Sustainable Second-Generation Bioethanol Production from Enzymatically Hydrolyzed Domestic Food Waste Using Pichia anomala as Biocatalyst




Abstract
:1. Introduction
2. Materials and Methods
2.1. Food Waste
2.2. Enzymatic Saccharification
2.3. Bioethanol Production
2.3.1. Preparation of Inoculum
2.3.2. Fermentation Experiments
2.4. Analytical Methods
2.5. Calculations
2.6. Statistical Analysis of Data
2.7. Kinetics Expressions and Simulation of Data
3. Results and Discussion
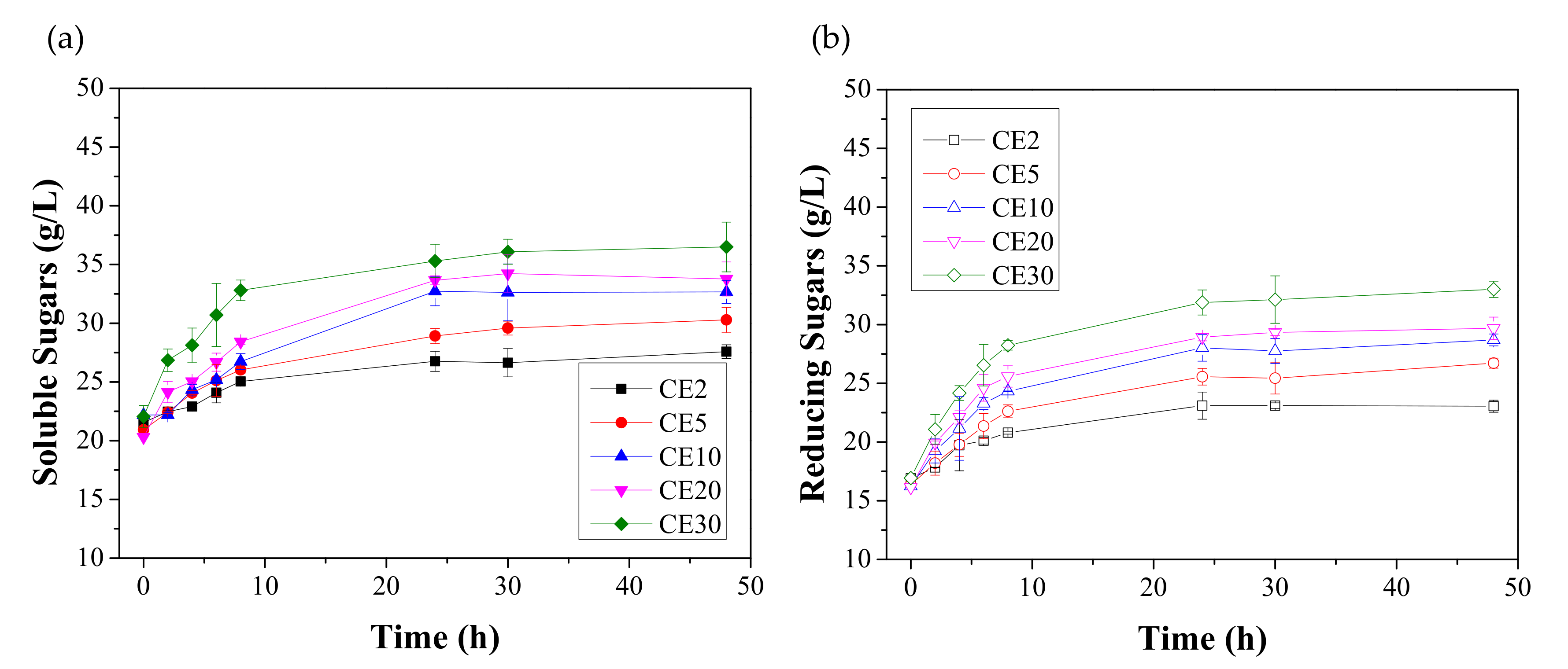
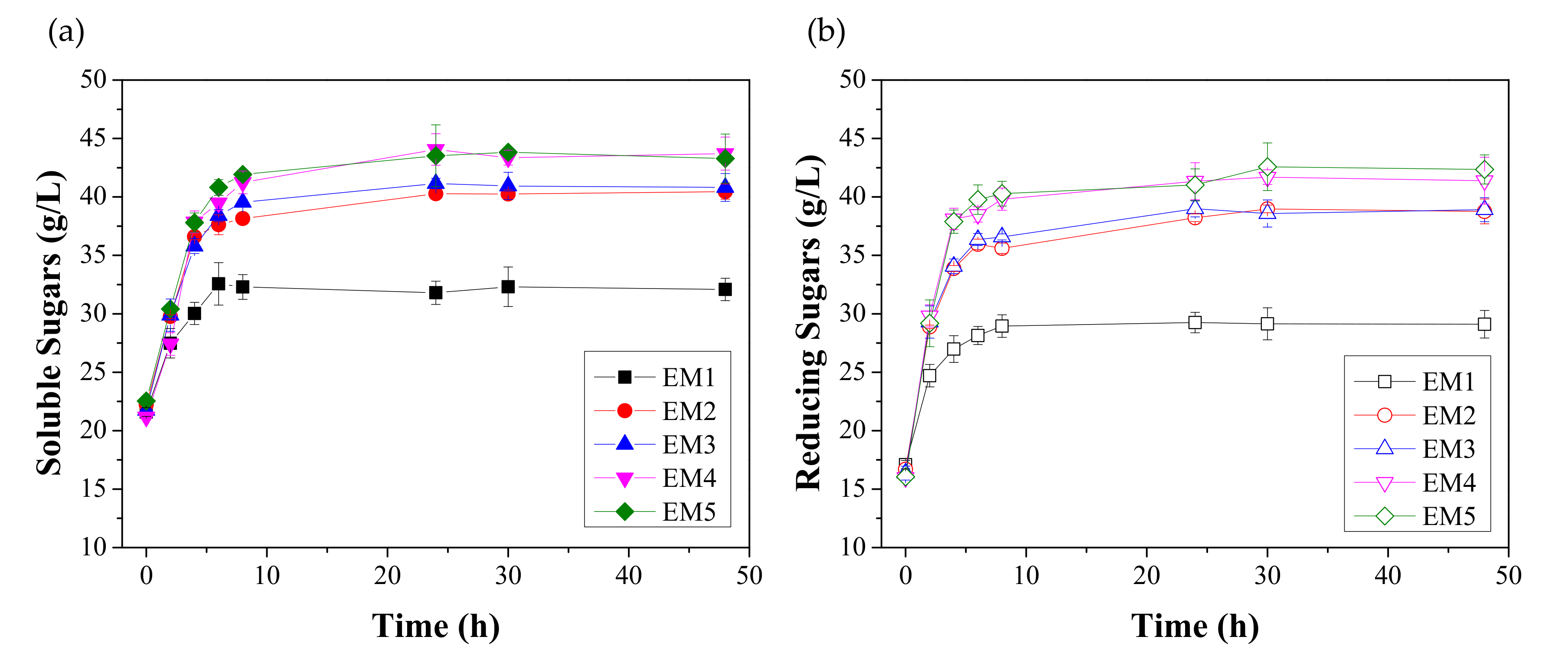
3.1. Effect of Enzymatic Hydrolysis on the Saccharification of FORBI
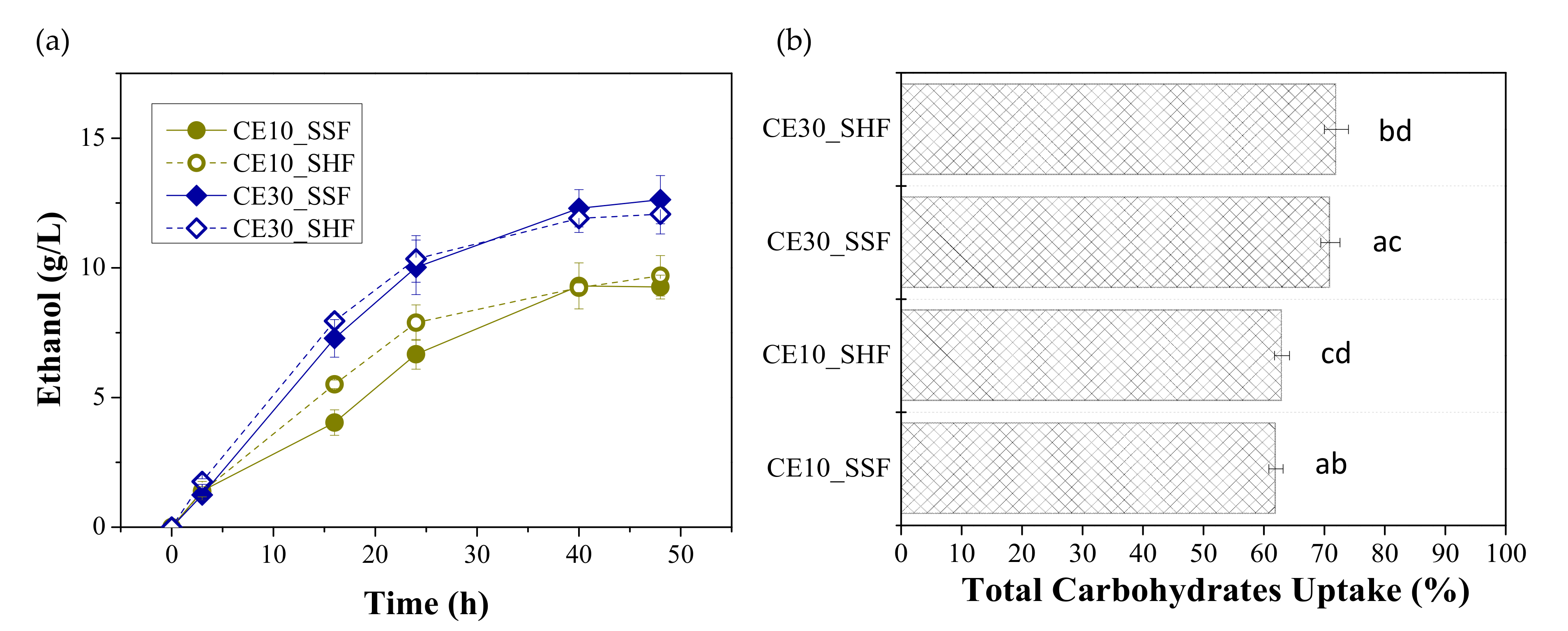
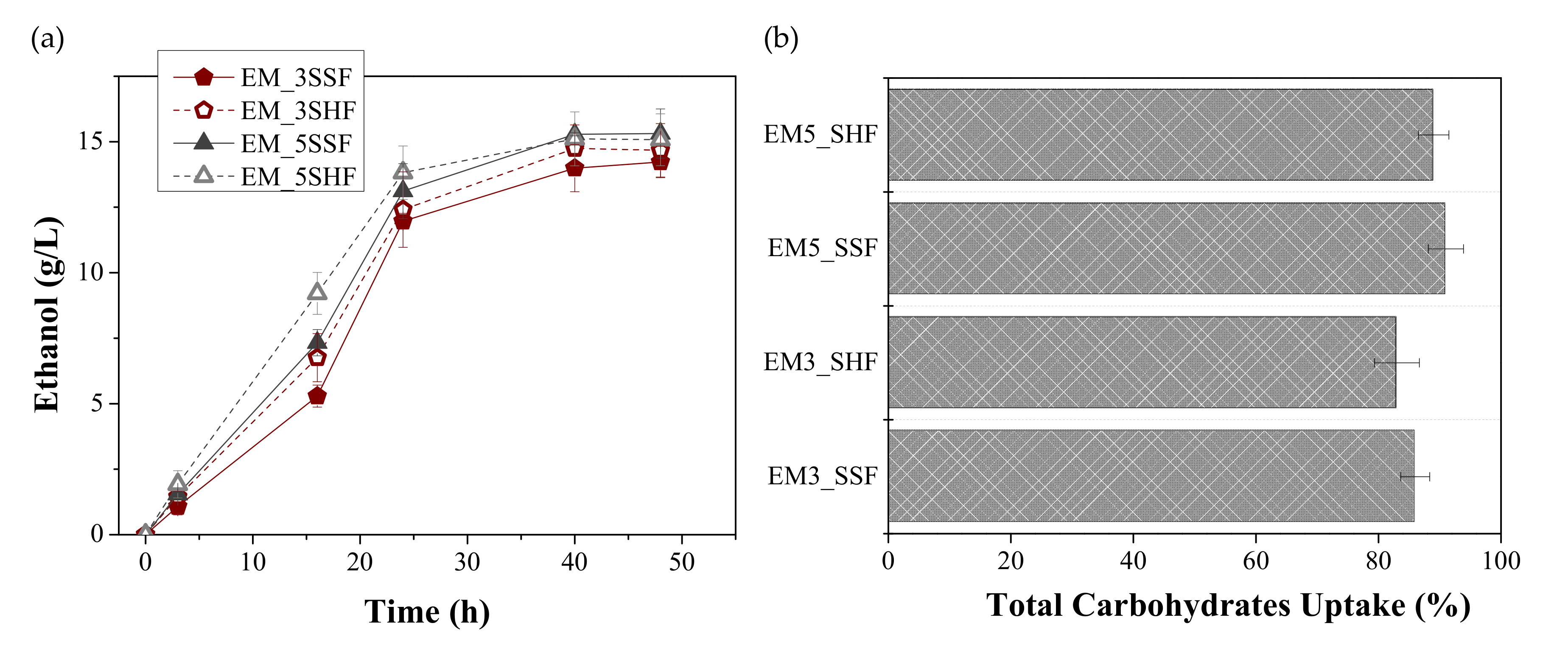
3.2. Effect of Enzymatic Loading and Fermentation Mode on Ethanol Production
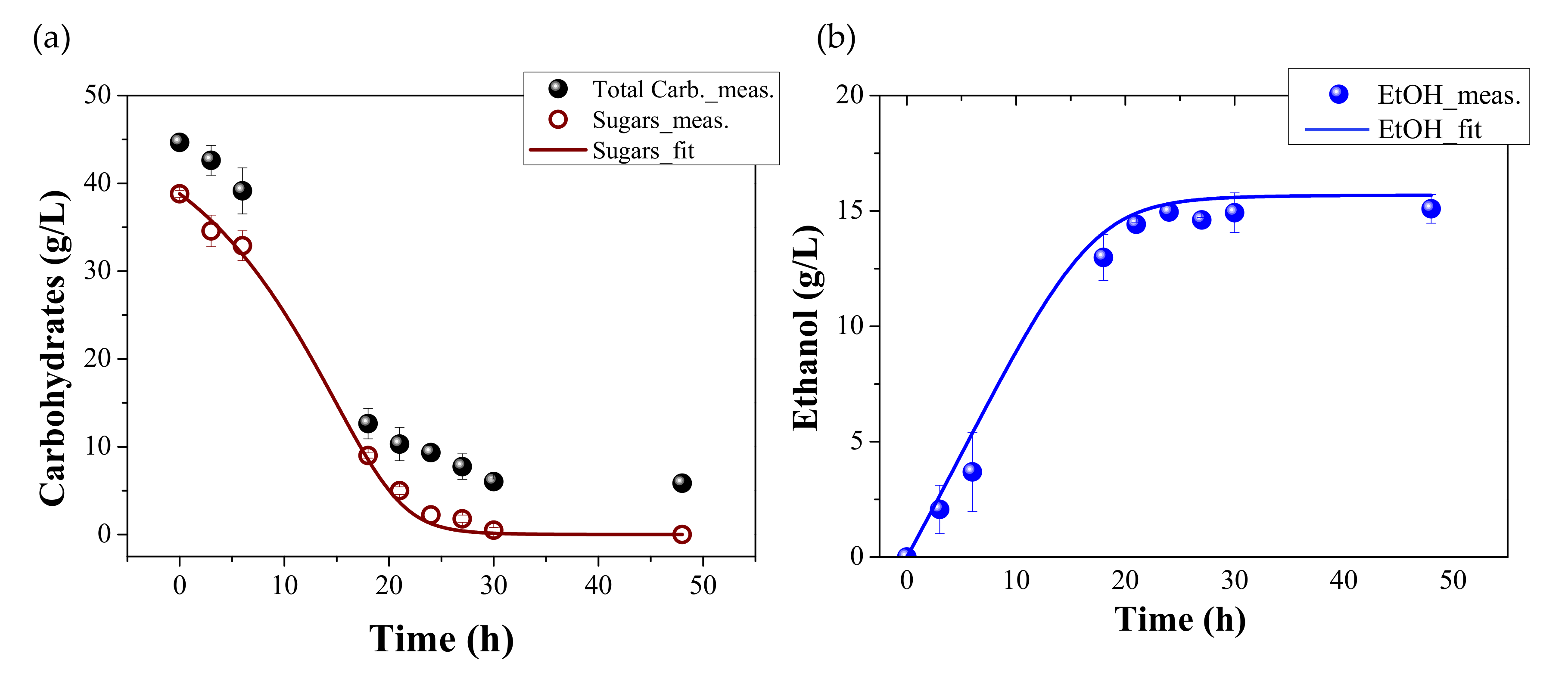
3.3. Verification of Ethanol Production Kinetics of P. anomala during Fermentation of Hydrolysed FORBI
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teoh, Y.H.; Yu, K.H.; How, H.G.; Nguyen, H.-T. Experimental investigation of performance, emission and combustion characteristics of a common-rail diesel engine fuelled with bioethanol as a fuel additive in coconut oil biodiesel blends. Energies 2019, 12, 1954. [Google Scholar] [CrossRef] [Green Version]
- Sarris, D.; Papanikolaou, S. Biotechnological production of ethanol: Biochemistry, processes and technologies. Eng. Life Sci. 2016, 16, 307–329. [Google Scholar] [CrossRef] [Green Version]
- FAO. 2011 Global Food Losses and Food Waste—Extent, Causes and Prevention. Rome. Available online: http://www.fao.org/3/mb060e/mb060e00.htm (accessed on 15 November 2020).
- United Nations, Sustainable Development Goals. Available online: https://sustainabledevelopment.un.org/sdg12 (accessed on 15 November 2020).
- Uckun Kiran, E.; Liu, Y. Bioethanol production from mixed food waste by an effective enzymatic pretreatment. Fuel 2015, 159, 463–469. [Google Scholar] [CrossRef]
- Aruna, A.; Nagavalli, M.; Girijashankar, V.; Ponamgi, S.P.D.; Swathisree, V.; Venkateswar Rao, L. Direct bioethanol production by amylolytic yeast Candida albicans. Lett. Appl. Microbiol. 2015, 60, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.T.; Praveen, P.; Loh, K.-C. Zymomonas mobilis immobilization in polymeric membranes for improved resistance to lignocellulose-derived inhibitors in bioethanol fermentation. Biochem. Eng. J. 2018, 140, 29–37. [Google Scholar] [CrossRef]
- Sowatad, A.; Todhanakasem, T. Bioethanol production by repeated batch using immobilized yeast cells on sugarcane bagasse. Waste Biomass Valor. 2020, 11, 2009–2016. [Google Scholar] [CrossRef]
- Selim, K.A.; Easa, S.M.; El-Diwany, A.I. The xylose metabolizing yeast Spathaspora passalidarum is a promising genetic treasure for improving bioethanol production. Fermentation 2020, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Menon, V.; Divate, R.; Rao, M. Bioethanol production from renewable polymer lichenan using lichenase from an alkalothermophilic Thermomonospora sp. and thermotolerant yeast. Fuel Process Technol. 2011, 92, 401–406. [Google Scholar] [CrossRef]
- Kumari, R.; Pramanik, K. Improved bioethanol production using fusants of Saccharomyces cerevisiae and xylose-fermenting yeasts. Appl. Biochem. Biotech. 2012, 167, 873–884. [Google Scholar] [CrossRef]
- Swain, M.R.; Mishra, J.; Thatoi, H. Bioethanol production from Sweet Potato (Ipomoea batatas L.) Flour using Co-Culture of Trichoderma sp. and Saccharomyces cerevisiae in solid-state fermentation. Braz. Arch. Biol. Techn. 2013, 56, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Izmirlioglu, G.; Demirci, A. Improved simultaneous saccharification and fermentation of bioethanol from industrial potato waste with co-cultures of Aspergillus niger and Saccharomyces cerevisiae by medium optimization. Fuel 2016, 185, 684–691. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Wang, L.; Yang, R.; Hou, P.; Liu, J. Direct bioethanol production from wheat straw using xylose/glucose co-fermentation by co-culture of two recombinant yeasts. J. Ind. Microbiol. Biot. 2017, 44, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Farias, D.; Maugeri Filho, F. Co-culture strategy for improved 2G bioethanol production using a mixture of sugarcane molasses and bagasse hydrolysate as substrate. Biochem. Eng. J. 2019, 147, 29–38. [Google Scholar] [CrossRef]
- Senkevich, S.; Ntaikou, I.; Lyberatos, G. Bioethanol production from thermochemically pre-treated olive mill solid residues using the yeast Pachysolen tannophylus. Glob. Nest. J. 2012, 14, 118–124. [Google Scholar]
- Martinez-Patino, J.C.; Romero-Garcia, J.M.; Ruiz, E.; Oliva, J.M.; Alvarez, C.; Romero, I.; Negro, M.J.; Castro, E. High solids loading pretreatment of olive tree pruning with dilute phosphoric acid for bioethanol production by Escherichia coli. Energy Fuels 2015, 29, 1735–1742. [Google Scholar] [CrossRef]
- Ben Atitallah, I.; Antonopoulou, G.; Ntaikou, I.; Alexandropoulou, M.; Nasri, M.; Mechichi, T.; Lyberatos, G. On the evaluation of different saccharification schemes for enhanced bioethanol production from potato peels waste via a newly isolated yeast strain of Wickerhamomyces anomalus. Biores. Technol. 2019, 289, 121614. [Google Scholar] [CrossRef]
- Lee, I.; Yu, J.-H. The production of fermentable sugar and bioethanol from acacia wood by optimizing dilute sulfuric acid pretreatment and post treatment. Fuel 2020, 275, 117943. [Google Scholar] [CrossRef]
- Ntaikou, I.; Siankiavich, S.; Lyberatos, G. Effect of thermo-chemical pretreatment on the saccharification and enzymatic digestibility of olive mill stones and their bioconversion towards alcohols. Environ. Sci. Pollut. Res. 2020, in press. [Google Scholar] [CrossRef]
- Alexandropoulou, M.; Antonopoulou, G.; Ntaikou, I.; Lyberatos, G. Fungal pretreatment of willow sawdust with Abortiporus biennis for anaerobic digestion: Impact of an external nitrogen source. Sustainability 2017, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Antonopoulou, G.; Kampranis, A.; Ntaikou, I.; Lyberatos, G. Enhancement of liquid and gaseous biofuels production from agro-industrial residues after thermochemical and enzymatic pretreatment. Front. Sustain. Food Syst. 2019, 3, 92. [Google Scholar] [CrossRef]
- Ntaikou, I.; Antonopoulou, G.; Vayenas, D.; Lyberatos, G. Assessment of electro.coagulation as a pretreatment method of olive mill wastewater towards alternative processes for biofuels production. Renew. Energy 2020, 154, 1252–1262. [Google Scholar] [CrossRef]
- Dimitrellos, G.; Lyberatos, G.; Antonopoulou, G. Does acid addition improve liquid hot water pretreatment of lignocellulosic biomass towards biohydrogen and biogas production? Sustainability 2020, 12, 8935. [Google Scholar] [CrossRef]
- Antonopoulou, G. Designing efficient processes for sustainable bioethanol and bio-hydrogen production from grass lawn waste. Molecules 2020, 25, 2889. [Google Scholar] [CrossRef] [PubMed]
- Silvennoinen, K.; Katajajuuri, J.-M.; Hartikainen, H.; Heikkila, L.; Reinikainen, A. Food waste volume and composition in Finnish households. Br. Food J. 2014, 116, 1058–1068. [Google Scholar] [CrossRef]
- Angelo, A.C.M.; Saraiva, A.B.; Climaco, J.C.N.; Infante, C.E.; Valle, R. Life cycle assessment and multi-criteria decision analysis: Selection of a strategy for domestic food waste management in Rio de Janeiro. J. Clean. Prod. 2017, 143, 744–756. [Google Scholar] [CrossRef]
- Di Bitonto, L.; Antonopoulou, G.; Braguglia, C.; Campanale, C.; Gallipoli, A.; Lyberatos, G.; Ntaikou, I.; Pastore, C. Lewis-BrØnsted acid catalysed ethanolysis of the organic fraction of municipal solid waste for efficient production of biofuels. Biores. Technol. 2018, 266, 297–305. [Google Scholar] [CrossRef]
- Ahmad, M.; Gani, A.; Hassan, I.; Huang, Q.; Shabbir, H. Production and characterization of starch nanoparticles by mild alkali hydrolysis and ultra-sonication process. Sci. Rep. 2020, 10, 3533. [Google Scholar] [CrossRef]
- Ulbrich, M.; Bai, Y.; Floter, E. The supporting effect of ultrasound on the acid hydrolysis of granular potato starch. Carbohydr. Polym. 2020, 230, 115633. [Google Scholar] [CrossRef]
- Zulnazri; Roesyadi, A.; Sumarno. Effects of hydrolysis conditions on the crystallinity, chemical structure, morphology, and thermal stability of cellulose nanocrystals extracted from oil palm biomass residue. Int. J. Chem. Tech. Res. 2016, 9, 456–464. [Google Scholar]
- Hashem, M.; Asseri, T.Y.A.; Alamri, S.A.; Alrumman, S.A. Feasibility and sustainability of bioethanol production from starchy restaurants’ bio-wastes by new yeast strains. Waste Biomass Valor. 2019, 10, 1617–1626. [Google Scholar] [CrossRef]
- Matsakas, L.; Christakopoulos, P. Ethanol production from enzymatically treated dried food waste using enzymes produced on-site. Sustainability 2015, 7, 1446–1458. [Google Scholar] [CrossRef] [Green Version]
- Tao, N.; Gao, Y.; Liu, Y. Isolation and characterization of a Pichia anomala strain: A promising candidate for bioethanol production. Braz. J. Microbiol. 2011, 42, 668–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, Y.; Hossain, A.H.; Tobola, F.; Sedee, N.; Havekes, M.; Punt, P.J. Pichia anomala 29X: A resistant strain for lignocellulosic biomass hydrolysate fermentation. FEMS Yeast Res. 2013, 13, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Atitallah, I.; Ntaikou, I.; Antonopoulou, G.; Alexandropoulou, M.; Brysch-Herzberg, M.; Nasri, M.; Lyberatos, G.; Mechichi, T. Evaluation of the non-conventional yeast strain Wickerhamomyces anomalus (Pichia anomala) X19 for enhanced bioethanol production using date palm sap as renewable feedstock. Renew. Energ. 2020, 154, 71–81. [Google Scholar] [CrossRef]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. Microbiological and biochemical performances of six yeast species as potential starter cultures for wet fermentation of coffee beans. LWT—Food Sci. Technol. 2020, 110430, in press. [Google Scholar] [CrossRef]
- Papanikola, K.; Papadopoulou, K.; Tsiliyannis, C.; Fotinopoulou, I.; Katsiampoulas, A.; Chalarakis, E.; Georgiopoulou, M.; Rontogianni, V.; Michalopoulos, I.; Mathioudakis, D.; et al. Food residue biomass product as an alternative fuel for the cement industry. Environ. Sci. Pollut. Res. 2019, 26, 35555–35564. [Google Scholar] [CrossRef]
- Ntaikou, I.; Menis, N.; Alexandropoulou, M.; Antonopoulou, G.; Lyberatos, G. Valorization of kitchen biowaste for ethanol production via simultaneous saccharification and fermentation using co-cultures of the yeasts Saccharomyces cerevisiae and Pichia stipitis. Biores. Technol. 2018, 263, 75–83. [Google Scholar] [CrossRef]
- APHA; AWWA; WPCF. Standard Methods for the Examination of Water and Wastewater; Franson, M.A., Ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Monlau, F.; Barakat, A.; Steyer, J.P.; Carrere, H. Comparison of seven types of thermo-chemical pretreatments on the structural features and anaerobic digestion of sunflower stalks. Bioresour. Technol. 2012, 120, 241–247. [Google Scholar] [CrossRef]
- VELP SCIENTIFICA. SER 148 Extraction Apparatus for the Quantitative Separation of a Substance from a Mixture by the Use of an Organic Solvent; Operation Manual. VELP Solvent Extraction Operation Manual; Velp Scientifica: Usmate, Italy, 2017; Volume 10001364/A11, p. 45. [Google Scholar]
- DuBois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylique acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass; Laboratory Analytical Procedure, NREL/TP-510-42618; National Renewable Energy Laboratory: Denver, CO, USA, 2008. [Google Scholar]
- Reichert, P. AQUASIM 2.0, Computer Program for the Identification and Simulation of Aquatic Systems; EAWAG: Dubendorf, Switzerland, 1998. [Google Scholar]
- Ye, Z.; Berson, R.E. Kinetic modeling of cellulose hydrolysis with first order inactivation of adsorbed cellulase. Biores. Technol. 2011, 102, 11194–11199. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.T.; Lee, Y.-H.; Beardmore, D.R. The influence of major structural features of cellulose on rate of enzymatic hydrolysis. Biotechnol. Bioeng. 1981, 23, 419–424. [Google Scholar] [CrossRef]
- Szymanska-Chargot, M.; Chylinska, M.; Gdula, K.; Koziol, A.; Zdunek, A. Isolation and characterization of cellulose from different fruit and vegetable pomaces. Polymers 2017, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, U.P.; Reiner, R.R.; Ralph, S.A. Estimation of cellulose crystallinity of lignocelluloses using near-IR FT-Raman spectroscopy and comparison of the Raman and Segal-WAXS methods. J. Agric. Food Chem. 2013, 61, 103–113. [Google Scholar] [CrossRef]
- Al-Zuhair, S. The effect of crystallinity of cellulose on the rate of reducing sugars production by heterogeneous enzymatic hydrolysis. Biores. Technol. 2008, 99, 4078–4085. [Google Scholar] [CrossRef]
- Dekker, R.F. Enzymatic hydrolysis of plant polysaccharides: Substrates for fermentation. Braz. J. Med. Biol. Res. 1989, 22, 1441–1456. [Google Scholar]
- Matsakas, L.; Kekos, D.; Loizidou, M.; Christakopoulos, P. Utilization of household food waste for the production of ethanol at high dry material content. Biotechnol. Biofuels 2014, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.C.; Song, I.S.; Kim, J.C.; Shirai, Y.; Lee, D.H.; Kim, J.K.; Chung, S.O.; Kim, D.H.; Oh, K.K.; Cho, Y.S. Enzymatic hydrolysis of food waste and ethanol fermentation. Int. J. Energy Res. 2009, 3, 164–172. [Google Scholar] [CrossRef]
- Salimi, E.; Saragas, K.; Taheri, M.E.; Novakovic, J.; Barampouti, E.M.; Mai, S.; Moustakas, K.; Malamis, D.; Loizidou, M. The Role of Enzyme Loading on Starch and Cellulose Hydrolysis of Food Waste. Waste Biomass Valor. 2019, 10, 3753–3762. [Google Scholar] [CrossRef]
- Pereira, C.R.; Resende, J.T.V.; Guerra, E.P.; Lima, V.A.; Martins, M.D.; Knob, A. Enzymatic conversion of sweet potato granular starch into fermentable sugars: Feasibility of sweet potato peel as alternative substrate for α-amylase production. Biocat. Agric. Biotech. 2017, 11, 231–238. [Google Scholar] [CrossRef]
- Bothast, R.J.; Schlicher, M.A. Biotechnological processes for conversion of corn into ethanol. Appl. Microbiol. Biotech. 2005, 67, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Li, P.; Gong, X.; Wang, J. A comparison of the production of ethanol between simultaneous saccharification and fermentation and separate hydrolysis and fermentation using unpretreated cassava pulp and enzyme cocktail. Biosci. Biotechnol. Biochem. 2012, 76, 671–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahnum, D.; Tasum, S.O.; Triwahyuni, E.; Nurdin, M.; Abimanyu, H. Comparison of SHF and SSF processes using enzyme and dry yeast for optimization of bioethanol production from empty fruit bunch. Energy Procedia 2015, 68, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.J. Factors affecting efficiency of biomass fermentation to ethanol. Biotechnol. Bioeng. Symp. 1986, 17, 617–627. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Code | Cellulase Blend | Fungal Amylase | Amyloglucosidase | |||
|---|---|---|---|---|---|---|---|
| FPU/g TS | FPU/g Cellulose | U/g TS | U/g Starch | U/g TS | U/g Starch | ||
| Cellulolytic | CE2 | 2 | 18 | 0 | 0 | 0 | 0 |
| CE5 | 5 | 44 | 0 | 0 | 0 | 0 | |
| CE10 | 10 | 88 | 0 | 0 | 0 | 0 | |
| CE20 | 20 | 176 | 0 | 0 | 0 | 0 | |
| CE30 | 30 | 265 | 0 | 0 | 0 | 0 | |
| Cellulolytic and Amylolytic | EM1 | 0 | 0 | 10 | 61 | 5 | 31 |
| EM2 | 30 | 265 | 2 | 12 | 0 | 0 | |
| EM3 | 30 | 265 | 2 | 12 | 1 | 6 | |
| EM4 | 30 | 265 | 10 | 61 | 0 | 0 | |
| EM5 | 30 | 265 | 10 | 61 | 5 | 31 | |
| Enzymes | Reducing/Soluble Sugars (%) | SE (%) |
|---|---|---|
| CE2 | 83.55 ± 0.78 abc | 24.70 ± 1.64 abcd |
| CE5 | 85.18 ± 1.36 d | 29.81 ± 2.82 aefg |
| CE10 | 87.08 ± 2.01 a | 37.55 ± 1.44 abeh |
| CE20 | 88.12 ± 1.49 b | 39.35 ± 2.16 afc |
| CE30 | 90.03 ± 1.46 cd* | 49.51 ± 3.04 adgh* |
| EM1 | 89.86 ± 1.67 AB | 45.35 ± 1.66 ABCD |
| EM2 | 91.95 ± 1.55 | 74.40 ± 1.48 EF* |
| EM3 | 92.66 ± 1.34 | 75.81 ± 1.99 GH* |
| EM4 | 94.22 ± 2.16 A | 87.04 ± 2.22 EG* |
| EM5 | 94.68 ± 1.09 B* | 86.89 ± 2.06 FH* |
| Enzymes | Process | CE (g/L) | YE/FORBI (g Ethanolkg FORBI) | YE/S (g Ethanol/g util.carb.) | FE (%) |
|---|---|---|---|---|---|
| CE10 | SSF | 9.26 ± 0.80 ab | 84.55 ± 7.28 AB | 0.39 ± 0.03 | 77.29 ± 5.88 |
| SHF | 9.70 ± 0.62 cd | 88.50 ± 5.62 CD | 0.41 ± 0.02 | 81.01 ± 4.31 | |
| CE30 | SSF | 12.73 ± 0.76 ac* | 116.21 ± 6.91 AC* | 0.40 ± 0.02 | 78.86 ± 4.71 |
| SHF | 12.07 ± 1.00 bd+ | 110.21 ± 9.12 BD+ | 0.40 ± 0.01 | 79.24 ± 2.75 | |
| EM3 | SSF | 14.15 ± 1.02 + | 129.12 ± 9.30 + | 0.40 ± 0.02 | 79.20 ± 4.51 |
| SHF | 14.47 ± 0.69 *+ | 132.04 ± 6.26 *+ | 0.39 ± 0.02 | 77.30 ± 3.92 | |
| EM5 | SSF | 15.45 ± 0.75 *+ | 141.06 ± 6.81 *+ | 0.40 ± 0.03 | 79.25 ± 4.92 |
| SHF | 15.41 ± 0.64 *+ | 140.69 ± 5.80 *+ | 0.42 ± 0.02 | 82.59 ± 4.71 |
| CE (g/L) | Carbohydrates Uptake (%) | YE/FORBI (g Ethanol/kg FORBI) | YE/S (g Ethanol/g util.carb.) | FE (%) |
|---|---|---|---|---|
| 15.09 ± 0.62 | 88.08 ± 2.68 | 138.67 ± 5.69 | 0.40 ± 0.01 | 77.98 ± 2.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntaikou, I.; Antonopoulou, G.; Lyberatos, G. Sustainable Second-Generation Bioethanol Production from Enzymatically Hydrolyzed Domestic Food Waste Using Pichia anomala as Biocatalyst. Sustainability 2021, 13, 259. https://doi.org/10.3390/su13010259
Ntaikou I, Antonopoulou G, Lyberatos G. Sustainable Second-Generation Bioethanol Production from Enzymatically Hydrolyzed Domestic Food Waste Using Pichia anomala as Biocatalyst. Sustainability. 2021; 13(1):259. https://doi.org/10.3390/su13010259
Chicago/Turabian StyleNtaikou, Ioanna, Georgia Antonopoulou, and Gerasimos Lyberatos. 2021. "Sustainable Second-Generation Bioethanol Production from Enzymatically Hydrolyzed Domestic Food Waste Using Pichia anomala as Biocatalyst" Sustainability 13, no. 1: 259. https://doi.org/10.3390/su13010259