Ecol. Eng. 2009, 35, 303–309. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H.; El-Sheikh, M.A.; Asaeda, T. Seasonal courses of nutrients and heavy metals in water, sediment and above- and below-ground Typha domingensis biomass in Lake Burullus (Egypt): Perspective for phytoremediation. Flora 2012, 207, 783–794. [Google Scholar] [CrossRef]
- Teuchies, J.; Jacobs, S.; Oosterlee, L.; Bervoets, L.; Meire, P. Role of plants in metal cycling in a tidal wetland: Implications for phytoremediation. Sci. Total. Environ. 2013, 445–446, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Everard, M. Nutrient cycling in wetlands. The Wetland Book, Structure and Function, Management, and Methods; Finlayson, C.M., Everard, M., Irvine, K., McInnes, R., Middleton, B., van Dam, A., Davidson, N.C., Eds.; Springer: The Netherlands, 2017; Available online: https://link.springer.com/referencework/10.1007/978-90-481-9659-3 (accessed on 10 December 2021).
- Kurilenko, V.V.; Osmolovskaya, N.G. Ecological-biogeochemical role of macrophytes in aquatic ecosystems of urbanized territories (an example of small bodies of St. Petersburg). Russ. J. Ecol. 2006, 37, 163–167. [Google Scholar] [CrossRef]
- Bonanno, G.; Vymazal, J.; Cirelli, G.L. Translocation, accumulation and bioindication of trace elements in wetland plants. Sci. Total. Environ. 2018, 631–632, 252–261. [Google Scholar] [CrossRef]
- Zeng, F.; Ali, S.; Zhang, H.; Ouyang, Y.; Qiu, B.; Wu, F.; Zhang, G. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants. Environ. Pollut. 2011, 159, 84–91. [Google Scholar] [CrossRef]
- Raguž, V.; Jarsjo, J.; Grolander, S.; Lindborg, R.; Avila, R. Plant uptake of elements in soil and pore water: Field observations versus model assumptions. J. Environ. Manag. 2013, 126, 147–156. [Google Scholar] [CrossRef]
- Waegeneers, N.; Ruttens, A.; De Temmerman, L. A dynamic model to calculate cadmium concentrations in bovine tissues from basic soil characteristics. Sci. Total. Environ. 2011, 409, 2815–2823. [Google Scholar] [CrossRef]
- Novotná, M.; Mikeš, O.; Komprdová, K. Development and comparison of regression models for the uptake of metals into various field crops. Environ. Pollut. 2015, 207, 357–364. [Google Scholar] [CrossRef]
- Bešter, P.K.; Lobnik, F.; Eržen, I.; Kastelec, D.; Zupan, M. Prediction of cadmium concentration in selected home-produced vegetables. Ecotoxicol. Environ. Saf. 2013, 96, 182–190. [Google Scholar] [CrossRef]
- Boshoff, M.; De Jonge, M.; Scheifler, R.; Bervoets, L. Predicting As, Cd, Cu, Pb and Zn levels in grasses (Agrostis sp. and Poa sp.) and stinging nettle (Urtica dioica) applying soil-plant transfer models. Sci. Total. Environ. 2014, 493, 862–871. [Google Scholar] [CrossRef]
- Eid, E.M.; Galal, T.M.; Shaltout, K.H.; El-Sheikh, M.A.; Asaeda, T.; Alatar, A.A.; Alfarhan, A.H.; Alharthi, A.; Alshehri, A.M.A.; Picó, Y.; et al. Biomonitoring potential of the native aquatic plant Typha domingensis by predicting trace metals accumulation in the Egyptian Lake Burullus. Sci. Total Environ. 2020, 714, 136603. [Google Scholar] [CrossRef] [PubMed]
- Farahat, E.A.; Mahmoud, W.F.; Awad, H.E.A.; Farrag, H.F.; Arshad, M.; Eid, E.M.; Fahmy, G.M. Prediction models for evaluating the uptake of heavy metals by the invasive grass Vossia cuspidata (Roxb.) Griff. in the River Nile, Egypt: A biomonitoring approach. Sustainability 2021, 13, 10558. [Google Scholar] [CrossRef]
- Singh, G.; Narwal, S.; Agnihotri, S. Typha elephantina Roxb.: A review on ethanomedicinal, morphological, phytochemical and pharmacological perspectives. Res. J. Pharm. Technol. 2020, 13, 5546–5550. [Google Scholar]
- Panich-Pat, T.; Pokethitiyook, P.; Kruatrachue, P.M.; Upatham, E.S.; Srinives, P.; Lanza, G.R. Removal of lead from contaminated soils by typha angustifolia. Water, Air, Soil Pollut. 2004, 155, 159–171. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt, Monocotyledons (Altismataceae-Orchidaceae); Al-Hadara Publishing: Cairo, Egypt, 2005; Volume 4. [Google Scholar]
- Qian, Y.; Gallagher, F.J.; Feng, H.; Wu, M. A geochemical study of toxic metal translocation in an urban brownfield wetland. Environ. Pollut. 2012, 166, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Eid, E.M.; Shaltout, K.H. Bioaccumulation and translocation of heavy metals by nine native plant species grown at a sewage sludge dump site. Int. J. Phytoremediation 2016, 18, 1075–1085. [Google Scholar] [CrossRef]
- Pandey, S.K.; Bhattacharya, T.; Chakraborty, S. Metal phytoremediation potential of naturally growing plants on fly ash dumpsite of Patratu thermal power station, Jharkhand, India. Int. J. Phytoremediation 2016, 18, 87–93. [Google Scholar] [CrossRef]
- UN-Habitat. Future Saudi Program: Taif CPI Profile; Ministry of Municipal and Rural Affairs: Riyadh, Saudi Arabia, 2019. [Google Scholar]
- Saleh, M.A.; Al-Sodany, Y.M.; Abdel Khalik, K.N.; Eid, E.M. Heavy metals accumulation and translocation by Typha elephantina Roxb. and Typha domingensis Pers. in an arid habitat: Perspectives for phytoremediation. World J. Adv. Res. Rev. 2019, 4, 44–53. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H.; Asaeda, T. Modeling growth dynamics of Typha domingensis (Pers.) Poir. ex Steud. in Lake Burullus, Egypt. Ecol. Model. 2012, 243, 63–72. [Google Scholar] [CrossRef]
- Baldantoni, D.; Alfani, A.; Tommasi, P.D.; Bartoli, G.; Virzo De Santo, A. Assessment of macro and microelement accumulation capability of two aquatic plants. Environ. Pollut. 2004, 130, 149–156. [Google Scholar] [CrossRef]
- Rzymski, P.; Niedzielski, P.; Klimaszyk, P.; Poniedziałek, B. Bioaccumulation of selected metals in bivalves (Unionide) and Phragmites australis inhabiting a municipal water reservoir. Environ. Monit. Assess. 2014, 186, 3199–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: London, UK, 1989. [Google Scholar]
- Statistical Package for the Social Sciences. In SPSS Base 15.0 User’s Guide; SPSS Inc.: Chicago, IL, USA, 2006.
- Balkhair, K.S.; Ashraf, M.A. Field accumulation risks of heavy metals in a soil and vegetable crop irrigated with sewage water in western region of Saudi Arabia. Saudi J. Biol. Sci. 2016, 23, 32–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiroma, T.M.; Ebewele, R.O.; Hymore, F.K. Comparative assessment of heavy metal levels in soil, vegetables and urban grey waste water used for irrigation in Yola and Kano. Int. Refereed J. Eng. Sci. 2014, 3, 1–9. [Google Scholar]
- Du Laing, G.; Van de Moortel, A.M.K.; Moors, W.; De Grauwe, P.; Meers, E.; Tack, F.M.G.; Verloo, M.G. Factors affecting metal concentrations in reed plants (Phragmites australis) of intertidal marshes in the Scheldt estuary. Ecol. Eng. 2009, 35, 310–318. [Google Scholar] [CrossRef]
- Nawab, J.; Khan, S.; Shah, M.T.; Khan, K.; Huang, Q.; Ali, R. Quantification of heavy metals in mining affected soil and their bioaccumulation in native plant species. Int. J. Phytoremediation 2015, 17, 801–813. [Google Scholar] [CrossRef]
- Eid, E.M.; Shaltout, K.H.; Al-Sodany, Y.M.; Haroun, S.A.; Galal, T.M.; Ayed, H.; Khedher, K.M.; Jensen, K. Common reed (Phragmites australis (Cav.) Trin. ex Steudel) as a candidate for predicting heavy metal contamination in Lake Burullus, Egypt: A biomonitoring approach. Ecol. Eng. 2020, 148, 105787. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Galal, T.M.; El-Komy, T.M. Evaluation of the nutrient status of some hydrophytes in the water courses of Nile Delta, Egypt. Ecol. Mediterr. 2010, 36, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Galal, T.M.; Shehata, H.S. Growth and nutrients accumulation potentials of giant reed (Arundo donax L.) in different habitats in Egypt. Int. J. Phytoremediation 2016, 18, 1221–1230. [Google Scholar] [CrossRef]
- Galal, T.M.; Gharib, F.A.; Ghazi, S.M.; Mansour, K.H. Phytostabilization of heavy metals by the emergent macrophyte Vossia cuspidata (Roxb.) Griff.: A phytoremediation approach. Int. J. Phytoremediat. 2017, 19, 992–999. [Google Scholar] [CrossRef]
- Bonanno, G.; Borg, J.A.; Di Martino, V. Levels of heavy metals in wetland and marine vascular plants and their biomonitoring potential: A comparative assessment. Sci. Total. Environ. 2017, 576, 796–806. [Google Scholar] [CrossRef]
- Singh, S.; Saxena, R.; Pandey, K.; Bhatt, K.; Sinha, S. Response of antioxidants in sunflower (Helianthus annuus L.) grown on different amendments of tannery sludge: Its metal accumulation potential. Chemosphere 2004, 57, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Eid, E.M.; Shaltout, K.H. Monthly variations of trace elements accumulation and distribution in above- and below-ground biomass of Phragmites australis (Cav.) Trin. ex Steudel in Lake Burullus (Egypt): A biomonitoring application. Ecol. Eng. 2014, 73, 17–25. [Google Scholar] [CrossRef]
- Bonanno, G. Comparative performance of trace element bioaccumulation and biomonitoring in the plant species Typha domingensis, Phragmites australis and Arundo donax. Ecotoxicol. Environ. Saf. 2013, 97, 124–130. [Google Scholar] [CrossRef]
- Markert, B. Presence and significance of naturally occurring chemical elements of the periodic system in the plant organism and consequences for future investigations on inorganic environmental chemistry in ecosystems. Vegetatio 1992, 103, 1–30. [Google Scholar]
- Carranza-Álvarez, C.; Alonso-Castro, A.J.; Alfaro-De La Torre, M.C.; García-De La Cruz, R.F. Accumulation and distribution of heavy metals in Scirpus americanus and Typha latifolia from an artificial lagoon in San Luis Potosí, México. Water Air Soil Pollut. 2008, 188, 297–309. [Google Scholar] [CrossRef]
- Sasmaz, A.; Obek, E.; Hasar, H. The accumulation of heavy metals in Typha latifolia L. grown in a stream carrying secondary effluent. Ecol. Eng. 2008, 33, 278–284. [Google Scholar] [CrossRef]
- Bonanno, G.; Lo Giudice, R. Heavy metal bioaccumulation by the organs of Phragmites australis (common reed) and their potential use as contamination indicators. Ecol. Indic. 2010, 10, 639–645. [Google Scholar] [CrossRef]
- Basta, N.T.; Ryan, J.A.; Chaney, R.L. Trace element chemistry in residual-treated soil: Key concepts and metal bioavailability. J. Environ. Qual. 2005, 34, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branzini, A.; González, R.S.; Zubillaga, M. Absorption and translocation of copper, zinc and chromium by Sesbania virgata. J. Environ. Manag. 2012, 102, 50–54. [Google Scholar] [CrossRef]
- Environmental Protection Agency. Framework for Metal Risk Assessment; Environmental Protection Agency, Office of the Science Advisor: Washington, DC, USA, 2007. [Google Scholar]
- Zhang, W.; Cai, Y.; Tu, C.; Ma, L.Q. Arsenic speciation and distribution in an arsenic hyperaccumulating plant. Sci. Total. Environ. 2002, 300, 167–177. [Google Scholar] [CrossRef]
- Bothe, H.; Słomka, A. Divergent biology of facultative heavy metal plants. J. Plant Physiol. 2017, 219, 45–61. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Agrawal, M. Variations in heavy metal accumulation, growth and yield of rice plants grown at different sewage sludge amendment rates. Ecotoxicol. Environ. Saf. 2010, 73, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Kalis, E.J.J.; Temminghoff, E.J.M.; Town, R.M.; Unsworth, E.R.; Van Riemsdijk, W.H. Relationship between metal speciation in soil solution and metal adsorption at the root surface of ryegrass. J. Environ. Qual. 2008, 37, 2221–2231. [Google Scholar] [CrossRef]
- Lyubenova, L.; Pongrac, P.; Vogel-Mikuš, K.; Kukek Mezek, G.; Vavpetič, P.; Grlj, N.; Regvar, M.; Pelicon, P.; Schröder, P. The fate of arsenic, cadmium and lead in Typha latifolia: A case study on the applicability of micro-PIXE in plant ionomics. J. Hazard. Mater. 2013, 248–249, 371–378. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Fairbrother, A.; Wenstel, R.; Sappington, S.; Wood, W. Framework for metals risk assessment. Ecotoxicol. Environ. Saf. 2007, 68, 145–227. [Google Scholar] [CrossRef]
- Vardanyan, L.G.; Ingole, B.S. Studies on heavy metal accumulation in aquatic macrophytes from Sevan (Armenia) and Carambolim (India) lake systems. Environ. Int. 2006, 32, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.E.; Long, X.X.; Ye, H.B.; He, Z.L.; Calvert, D.V.; Stoffella, P.J. Cadmium tolerance and hyperaccumulation in a new Zn-hyperaccumulating plant species (Sedum alfredii Hance). Plant Soil 2004, 259, 181–189. [Google Scholar] [CrossRef]
- Weis, J.S.; Glover, T.; Weis, P. Interactions of metals affect their distribution in tissues of Phragmites australis. Environ. Pollut. 2004, 131, 409–415. [Google Scholar] [CrossRef]
- Bello, A.O.; Tawabini, B.S.; Khalil, A.B.; Boland, C.R.; Saleh, T.A. Phytoremediation of cadmium-, lead- and nickel-contaminated water by Phragmites australis in hydroponic systems. Ecol. Eng. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Aksoy, A.; Demirezen, D.; Duman, F. Bioaccumulation, detection and analyses of heavy metal pollution in sultan marsh and its environment. Water Air Soil Pollut. 2005, 164, 241–255. [Google Scholar] [CrossRef]
- Priju, C.P.; Narayana, A.C. Heavy and trace metals in Vembanad Lake sediments. Int. J. Environ. Res. 2007, 1, 280–289. [Google Scholar]
- Kumar, V.; Singh, J.; Kumar, P. Heavy metal uptake by water lettuce (Pistia stratiotes L.) from paper mill effluent (PME): Experimental and prediction modelling studies. Environ. Sci. Pollut. Res. 2019, 26, 14400–14413. [Google Scholar] [CrossRef] [PubMed]
- Luna, A.S.; Gonzaga, F.B.; Werickson, F.C.; Lima, I.C.A. A comparison of different strategies in multivariate regression models for the direct determination of Mn, Cr, and Ni in steel samples using laser-induced breakdown spectroscopy. Spectrochim. Acta Part B At. Spectrosc. 2018, 139, 20–26. [Google Scholar] [CrossRef]
- Tudoreanu, L.; Phillips, C.J.C. Empirical models of cadmium accumulation in maize, rye grass and soya bean plants. J. Sci. Food Agric. 2004, 84, 845–852. [Google Scholar] [CrossRef]
 ). Td: Typha domingensis; Te: Typha elephantina.
). Td: Typha domingensis; Te: Typha elephantina.
). Td: Typha domingensis; Te: Typha elephantina.
). Td: Typha domingensis; Te: Typha elephantina.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Value | pH | Metal Concentration in Sediment (mg/kg) | |||||
|---|---|---|---|---|---|---|---|
| Cu | Fe | Mn | Ni | Pb | Zn | ||
| Typha domingensis | |||||||
| Minimum | 7.80 | 1.88 | 2.22 | 2.92 | 0.09 | 0.12 | 0.46 |
| Maximum | 8.50 | 3.19 | 12.15 | 4.64 | 0.13 | 0.15 | 2.23 |
| Mean (n = 48) | 8.06 | 2.39 | 4.76 | 4.01 | 0.11 | 0.14 | 1.07 |
| CV (%) | 2.41 | 17.73 | 75.92 | 14.29 | 12.16 | 7.22 | 54.95 |
| Typha elephantina | |||||||
| Minimum | 7.80 | 2.01 | 2.28 | 3.35 | 0.10 | 0.13 | 0.72 |
| Maximum | 8.70 | 5.38 | 3.55 | 4.00 | 0.20 | 0.24 | 1.07 |
| Mean (n = 48) | 8.29 | 3.74 | 2.93 | 3.73 | 0.14 | 0.18 | 0.88 |
| CV (%) | 2.59 | 38.26 | 15.56 | 6.19 | 28.98 | 25.12 | 14.93 |
| t-value | 5.98 | 7.09 | 3.72 | 3.29 | 5.15 | 5.78 | 1.98 |
| p | 0.000 | 0.000 | 0.001 | 0.002 | 0.000 | 0.000 | 0.054 |
| Organ | Value | Metal Concentration (mg/kg) | |||||
|---|---|---|---|---|---|---|---|
| Cu | Fe | Mn | Ni | Pb | Zn | ||
| Roots | Typha domingensis | ||||||
| Minimum | 8.70 | 699.98 | 69.75 | 3.05 | 3.70 | 26.58 | |
| Maximum | 40.60 | 2003.05 | 183.10 | 282.85 | 304.40 | 84.40 | |
| Mean (n = 48) | 20.09 | 1209.05 | 118.16 | 141.97 | 149.42 | 52.54 | |
| CV (%) | 53.33 | 35.66 | 37.38 | 86.20 | 82.80 | 40.88 | |
| Typha elephantina | |||||||
| Minimum | 8.77 | 551.97 | 35.93 | 5.57 | 5.13 | 11.17 | |
| Maximum | 39.40 | 2281.10 | 117.68 | 226.88 | 205.90 | 84.18 | |
| Mean (n = 48) | 20.40 | 1207.41 | 78.92 | 97.09 | 90.87 | 33.59 | |
| CV (%) | 53.85 | 49.38 | 40.99 | 84.53 | 82.71 | 70.96 | |
| t-value | 0.23 | 0.02 | 11.02 | 3.51 | 4.37 | 7.92 | |
| p | 0.817 | 0.988 | 0.000 | 0.001 | 0.000 | 0.000 | |
| Rhizomes | Typha domingensis | ||||||
| Minimum | 12.65 | 980.50 | 157.80 | 25.00 | 22.65 | 13.35 | |
| Maximum | 38.88 | 5555.03 | 642.50 | 148.75 | 144.05 | 41.35 | |
| Mean (n = 48) | 29.39 | 3154.89 | 289.38 | 108.29 | 102.46 | 27.89 | |
| CV (%) | 31.63 | 53.92 | 59.02 | 36.91 | 37.43 | 33.79 | |
| Typha elephantina | |||||||
| Minimum | 56.60 | 1309.50 | 103.77 | 17.93 | 16.93 | 18.18 | |
| Maximum | 91.18 | 4248.80 | 279.33 | 127.60 | 118.23 | 52.00 | |
| Mean (n = 48) | 71.69 | 2517.53 | 218.55 | 72.92 | 68.06 | 27.61 | |
| CV (%) | 16.82 | 39.82 | 28.71 | 50.95 | 50.13 | 43.80 | |
| t-value | 26.96 | 1.85 | 2.22 | 10.85 | 9.95 | 0.12 | |
| p | 0.000 | 0.071 | 0.031 | 0.000 | 0.000 | 0.907 | |
| Peduncles | Typha domingensis | ||||||
| Minimum | 9.20 | 339.15 | 35.90 | 31.90 | 30.80 | 6.50 | |
| Maximum | 47.10 | 2362.10 | 149.00 | 304.70 | 310.70 | 52.55 | |
| Mean (n = 36) | 26.02 | 1276.01 | 94.61 | 156.65 | 147.07 | 17.53 | |
| CV (%) | 58.43 | 71.04 | 43.95 | 71.31 | 74.64 | 101.87 | |
| Typha elephantina | |||||||
| Minimum | 3.40 | 102.11 | 44.43 | 1.37 | 2.07 | 2.33 | |
| Maximum | 100.95 | 1247.93 | 147.10 | 140.00 | 127.90 | 18.80 | |
| Mean (n = 36) | 39.19 | 652.32 | 79.37 | 77.02 | 71.62 | 10.62 | |
| CV (%) | 100.07 | 69.85 | 45.31 | 80.34 | 79.94 | 67.89 | |
| t-value | 2.17 | 3.49 | 1.69 | 3.89 | 3.65 | 2.88 | |
| p | 0.036 | 0.001 | 0.099 | 0.000 | 0.001 | 0.006 | |
| Flowers | Typha domingensis | ||||||
| Minimum | 10.65 | 390.00 | 34.90 | 41.50 | 41.10 | 14.85 | |
| Maximum | 39.80 | 1592.35 | 293.20 | 263.20 | 275.40 | 58.75 | |
| Mean (n = 36) | 20.53 | 786.72 | 181.31 | 123.07 | 127.96 | 27.19 | |
| CV (%) | 49.95 | 54.67 | 51.51 | 64.26 | 65.08 | 62.48 | |
| Typha elephantina | |||||||
| Minimum | 2.40 | 144.70 | 55.23 | 1.10 | 3.23 | 10.07 | |
| Maximum | 40.97 | 926.00 | 231.30 | 96.83 | 92.63 | 15.60 | |
| Mean (n = 36) | 14.90 | 529.63 | 130.27 | 32.26 | 31.81 | 12.77 | |
| CV (%) | 99.74 | 59.53 | 55.35 | 106.53 | 101.69 | 15.89 | |
| t-value | 1.67 | 2.56 | 2.05 | 6.54 | 6.62 | 5.58 | |
| p | 0.103 | 0.015 | 0.047 | 0.000 | 0.000 | 0.000 | |
| Leaves | Typha domingensis | ||||||
| Minimum | 1.15 | 162.25 | 87.75 | 2.45 | 2.40 | 5.95 | |
| Maximum | 21.80 | 1181.75 | 691.15 | 177.55 | 172.80 | 81.85 | |
| Mean (n = 48) | 13.68 | 721.90 | 245.65 | 103.86 | 101.32 | 35.28 | |
| CV (%) | 49.53 | 45.63 | 84.39 | 60.69 | 61.09 | 80.08 | |
| Typha elephantina | |||||||
| Minimum | 9.15 | 392.60 | 91.40 | 4.20 | 16.47 | 6.68 | |
| Maximum | 22.80 | 1058.85 | 187.07 | 234.10 | 214.30 | 70.46 | |
| Mean (n = 48) | 13.64 | 663.68 | 142.40 | 86.66 | 83.17 | 31.72 | |
| CV (%) | 34.40 | 34.39 | 22.55 | 85.49 | 78.36 | 66.39 | |
| t-value | 0.05 | 1.66 | 3.33 | 2.72 | 2.85 | 0.57 | |
| p | 0.961 | 0.103 | 0.002 | 0.009 | 0.007 | 0.573 | |
| Equation | R2 | ME | MNAE | Student’s t-Test | |
|---|---|---|---|---|---|
| t-Value | p | ||||
| Roots | |||||
| Cu = −85.76 + (5.08 × CuSediment) + (11.62 × pH) | 0.111 | 0.445 | 0.173 | 1.517 | 0.163 |
| Fe = 6962.87 − (72.92 × FeSediment) − (670.61 × pH) | 0.286 ** | 0.798 | 0.102 | 1.155 | 0.273 |
| Mn = −51.74 + (17.56 × MnSediment) + (12.35 × pH) | 0.070 | 0.391 | 0.183 | 1.730 | 0.115 |
| Ni = 38.52 − (7601.65 × NiSediment) + (118.12 × pH) | 0.754 *** | 0.979 | 0.028 | 0.190 | 0.853 |
| Pb = 239.09 − (3783.22 × PbSediment) + (54.57 × pH) | 0.123 | 0.462 | 0.155 | 1.496 | 0.164 |
| Zn = −165.19 − (10.52 × ZnSediment) + (28.39 × pH) | 0.222 ** | 0.691 | 0.119 | 1.360 | 0.201 |
| Rhizomes | |||||
| Cu = −168.31 + (10.60 × CuSediment) + (21.38 × pH) | 0.569 *** | 0.900 | 0.062 | 0.624 | 0.545 |
| Fe = −21,294.84 + (455.45 × FeSediment) + (2763.73 × pH) | 0.739 *** | 0.945 | 0.033 | 0.282 | 0.783 |
| Mn = 1696.59 − (210.24 × MnSediment) − (70.10 × pH) | 0.570 *** | 0.916 | 0.054 | 0.436 | 0.673 |
| Ni = −604.81 − (1430.48 × NiSediment) + (108.26 × pH) | 0.525 *** | 0.876 | 0.068 | 0.898 | 0.388 |
| Pb = −553.23 − (578.10 × PbSediment) + (91.36 × pH) | 0.292 *** | 0.811 | 0.081 | 1.065 | 0.310 |
| Zn = −61.67 − (0.70 × ZnSediment) + (11.20 × pH) | 0.065 | 0.353 | 0.267 | 1.743 | 0.112 |
| Peduncles | |||||
| Cu = 190.46 − (10.68 × CuSediment) − (17.07 × pH) | 0.170 * | 0.610 | 0.133 | 1.384 | 0.194 |
| Fe = 7590.06 − (185.50 × FeSediment) − (703.10 × pH) | 0.134 | 0.556 | 0.152 | 1.463 | 0.178 |
| Mn = −84.28 + (3.89 × MnSediment) + (20.02 × pH) | 0.008 | 0.100 | 0.346 | 1.939 | 0.079 |
| Ni = 1256.03 − (7396.26 × NiSediment) − (35.22 × pH) | 0.921 *** | 0.982 | 0.008 | 0.164 | 0.872 |
| Pb = 283.88 + (5222.91 × PbSediment) − (105.68 × pH) | 0.275 ** | 0.788 | 0.105 | 1.247 | 0.244 |
| Zn = −108.79 − (46.12 × ZnSediment) + (20.29 × pH) | 0.522 *** | 0.854 | 0.069 | 0.974 | 0.351 |
| Flowers | |||||
| Cu = 106.31 + (2.43 × CuSediment) − (11.30 × pH) | 0.036 | 0.240 | 0.272 | 1.874 | 0.094 |
| Fe = 3663.54 − (16.50 × FeSediment) − (347.84 × pH) | 0.026 | 0.195 | 0.299 | 1.928 | 0.086 |
| Mn = 1033.07 − (177.63 × MnSediment) − (12.55 × pH) | 0.386 *** | 0.814 | 0.081 | 1.000 | 0.343 |
| Ni = 472.96 − (4068.78 × NiSediment) + (12.04 × pH) | 0.536 *** | 0.880 | 0.067 | 0.840 | 0.419 |
| Pb = −567.01 + (5804.10 × PbSediment) − (13.06 × pH) | 0.485 *** | 0.825 | 0.074 | 0.977 | 0.350 |
| Zn = −86.52 − (46.99 × ZnSediment) + (18.83 × pH) | 0.595 *** | 0.928 | 0.047 | 0.361 | 0.725 |
| Leaves | |||||
| Cu = −127.11 − (0.56 × CuSediment) + (17.63 × pH) | 0.245 ** | 0.750 | 0.109 | 1.356 | 0.208 |
| Fe = −1131.89 − (60.78 × FeSediment) + (265.80 × pH) | 0.570 *** | 0.901 | 0.062 | 0.622 | 0.550 |
| Mn = 83.76 − (47.56 × MnSediment) + (43.71 × pH) | 0.013 | 0.140 | 0.346 | 1.937 | 0.084 |
| Ni = −406.67 − (3649.16 × NiSediment) + (113.86 × pH) | 0.754 *** | 0.967 | 0.032 | 0.247 | 0.809 |
| Pb = −462.69 − (1131.34 × PbSediment) + (89.60 × pH) | 0.154 * | 0.578 | 0.139 | 1.445 | 0.182 |
| Zn = 318.75 − (41.81 × ZnSediment) − (29.64 × pH) | 0.616 *** | 0.939 | 0.041 | 0.336 | 0.743 |
| Equation | R2 | ME | MNAE | Student’s t-Test | |
|---|---|---|---|---|---|
| t-Value | p | ||||
| Roots | |||||
| Cu = −141.88 + (3.56 × CuSediment) + (17.98 × pH) | 0.232 ** | 0.770 | 0.078 | 1.204 | 0.254 |
| Fe = 911.54 − (505.10 × FeSediment) + (214.12 × pH) | 0.165 * | 0.615 | 0.119 | 1.459 | 0.172 |
| Mn = −448.87 + (86.00 × MnSediment) + (24.97 × pH) | 0.443 *** | 0.888 | 0.058 | 0.644 | 0.533 |
| Ni = −1008.38 − (273.78 × NiSediment) + (138.09 × pH) | 0.126 * | 0.342 | 0.165 | 1.912 | 0.082 |
| Pb = −892.57 − (148.37 × PbSediment) + (121.77 × pH) | 0.115 | 0.221 | 0.201 | 2.136 | 0.061 |
| Zn = −241.09 + (136.91 × ZnSediment) + (18.63 × pH) | 0.623 *** | 0.949 | 0.036 | 0.144 | 0.889 |
| Rhizomes | |||||
| Cu = 126.50 + (7.02 × CuSediment) − (9.78 × pH) | 0.816 *** | 0.970 | 0.034 | 0.137 | 0.893 |
| Fe = 12,064.24 + (715.89 × FeSediment) − (1404.54 × pH) | 0.231 ** | 0.675 | 0.084 | 1.211 | 0.251 |
| Mn = −391.29 + (137.53 × MnSediment) + (11.69 × pH) | 0.266 ** | 0.830 | 0.071 | 0.915 | 0.384 |
| Ni = −134.81 − (685.53 × NiSediment) + (36.91 × pH) | 0.556 *** | 0.905 | 0.044 | 0.288 | 0.779 |
| Pb = −118.83 − (574.13 × PbSediment) + (34.67 × pH) | 0.521 *** | 0.899 | 0.055 | 0.430 | 0.675 |
| Zn = −147.20 + (46.16 × ZnSediment) + (16.20 × pH) | 0.364 *** | 0.886 | 0.061 | 0.670 | 0.520 |
| Peduncles | |||||
| Cu = −34.96 − (27.66 × CuSediment) + (22.61 × pH) | 0.931 *** | 0.996 | 0.029 | 0.019 | 0.985 |
| Fe = −6254.05 − (397.58 × FeSediment) + (970.87 × pH) | 0.470 *** | 0.895 | 0.057 | 0.493 | 0.634 |
| Mn = −196.19 − (17.98 × MnSediment) + (41.48 × pH) | 0.060 | 0.100 | 0.457 | 2.227 | 0.053 |
| Ni = −1103.38 + (938.76 × NiSediment) + (127.71 × pH) | 0.577 *** | 0.926 | 0.043 | 0.280 | 0.785 |
| Pb = −1043.30 + (753.06 × PbSediment) + (119.85 × pH) | 0.556 *** | 0.900 | 0.051 | 0.420 | 0.683 |
| Zn = −72.47 − (17.71 × ZnSediment) + (11.99 × pH) | 0.165 * | 0.576 | 0.148 | 1.663 | 0.125 |
| Flowers | |||||
| Cu = −239.94 + (0.21 × CuSediment) + (30.70 × pH) | 0.193 * | 0.647 | 0.106 | 1.454 | 0.180 |
| Fe = −4581.89 − (127.53 × FeSediment) + (661.52 × pH) | 0.287 ** | 0.880 | 0.065 | 0.797 | 0.446 |
| Mn = 562.43 − (149.02 × MnSediment) + (15.42 × pH) | 0.247 ** | 0.775 | 0.074 | 1.147 | 0.281 |
| Ni = −420.22 + (124.09 × NiSediment) + (52.72 × pH) | 0.140 * | 0.391 | 0.159 | 1.845 | 0.092 |
| Pb = −391.59 + (100.75 × PbSediment) + (49.18 × pH) | 0.136 * | 0.386 | 0.161 | 1.848 | 0.092 |
| Zn = −24.82 − (4.45 × ZnSediment) + (5.03 × pH) | 0.284 ** | 0.850 | 0.067 | 0.830 | 0.424 |
| Leaves | |||||
| Cu = −6.84 − (1.47 × CuSediment) + (3.13 × pH) | 0.262 ** | 0.794 | 0.074 | 0.924 | 0.375 |
| Fe = −810.57 − (178.38 × FeSediment) + (240.85 × pH) | 0.206 ** | 0.675 | 0.087 | 1.283 | 0.226 |
| Mn = −297.27 + (1.14 × MnSediment) + (52.53 × pH) | 0.124 * | 0.297 | 0.167 | 1.995 | 0.077 |
| Ni = −140.59 + (499.08 × NiSediment) + (18.78 × pH) | 0.088 | 0.129 | 0.359 | 2.141 | 0.056 |
| Pb = −158.73 + (570.43 × PbSediment) + (17.14 × pH) | 0.161 * | 0.430 | 0.151 | 1.828 | 0.095 |
| Zn = −242.09 + (1.50 × ZnSediment) + (32.87 × pH) | 0.113 | 0.167 | 0.239 | 2.141 | 0.061 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Sodany, Y.M.; Saleh, M.A.; Arshad, M.; Abdel Khalik, K.N.; Al-Bakre, D.A.; Eid, E.M. Regression Models to Estimate Accumulation Capability of Six Metals by Two Macrophytes, Typha domingensis and Typha elephantina, Grown in an Arid Climate in the Mountainous Region of Taif, Saudi Arabia. Sustainability 2022, 14, 1. https://doi.org/10.3390/su14010001
Al-Sodany YM, Saleh MA, Arshad M, Abdel Khalik KN, Al-Bakre DA, Eid EM. Regression Models to Estimate Accumulation Capability of Six Metals by Two Macrophytes, Typha domingensis and Typha elephantina, Grown in an Arid Climate in the Mountainous Region of Taif, Saudi Arabia. Sustainability. 2022; 14(1):1. https://doi.org/10.3390/su14010001
Chicago/Turabian StyleAl-Sodany, Yassin M., Muneera A. Saleh, Muhammad Arshad, Kadry N. Abdel Khalik, Dhafer A. Al-Bakre, and Ebrahem M. Eid. 2022. "Regression Models to Estimate Accumulation Capability of Six Metals by Two Macrophytes, Typha domingensis and Typha elephantina, Grown in an Arid Climate in the Mountainous Region of Taif, Saudi Arabia" Sustainability 14, no. 1: 1. https://doi.org/10.3390/su14010001