
Sustainable Biological Ammonia Production towards a Carbon-Free Society


Abstract
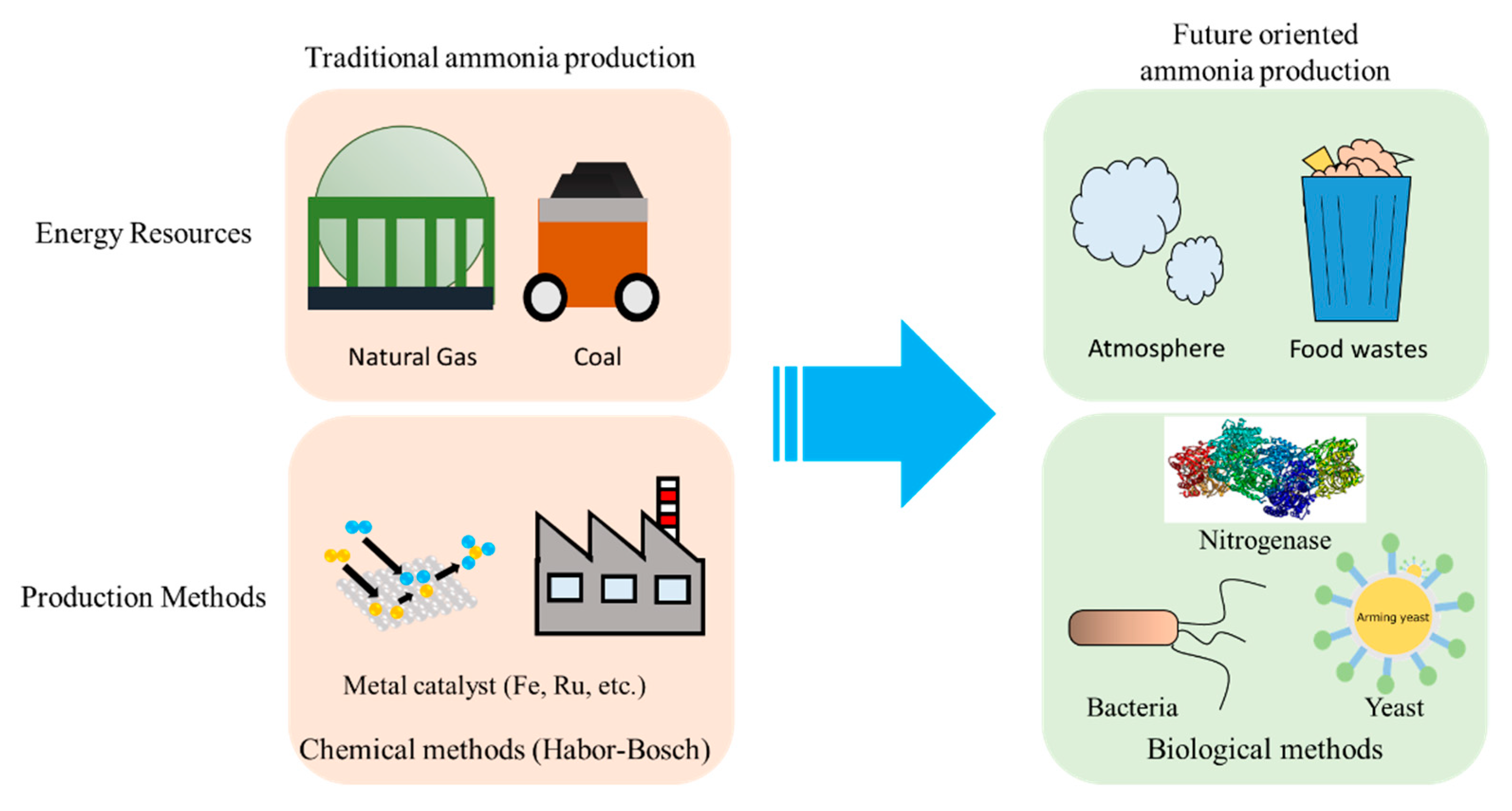
:1. Introduction
2. Ammonia Production Using Nitrogenase
3. Ammonia Production by Metabolic and Cell Engineering
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tortell, P.D. Earth 2020: Science, society, and sustainability in the Anthropocene. Proc. Natl. Acad. Sci. USA 2020, 117, 8683–8691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capua, I.; Giovannini, E. Coding system to track research progress towards SDGs. Nature 2019, 572, 178. [Google Scholar] [CrossRef]
- Demirhan, C.D.; Tso, W.W.; Powell, J.B.; Pistikopoulos, E.N. Sustainable ammonia production through process synthesis and global optimization. Aiche J. 2019, 65, e16498. [Google Scholar] [CrossRef]
- Mitsos, A.; Asprion, N.; Floudas, C.A.; Bortz, M.; Baldea, M.; Bonvin, D.; Caspari, A.; Schafer, P. Challenges in process optimization for new feedstocks and energy sources. Comput. Chem. Eng. 2018, 113, 209–221. [Google Scholar] [CrossRef]
- Wang, L.; Xia, M.K.; Wang, H.; Huang, K.F.; Qian, C.X.; Maravelias, C.T.; Ozin, G.A. Greening Ammonia toward the Solar Ammonia Refinery. Joule 2018, 2, 1055–1074. [Google Scholar] [CrossRef] [Green Version]
- Valera-Medina, A.; Amer-Hatem, F.; Azad, A.K.; Dedoussi, I.C.; de Joannon, M.; Fernandes, R.X.; Glarborg, P.; Hashemi, H.; He, X.; Mashruk, S.; et al. Review on Ammonia as a Potential Fuel: From Synthesis to Economics. Energy Fuel 2021, 35, 6964–7029. [Google Scholar] [CrossRef]
- Graziano, M.; Lecca, P.; Musso, M. Historic paths and future expectations: The macroeconomic impacts of the offshore wind technologies in the UK. Energy Policy 2017, 108, 715–730. [Google Scholar] [CrossRef]
- Elishav, O.; Mosevitzky Lis, B.; Miller, E.M.; Arent, D.J.; Valera-Medina, A.; Grinberg Dana, A.; Shter, G.E.; Grader, G.S. Progress and Prospective of Nitrogen-Based Alternative Fuels. Chem. Rev. 2020, 120, 5352–5436. [Google Scholar] [PubMed]
- Baliban, R.C.; Elia, J.A.; Floudas, C.A. Biomass and Natural Gas to Liquid Transportation Fuels: Process Synthesis, Global Optimization, and Topology Analysis. Ind. Eng. Chem. Res. 2013, 52, 3381–3406. [Google Scholar] [CrossRef]
- Lynd, L.R.; Larson, E.; Greene, N.; Laser, M.; Sheehan, J.; Dale, B.E.; McLaughlin, S.; Wang, M. The role of biomass in America’s energy future: Framing the analysis. Biofuels Bioprod. Biorefin. 2009, 3, 113–123. [Google Scholar] [CrossRef]
- Paritosh, K.; Kushwaha, S.K.; Yadav, M.; Pareek, N.; Chawade, A.; Vivekanand, V. Food Waste to Energy: An Overview of Sustainable Approaches for Food Waste Management and Nutrient Recycling. BioMed Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Erdemir, D.; Dincer, I. A perspective on the use of ammonia as a clean fuel: Challenges and solutions. Int. J. Energy Res. 2021, 45, 4827–4834. [Google Scholar] [CrossRef]
- Fedoruk, M.J.; Bronstein, R.; Kerger, B.D. Ammonia exposure and hazard assessment for selected household cleaning product uses. J. Expo. Sci. Environ. Epidemiol. 2005, 15, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Huang, S.M.; Shao, Y.Y.; Chen, J.C. Thermodynamic analysis of a new ammonia-water power cycle. Energy Rep. 2020, 6, 567–573. [Google Scholar] [CrossRef]
- Wakida, T.; Tokuyama, T.; Doi, C.; Lee, M.; Jeong, D.S.; Ishida, S. Mechanical properties of polyester/cotton and polyester/rayon fabrics treated with ammonia-gas. Sen-I Gakkaishi 2004, 60, 34–37. [Google Scholar] [CrossRef] [Green Version]
- Muller, T.E.; Beller, M. Metal-Initiated Amination of Alkenes and Alkynes. Chem. Rev. 1998, 98, 675–704. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jeong, J.; Fujita, K.I.; Yamamoto, A.; Yoshida, H. Anti-Markovnikov Hydroamination of Alkenes with Aqueous Ammonia by Metal-Loaded Titanium Oxide Photocatalyst. J. Am. Chem. Soc. 2020, 142, 12708–12714. [Google Scholar] [CrossRef] [PubMed]
- Miura, D.; Tezuka, T. A comparative study of ammonia energy systems as a future energy carrier, with particular reference to vehicle use in Japan. Energy 2014, 68, 428–436. [Google Scholar] [CrossRef]
- Green, L. An Ammonia Energy Vector for the Hydrogen Economy. Int. J. Hydrogen Energy 1982, 7, 355–359. [Google Scholar] [CrossRef]
- Mukherjee, P.; Rao, V.V. Design and development of high temperature superconducting magnetic energy storage for power applications—A review. Physics C 2019, 563, 67–73. [Google Scholar] [CrossRef]
- Rehman, S.; Al-Hadhrami, L.M.; Alam, M.M. Pumped hydro energy storage system: A technological review. Renew. Sustain. Energy Rev. 2015, 44, 586–598. [Google Scholar] [CrossRef]
- Gur, T.M. Review of electrical energy storage technologies, materials and systems: Challenges and prospects for large-scale grid storage. Energy Environ. Sci. 2018, 11, 3055. [Google Scholar] [CrossRef]
- Al Shaqsi, A.Z.; Sopian, K.; Al-Hinai, A. Review of energy storage services, applications, limitations, and benefits. Energy Rep. 2020, 6, 288–306. [Google Scholar] [CrossRef]
- Djinović, P.S. Electrochemical Energy Storage for Renewable Sources and Grid Balancing; Elsevier: Amsterdam, The Netherlands, 2015; pp. 183–199. [Google Scholar]
- Lan, R.; Irvine, J.T.S.; Tao, S.W. Ammonia and related chemicals as potential indirect hydrogen storage materials. Int. J. Hydrogen Energy 2012, 37, 1482–1494. [Google Scholar] [CrossRef]
- Wang, W.T.; Herreros, J.M.; Tsolakis, A.; York, A.P.E. Ammonia as hydrogen carrier for transportation; investigation of the ammonia exhaust gas fuel reforming. Int. J. Hydrogen Energy 2013, 38, 9907–9917. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, M.; Muroyama, H.; Suzuki, S.; Saito, M.; Koide, T.; Takahashi, Y.; Horiuchi, T.; Yamasaki, H.; Matsumoto, S.; Kubo, H.; et al. Development of 1 kW-class Ammonia-fueled Solid Oxide Fuel Cell Stack. Fuel Cells 2020, 20, 80–88. [Google Scholar] [CrossRef]
- Zhao, Y.; Setzler, B.P.; Wang, J.H.; Nash, J.; Wang, T.; Xu, B.J.; Yan, Y.S. An Efficient Direct Ammonia Fuel Cell for Affordable Carbon-Neutral Transportation. Joule 2019, 3, 2472–2484. [Google Scholar] [CrossRef]
- Schrock, R.R. Reduction of dinitrogen. Proc. Natl. Acad. Sci. USA 2006, 103, 17087. [Google Scholar] [CrossRef] [Green Version]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The Evolution and Future of Earth’s Nitrogen Cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Smil, V. Population-Growth and Nitrogen—An Exploration of a Critical Existential Link. Popul. Dev. Rev. 1991, 17, 569–601. [Google Scholar] [CrossRef]
- John Humphreys, R.L.; Tao, S. Development and Recent Progress on Ammonia Synthesis Catalysts for Haber–Bosch Process. Adv. Energy Sustain. Res. 2020, 2, 1–23. [Google Scholar]
- Ashida, Y.; Arashiba, K.; Tanaka, H.; Egi, A.; Nakajima, K.; Yoshizawa, K.; Nishibayashi, Y. Molybdenum-Catalyzed Ammonia Formation Using Simple Monodentate and Bidentate Phosphines as Auxiliary Ligands. Inorg. Chem. 2019, 58, 8927–8932. [Google Scholar] [CrossRef] [PubMed]
- Buscagan, T.M.; Rees, D.C. Rethinking the Nitrogenase Mechanism: Activating the Active Site. Joule 2019, 3, 2662–2678. [Google Scholar] [CrossRef]
- Ashida, Y.; Arashiba, K.; Nakajima, K.; Nishibayashi, Y. Molybdenum-catalysed ammonia production with samarium diiodide and alcohols or water. Nature 2019, 568, 536–540. [Google Scholar] [CrossRef]
- Reid, W.V.; Ali, M.K.; Field, C.B. The future of bioenergy. Glob. Chang. Biol. 2020, 26, 274–286. [Google Scholar] [CrossRef] [Green Version]
- Akhlaghi, N.; Najafpour-Darzi, G. A comprehensive review on biological hydrogen production. Int. J. Hydrogen Energy 2020, 45, 22492–22512. [Google Scholar] [CrossRef]
- Mohamad, N.L.; Kamal, S.M.M.; Mokhtar, M.N. Xylitol Biological Production: A Review of Recent Studies. Food Rev. Int. 2015, 31, 74–89. [Google Scholar] [CrossRef]
- Ma, H.Z.; Wang, Q.H.; Qian, D.Y.; Gong, L.J.; Zhang, W.Y. The utilization of acid-tolerant bacteria on ethanol Production from kitchen garbage. Renew. Energy 2009, 34, 1466–1470. [Google Scholar] [CrossRef]
- Mahidhara, G.; Burrow, H.; Sasikala, C.; Ramana, C.V. Biological hydrogen production: Molecular and electrolytic perspectives. World J. Microbiol. Biotechnol. 2019, 35, 8. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, S.; Herwig, C. A comprehensive and quantitative review of dark fermentative biohydrogen production. Microbiol. Cell Fact. 2012, 11, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, F.; Lago, A.; Rocha, V.; Barros, O.; Costa, L.; Vipotnik, Z.; Silva, B.; Tavares, T. A Review on Biological Processes for Pharmaceuticals Wastes Abatement-A Growing Threat to Modern Society. Environ. Sci. Technol. 2019, 53, 7185–7202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, M.S.G. Engineering biological systems toward a sustainable bioeconomy. J. Ind. Microbiol. Biotechnol. 2015, 42, 813–838. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A.; Dordick, J.S.; Hauer, B.; Kiener, A.; Wubbolts, M.; Witholt, B. Industrial biocatalysis today and tomorrow. Nature 2001, 409, 258–268. [Google Scholar] [CrossRef]
- Thomas, S.M.; DiCosimo, R.; Nagarajan, A. Biocatalysis: Applications and potentials for the chemical industry. Trends Biotechnol. 2002, 20, 444. [Google Scholar] [CrossRef]
- Ogawa, J.; Shimizu, S. Microbial enzymes: New industrial applications from traditional screening methods. Trends Biotechnol. 1999, 17, 13–21. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, N.; Kisker, C.; Sehlessman, J.L.; Howard, J.B.; Rees, D.C. Structure of ADP center dot AIF(4)(−)-stabilized nitrogenase complex and its implications for signal transduction. Nature 1997, 387, 370–376. [Google Scholar] [CrossRef]
- Rapson, T.D.; Gregg, C.M.; Allen, R.S.; Ju, H.; Doherty, C.M.; Mulet, X.; Giddey, S.; Wood, C.C. Insights into Nitrogenase Bioelectrocatalysis for Green Ammonia Production. Chemsuschem 2020, 13, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, K.; Mousavi, S.A. Effectiveness of nitrogen fixation in rhizobia. Microbiol. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293–296. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Snider, M.G.; Temple, B.S.; Wolfenden, R. The path to the transition state in enzyme reactions: A survey of catalytic efficiencies. J. Phys. Org. Chem. 2004, 17, 586–591. [Google Scholar] [CrossRef]
- Garcia, H.G.; Tikhonov, M.; Lin, A.; Gregor, T. Quantitative imaging of transcription in living Drosophila embryos links polymerase activity to patterning. Curr. Biol. CB 2013, 23, 2140–2145. [Google Scholar] [CrossRef] [Green Version]
- Ingolia, N.T.; Lareau, L.F.; Weissman, J.S. Ribosome profiling of mouse embryonic stem cells reveals the complexity and dynamics of mammalian proteomes. Cell 2011, 147, 789–802. [Google Scholar] [CrossRef] [Green Version]
- Einsle, O.; Rees, D.C. Structural Enzymology of Nitrogenase Enzymes. Chem. Rev. 2020, 120, 4969–5004. [Google Scholar] [CrossRef]
- Cai, R.; Minteer, S.D. Nitrogenase Bioelectrocatalysis: From Understanding Electron-Transfer Mechanisms to Energy Applications. ACS Energy Lett. 2018, 3, 2736–2742. [Google Scholar] [CrossRef]
- Milton, R.D.; Minteer, S.D. Nitrogenase Bioelectrochemistry for Synthesis Applications. Acc. Chem. Res. 2019, 52, 3351–3360. [Google Scholar] [CrossRef]
- Peters, J.W.; Szilagyi, R.K. Exploring new frontiers of nitrogenase structure and mechanism. Curr. Opin. Chem. Biol. 2006, 10, 101–108. [Google Scholar] [CrossRef]
- Hoffman, B.M.; Lukoyanov, D.; Yang, Z.Y.; Dean, D.R.; Seefeldt, L.C. Mechanism of nitrogen fixation by nitrogenase: The next stage. Chem. Rev. 2014, 114, 4041–4062. [Google Scholar] [CrossRef] [PubMed]
- Eady, R.R. Structure-function relationships of alternative nitrogenases. Chem. Rev. 1996, 96, 3013–3030. [Google Scholar] [CrossRef] [PubMed]
- Weininger, M.S.; Mortenson, L.E. Crystallographic Properties of the Mofe Proteins of Nitrogenase from Clostridium-Pasteurianum and Azotobacter-Vinelandii. Proc. Natl. Acad. Sci. USA 1982, 79, 378–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einsle, O.; Tezcan, F.A.; Andrade, S.L.A.; Schmid, B.; Yoshida, M.; Howard, J.B.; Rees, D.C. Nitrogenase MoFe-protein at 1.16 angstrom resolution: A central ligand in the FeMo-cofactor. Science 2002, 297, 1696–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.G.; Xie, X.Q.; Xiang, N.; Tian, Z.X.; Dixon, R.; Wang, P. Polyprotein strategy for stoichiometric assembly of nitrogen fixation components for synthetic biology. Proc. Natl. Acad. Sci. USA 2018, 115, E8509–E8517. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Y.; Zhang, L.H.; Liu, Z.Z.; Zhao, D.H.; Liu, X.M.; Zhang, B.; Xie, J.B.; Hong, Y.Y.; Li, P.F.; Chen, S.F.; et al. A Minimal Nitrogen Fixation Gene Cluster from Paenibacillus sp WLY78 Enables Expression of Active Nitrogenase in Escherichia coli. PLoS Genet. 2013, 9, e1003865. [Google Scholar] [CrossRef]
- Li, X.X.; Liu, Q.; Liu, X.M.; Shi, H.W.; Chen, S.F. Using synthetic biology to increase nitrogenase activity. Microbiol. Cell Fact. 2016, 15, 43. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Xie, X.; Wang, X.; Dixon, R.; Wang, Y.P. Reconstruction and minimal gene requirements for the alternative iron-only nitrogenase in Escherichia coli. Proc. Natl. Acad. Sci. USA 2014, 111, E3718–E3725. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Torrejon, G.; Jimenez-Vicente, E.; Buesa, J.M.; Hernandez, J.A.; Verma, H.K.; Rubio, L.M. Expression of a functional oxygen-labile nitrogenase component in the mitochondrial matrix of aerobically grown yeast. Nat. Commun. 2016, 7, 11426. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Torrejon, G.; Buren, S.; Veldhuizen, M.; Rubio, L.M. Biosynthesis of cofactor-activatable iron-only nitrogenase in Saccharomyces cerevisiae. Microbiol. Biotechnol. 2021, 14, 1073–1083. [Google Scholar] [CrossRef]
- Buren, S.; Jiang, X.; Lopez-Torrejon, G.; Echavarri-Erasun, C.; Rubio, L.M. Purification and In Vitro Activity of Mitochondria Targeted Nitrogenase Cofactor Maturase NifB. Front. Plant. Sci. 2017, 8, 1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buren, S.; Pratt, K.; Jiang, X.; Guo, Y.; Jimenez-Vicente, E.; Echavarri-Erasun, C.; Dean, D.R.; Saaem, I.; Gordon, D.B.; Voigt, C.A.; et al. Biosynthesis of the nitrogenase active-site cofactor precursor NifB-co in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2019, 116, 25078–25086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, S.; Gregg, C.M.; Allen, R.S.; Menon, A.; Hussain, D.; Gillespie, V.; Johnston, E.; Byrne, K.; Colgrave, M.L.; Wood, C.C. A Synthetic Biology Workflow Reveals Variation in Processing and Solubility of Nitrogenase Proteins Targeted to Plant Mitochondria, and Differing Tolerance of Targeting Sequences in a Bacterial Nitrogenase Assay. Front. Plant. Sci. 2020, 11, 552160. [Google Scholar] [CrossRef] [PubMed]
- Xiang, N.; Guo, C.Y.; Liu, J.W.; Xu, H.; Dixon, R.; Yang, J.G.; Wang, Y.P. Using synthetic biology to overcome barriers to stable expression of nitrogenase in eukaryotic organelles. Proc. Natl. Acad. Sci. USA 2020, 117, 24602. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.S.; Gregg, C.M.; Okada, S.; Menon, A.; Hussain, D.; Gillespie, V.; Johnston, E.; Devilla, R.; Warden, A.C.; Taylor, M.; et al. Plant expression of NifD protein variants resistant to mitochondrial degradation. Proc. Natl. Acad. Sci. USA 2020, 117, 23165–23173. [Google Scholar] [CrossRef]
- Chretien, D.; Benit, P.; Ha, H.H.; Keipert, S.; El-Khoury, R.; Chang, Y.T.; Jastroch, M.; Jacobs, H.T.; Rustin, P.; Rak, M. Mitochondria are physiologically maintained at close to 50 degrees C. PLoS Biol. 2018, 16, e2003992. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Chen, S. Transfer of Nitrogen Fixation (nif) Genes to Non-diazotrophic Hosts. Chembiochem 2020, 21, 1717–1722. [Google Scholar] [CrossRef]
- Dong, F.; Lee, Y.S.; Gaffney, E.M.; Grattieri, M.; Haddadin, H.; Minteer, S.D.; Chen, H. An engineered, non-diazotrophic cyanobacterium and its application in bioelectrochemical nitrogen fixation. Cell Rep. Phys. Sci. 2021, 2, 100444. [Google Scholar] [CrossRef]
- Mehta, M.P.; Baross, J.A. Nitrogen fixation at 92 degrees C by a hydrothermal vent archaeon. Science 2006, 314, 1783–1786. [Google Scholar] [CrossRef]
- DiCosimo, R.; McAuliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Cai, R.; Milton, R.; Chen, H.; Minteer, S.D. Pyrene-Based Noncovalent Immobilization of Nitrogenase on Carbon Surfaces. Chembiochem 2020, 21, 1729–1732. [Google Scholar] [CrossRef] [PubMed]
- Schlesier, J.; Rohde, M.; Gerhardt, S.; Einsle, O. A Conformational Switch Triggers Nitrogenase Protection from Oxygen Damage by Shethna Protein II (FeSII). J. Am. Chem. Soc. 2016, 138, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xie, X.; Yang, M.; Dixon, R.; Wang, Y.P. Modular electron-transport chains from eukaryotic organelles function to support nitrogenase activity. Proc. Natl. Acad. Sci. USA 2017, 114, E2460–E2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheatley, R.M.; Ford, B.L.; Li, L.; Aroney, S.T.N.; Knights, H.E.; Ledermann, R.; East, A.K.; Ramachandran, V.K.; Poole, P.S. Lifestyle adaptations of Rhizobium from rhizosphere to symbiosis. Proc. Natl. Acad. Sci. USA 2020, 117, 23823–23834. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Rubio, M.C.; Xin, X.; Zhang, B.L.; Fan, Q.L.; Wang, Q.; Ning, G.G.; Becana, M.; Duanmu, D.Q. CRISPR/Cas9 knockout of leghemoglobin genes in Lotus japonicus uncovers their synergistic roles in symbiotic nitrogen fixation. New Phytol. 2019, 224, 818–832. [Google Scholar] [CrossRef]
- Nonaka, A.; Yamamoto, H.; Kamiya, N.; Kotani, H.; Yamakawa, H.; Tsujimoto, R.; Fujita, Y. Accessory Proteins of the Nitrogenase Assembly, NifW, NifX/NafY, and NifZ, Are Essential for Diazotrophic Growth in the Nonheterocystous Cyanobacterium Leptolyngbya boryana. Front. Microbiol. 2019, 10, 495. [Google Scholar] [CrossRef] [Green Version]
- Karsten, T.; Alvin, T.; Sarah, B.; Rosemary, C.; Emily, T.; Kevin, H.; Douglas, H.; Austin, D. Methods and Compositions for Improving Plant. Traits. Patent WO2017011602A1, 19 January 2017. [Google Scholar]
- Pivot Bio. Pivot Bio 2020 Performance Report; Pivot Bio: Berkeley, CA, USA, 2021. [Google Scholar]
- FAO. Towards the Future we Want: End Hunger and Make the Transition to Sustainable Agricultural and Food Systems; FAO: Rome, Italy, 2012. [Google Scholar]
- Melikoglu, M.; Lin, C.S.K.; Webb, C. Analysing global food waste problem: Pinpointing the facts and estimating the energy content. Open Eng. 2013, 3, 157–164. [Google Scholar] [CrossRef]
- Van Dyk, J.S.; Gama, R.; Morrison, D.; Swart, S.; Pletschke, B.I. Food processing waste: Problems, current management and prospects for utilisation of the lignocellulose component through enzyme synergistic degradation. Renew. Sustain. Energy Rev. 2013, 26, 521–531. [Google Scholar] [CrossRef]
- Nasir, I.M.; Ghazi, T.I.M.; Omar, R. Production of biogas from solid organic wastes through anaerobic digestion: A review. Appl. Microbiol. Biotechnol. 2012, 95, 321–329. [Google Scholar] [CrossRef]
- Morita, M.; Sasaki, K. Factors influencing the degradation of garbage in methanogenic bioreactors and impacts on biogas formation. Appl. Microbiol. Biotechnol. 2012, 94, 575–582. [Google Scholar] [CrossRef]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar] [CrossRef] [Green Version]
- Griffin, M.E.; McMahon, K.D.; Mackie, R.I.; Raskin, L. Methanogenic population dynamics during start-up of anaerobic digesters treating municipal solid waste and biosolids. Biotechnol. Bioeng. 1998, 57, 342–355. [Google Scholar] [CrossRef]
- Yang, X.; Lee, S.J.; Yoo, H.Y.; Choi, H.S.; Park, C.; Kim, S.W. Biorefinery of instant noodle waste to biofuels. Bioresour. Technol. 2014, 159, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Si, B.C.; Li, J.M.; Zhu, Z.B.; Zhang, Y.H.; Lu, J.W.; Shen, R.X.; Zhang, C.; Xing, X.H.; Liu, Z. Continuous production of biohythane from hydrothermal liquefied cornstalk biomass via two-stage high-rate anaerobic reactors. Biotechnol. Biofuels 2016, 9, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, A.D.; Joshi, T.R.; Srivastava, S.K.; Darani, K.K.; Khade, S.; Srivastava, J. Effect of nutritional supplements on bio-plastics (PHB) production utilizing sugar refinery waste with potential application in food packaging. Prep. Biochem. Biotechnol. 2019, 49, 567–577. [Google Scholar] [CrossRef]
- Vong, W.C.; Au Yang, K.L.C.; Liu, S.Q. Okara (soybean residue) biotransformation by yeast Yarrowia lipolytica. Int. J. Food Microbiol. 2016, 235, 1–9. [Google Scholar] [CrossRef]
- Liu, C.; Hotta, Y.; Santo, A.; Hengesbaugh, M.; Watabe, A.; Totoki, Y.; Allen, D.; Bengtsson, M. Food waste in Japan: Trends, current practices and key challenges. J. Clean Prod. 2016, 133, 557–564. [Google Scholar] [CrossRef]
- Choi, K.Y.; Wernick, D.G.; Tat, C.A.; Liao, J.C. Consolidated conversion of protein waste into biofuels and ammonia using Bacillus subtilis. Metab. Eng. 2014, 23, 53–61. [Google Scholar] [CrossRef]
- Huo, Y.X.; Cho, K.M.; Rivera, J.G.L.; Monte, E.; Shen, C.R.; Yan, Y.J.; Liao, J.C. Conversion of proteins into biofuels by engineering nitrogen flux. Nat. Biotechnol. 2011, 29, 346-U160. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kuroda, K.; Tatemichi, Y.; Nakahara, T.; Aoki, W.; Ueda, M. Construction of engineered yeast producing ammonia from glutamine and soybean residues (okara). AMB Express 2020, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Aoki, W.; Ueda, M. Improved ammonia production from soybean residues by cell surface-displayed l-amino acid oxidase on yeast. Biosci. Biotechnol. Biochem. 2021, 85, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Tatemichi, Y.; Kuroda, K.; Nakahara, T.; Ueda, M. Efficient ammonia production from food by-products by engineered Escherichia coli. AMB Express 2020, 10, 150. [Google Scholar] [CrossRef] [PubMed]
- Katami, T.; Yasuhara, A.; Shibamoto, T. Formation of dioxins from incineration of foods found in domestic garbage. Environ. Sci. Technol. 2004, 38, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.C.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [Green Version]
- Montzka, S.A.; Dlugokencky, E.J.; Butler, J.H. Non-CO2 greenhouse gases and climate change. Nature 2011, 476, 43–50. [Google Scholar] [CrossRef]
- Chan, L.Y.; Takahashi, M.; Lim, P.J.; Aoyama, S.; Makino, S.; Ferdinandus, F.; Ng, S.C.; Arai, S.; Fujita, H.; Tan, H.C.; et al. Eurotium Cristatum Fermented Okara as a Potential Food Ingredient to Combat Diabetes. Sci. Rep. 2019, 9, 17536. [Google Scholar] [CrossRef]
- Hadj Saadoun, J.; Calani, L.; Cirlini, M.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. Effect of fermentation with single and co-culture of lactic acid bacteria on okara: Evaluation of bioactive compounds and volatile profiles. Food Funct. 2021, 12, 3033–3043. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Yoneda, H.; Tatsukami, Y.; Aoki, W.; Ueda, M. Ammonia production from amino acid-based biomass-like sources by engineered Escherichia coli. AMB Express 2017, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Tanaka, A. Cell surface engineering of yeast: Construction of arming yeast with biocatalyst. J. Biosci. Bioeng. 2000, 90, 125–136. [Google Scholar] [CrossRef]
- Kuroda, K.; Ueda, M. Cell surface engineering of yeast for applications in white biotechnology. Biotechnol. Lett. 2011, 33, 1–9. [Google Scholar] [CrossRef]
- Ueda, M. Establishment of cell surface engineering and its development. Biosci. Biotechnol. Biochem. 2016, 80, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, A.; Kuroda, K.; Ueda, M. Direct fermentation of newspaper after laccase-treatment using yeast codisplaying endoglucanase, cellobiohydrolase, and beta-glucosidase. Renew. Energy 2012, 44, 199–205. [Google Scholar] [CrossRef]
- Takagi, T.; Yokoi, T.; Shibata, T.; Morisaka, H.; Kuroda, K.; Ueda, M. Engineered yeast whole-cell biocatalyst for direct degradation of alginate from macroalgae and production of non-commercialized useful monosaccharide from alginate. Appl. Microbiol. Biotechnol. 2016, 100, 1723–1732. [Google Scholar] [CrossRef]
- Motone, K.; Takagi, T.; Sasaki, Y.; Kuroda, K.; Ueda, M. Direct ethanol fermentation of the algal storage polysaccharide laminarin with an optimized combination of engineered yeasts. J. Biotechnol. 2016, 231, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.; Sousa, M.J.; Leao, C. Ammonium is toxic for aging yeast cells, inducing death and shortening of the chronological lifespan. PLoS ONE 2012, 7, e37090. [Google Scholar] [CrossRef] [Green Version]
- Pollegioni, L.; Motta, P.; Molla, G. l-Amino acid oxidase as biocatalyst: A dream too far? Appl. Microbiol. Biotechnol. 2013, 97, 9323–9341. [Google Scholar] [CrossRef]
- Lu, P.L.; Ma, D.; Chen, Y.L.; Guo, Y.Y.; Chen, G.Q.; Deng, H.T.; Shi, Y.G. l-glutamine provides acid resistance for Escherichia coli through enzymatic release of ammonia. Cell Res. 2013, 23, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Bloess, S.; Beuel, T.; Kruger, T.; Sewald, N.; Dierks, T.; Fischer von Mollard, G. Expression, characterization, and site-specific covalent immobilization of an l-amino acid oxidase from the fungus Hebeloma cylindrosporum. Appl. Microbiol. Biotechnol. 2019, 103, 2229–2241. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Johnson, P.M.; Ko, K.C.; Kamio, M.; Germann, M.W.; Derby, C.D.; Tai, P.C. Cloning, characterization and expression of escapin, a broadly antimicrobial FAD-containing l-amino acid oxidase from ink of the sea hare Aplysia californica. J. Exp. Biol. 2005, 208, 3609–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, K.C.; Wang, B.; Tai, P.C.; Derby, C.D. Identification of potent bactericidal compounds produced by escapin, an L-amino acid oxidase in the ink of the sea hare Aplysia californica. Antimicrob. Agents Chemother. 2008, 52, 4455–4462. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A. Structure-Function Studies and Mechanism of Action of Snake Venom l-Amino Acid Oxidases. Front. Pharm. 2020, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Hill, A.K.; Torrente-Murciano, L. Current and future role of Haber-Bosch ammonia in a carbon-free energy landscape. Energy Environ. Sci. 2020, 13, 331–344. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Microorganisms | Methods | Resource | References |
|---|---|---|---|
| Bacillus subtilis | Metabolic engineering | Amino acids-based medium | [102] |
| Escherichia coli | Metabolic engineering | Amino acids-based medium | [112] |
| Escherichia coli | Metabolic engineering | Amino acids-based medium and soybean residue | [106] |
| Saccharomyces cerevisiae | Cell surface engineering | Glutamine solution and soybean residue | [104] |
| Saccharomyces cerevisiae | Cell surface engineering | Amino acids solution and soybean residue | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, Y.; Aoki, W.; Ueda, M. Sustainable Biological Ammonia Production towards a Carbon-Free Society. Sustainability 2021, 13, 9496. https://doi.org/10.3390/su13179496
Watanabe Y, Aoki W, Ueda M. Sustainable Biological Ammonia Production towards a Carbon-Free Society. Sustainability. 2021; 13(17):9496. https://doi.org/10.3390/su13179496
Chicago/Turabian StyleWatanabe, Yukio, Wataru Aoki, and Mitsuyoshi Ueda. 2021. "Sustainable Biological Ammonia Production towards a Carbon-Free Society" Sustainability 13, no. 17: 9496. https://doi.org/10.3390/su13179496