Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production





Abstract
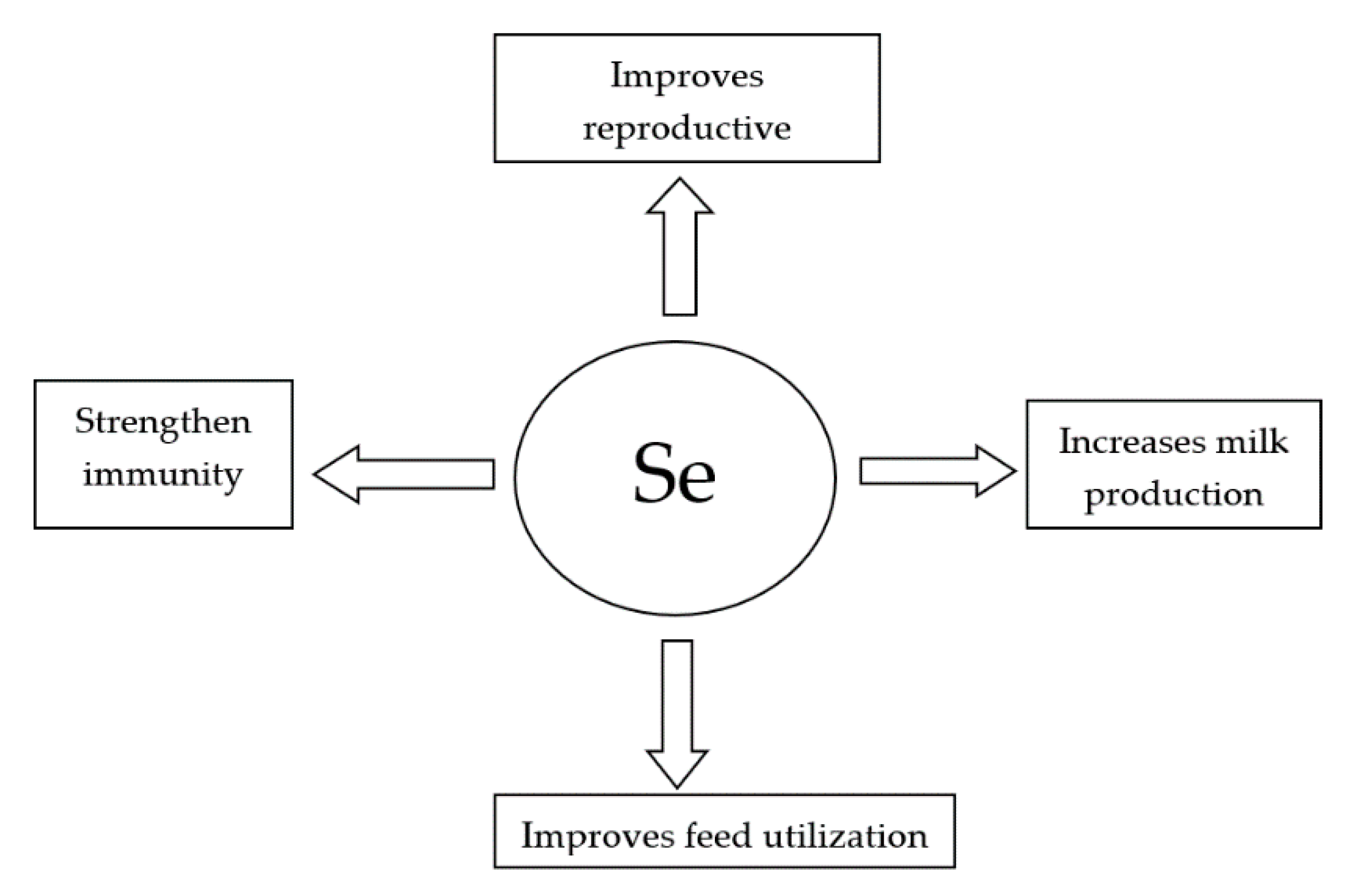
:1. Introduction
2. Absorption, Distribution, Metabolism, and Excretion of Se
3. Role of Se in Antioxidant Defense Mechanism
4. Role of Se in Milk Production
5. Role of Se in Feed Intake, Feed Utilization, and Body Growth
6. Role of Se in Cattle Reproductive Performance
7. Toxicity and Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Butterman, W.C.; Brown, R.D. Mineral Commodity Profiles—Selenium; U.S. USGS Numbered Series; USGS Publisher: Reston, Virginia, 2004; p. 20. [Google Scholar]
- Perrone, D.; Monteiro, M.; Nunes, J.C. The Chemistry of Selenium. R. Soc. Chem. 2015, 3–15. [Google Scholar] [CrossRef]
- Akl, M.A.; Ismael, D.S.; El-Asmy, A.A. Precipitate flotation-separation, speciation and hydride generation atomic absorption spectrometric determination of selenium (IV) in food stuffs. Microchem. J. 2006, 83, 61–69. [Google Scholar] [CrossRef]
- Shrivas, K.; Patel, D.K. Ultrasound assisted-hollow fibre liquid-phase microextraction for the determination of selenium in vegetable and fruit samples by using GF-AAS. Food Chem. 2011, 124, 1673–1677. [Google Scholar] [CrossRef]
- Fan, A.M.; Kizer, K.W. Selenium-nutritional, toxicologic, and clinical aspects. West. J. Med. 1990, 153, 160–167. [Google Scholar] [PubMed]
- Vonderheide, A.P.; Wrobel, K.; Kannamkumarath, S.S.; B’Hymer, C.; Montez-Bay’on, M.; Ponce de Leon, C.; Caruso, J.A. Characterization of selenium species in Brazil nuts by HPLC-ICP-MS and ES-MS. J. Agric. Food Chem. 2002, 50, 5722–5728. [Google Scholar] [CrossRef] [PubMed]
- Najafi, N.M.; Seidi, S.; Alizadeh, R.; Tavakoli, H. Inorganic selenium speciation in environmental samples using selective electrodeposition coupled with electrothermal atomic absorption spectrometry. Spectrochim. Acta B 2010, 65, 334–339. [Google Scholar] [CrossRef]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor-3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292. [Google Scholar] [CrossRef]
- Gergely, V.; Montes-Bayon, M.; Fodor, P.; Sanz-Medel, A. Selenium species in aqueous extracts of alfalfa sprouts by two-dimensional liquid chromatography coupled to inductively coupled plasma mass spectrometry and electrospray mass spectrometry detection. J. Agric. Food Chem. 2006, 54, 4524–4530. [Google Scholar] [CrossRef]
- Bodnar, M.; Konieczka, P.; Namieśnik, J. The Properties, Functions, and Use of Selenium Compounds in Living Organisms. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2012, 30, 225–252. [Google Scholar] [CrossRef]
- Puzanowska-Tarasiewicz, H.; Kuzmicka, L.; Tarasiewicz, M. Biological functions of selected elements and their compounds. 3, Zinc—An enzyme component and activator. Pol. Med. Merkur. Organ Pol. Med. Soc. 2009, 27, 419–422. [Google Scholar]
- Zarczy’nska, K.; Sobiech, P.; Radwi ´nska, J.; Rękawek, W. Effects of selenium on animal health. J. Elementol. 2013, 18, 329–340. [Google Scholar]
- Mehdi, Y.; Dufrasne, I. Selenium in Cattle: A Review. Molecules 2016, 21, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, V.S.; Bicalho, M.L.; Pereira, R.V.; Caixeta, L.S.; Knauer, W.A.; Oikonomou, G.; Gilbert, R.O.; Bicalho, R.C. Effect of an injectable trace mineral supplement containing selenium, copper, zinc, and manganese on the health and production of lactating Holstein cows. Vet. J. 2013, 197, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Alonso, M. Trace minerals and livestock: Not too much not too little. Int. Sch. Res. Netw. Vet. Sci. 2012, 704825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöne, F.; Steinhöfel, O.; Weigel, K.; Bergmann, H.; Herzog, E.; Dunkel, S.; Kirmse, R.; Leiterer, M. Selenium in feedstuffs and rations for dairy cows including a view of the food chain up to the consumer. J. Verbrauch. Lebensm. 2013, 8, 271–280. [Google Scholar]
- Lu, J.; Holmgren, A. Selenoproteins. J. Biol. Chem. 2009, 284, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Slavik, P.; Illek, J.; Brix, M.; Hlavicoca, J.; Rajmon, R.; Jilek, F. Influence of organic versus inorganic dietary selenium supplementation on the concentration of selenium in colostrum, milk and blood of beef cows. Acta Vet. Scand. 2008, 50, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.D.; Stewart, R.D. Metabolic studies of (75Se) selenomethionine and (75Se) selenite in the rat. Br. J. Nutr. 1973, 30, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Weiss, W.P. Selenium nutrition of dairy cows: Comparing responses to organic and inorganic selenium forms. In Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s; Lyons, P.T., Jacques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 12–14 May 2003; pp. 333–343. [Google Scholar]
- Tufarelli, V.; Ceci, E.; Laudadio, V. 2-Hydroxy-4-methylselenobutanoic acid as new organic selenium dietary supplement to produce selenium-enriched eggs. Biol. Trace Elem. Res. 2016, 171, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Podoll, K.L.; Bernard, J.B.; Ullrey, D.E.; DeBar, S.R.; Ku, P.K.; Magee, W.T. Dietary selenate versus selenite for cattle, sheep, and horses. J. Anim. Sci. 1992, 70, 1965–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, P.L.; Preston, R.L.; Pfander, W.H. Wholebody retention, tissue distribution and excretion of selenium after oral and intravenous administration in lambs fed varying selenium intakes. J. Nutr. 1969, 97, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galbraith, M.L.; Vorachek, W.R.; Estill, C.T.; Whanger, P.D.; Bobe, G.; Davis, T.Z.; Hall, J.A. Rumen microorganisms decrease bioavailability of inorganic selenium supplements. Biol. Trace Elem. Res. 2015, 171, 338–343. [Google Scholar] [CrossRef]
- Schrauzer, G.N. Selenomethionine: A review of its nutritional significance, metabolism and toxicity. J. Nutr. 2000, 130, 1653–1656. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.T.; Ogra, Y. Metabolic pathway for selenium in the body: Speciation by HPLC-ICP MS with enriched Se. Food Addit. Contam. 2002, 19, 974–983. [Google Scholar] [CrossRef]
- Meschy, F. Nutrition Minérale des Ruminants; Editions Quae: Versaille, France, 2010; p. 208. [Google Scholar]
- Juniper, D.T.; Phipps, R.H.; Ramos-Morales, E.; Bertin, G. Effect of dietary supplementation with selenium-enriched yeast or sodium selenite on selenium tissue distribution and meat quality in beef cattle. J. Anim. Sci. 2008, 86, 3100–3109. [Google Scholar] [CrossRef]
- Combs, G.F. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.G.; Herdt, T.H.; Stowe, H.D. Physical, hematologic, biochemical, and immunologic effects of supranutritional supplementation with dietary selenium in Holstein cows. Am. J. Vet. Res. 1997, 58, 760–764. [Google Scholar]
- Silvestre, F.T.; Rutigliano, H.M.; Thatcher, W.W.; Santos, J.E.P.; Staples, C.R. Florida Ruminant Nutrition Symposium; University of Florida: Gainesville, FL, USA, 2007; pp. 48–61. [Google Scholar]
- Lucia, F.; Pedrosa, C.; Motley, A.K.; Stevenson, T.D.; Hill, K.E. Fecal Selenium Excretion Is Regulated by Dietary Selenium Intake. Biol. Trace Elem. Res. 2012, 149, 377–381. [Google Scholar]
- Burk, R.F.; Hill, K.E. Regulation of Selenium Metabolism and Transport. Ann. Rev. Nutr. 2015, 35, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, L.L., Jr.; Pope, A.L.; Baumann, C.A. Distribution of microgram quantities of selenium in the tissues of the rat, effects of previous selenium intake. J. Nutr. 1966, 88, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Juniper, D.T.; Phipps, R.H.; Jones, A.K.; Bertin, G. Selenium Supplementation of Lactating Dairy Cows: Effect on Selenium Concentration in Blood, Milk, Urine, and Feces. J. Dairy Sci. 2006, 89, 3544–3551. [Google Scholar] [CrossRef]
- Hrbac, J.; Kohen, R. Biological redox activity: Its importance, methods for its quantification and implication for health and disease. Drug Dev. Res. 2000, 50, 516–527. [Google Scholar] [CrossRef]
- Yadav, A.; Kumari, R.; Yadav, A.; Mishra, J.P.; Srivatva, S.; Prabha, S. Antioxidants and its functions in human body—A Review. Res. Environ. Life Sci. 2016, 9, 1328–1331. [Google Scholar]
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of Reactive Oxygen Species in Cell Signaling Pathways. Biochem. Biomed. Asp. Oxidative Modif. 2001, 29, 345–350. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Adwas, A.A.; Elsayed, A.; Azab, A.E.; Quwaydir, F.A. Oxidative stress and antioxidant mechanisms in human body. J. Appl. Biotechnol. 2019, 6, 43–47. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Juniper, D.T. Revisiting Oxidative Stress and the Use of Organic Selenium in Dairy Cow Nutrition. Animals 2019, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F. Selenium in Poultry Nutrition and Health; Academic Publishers: Wageningen, The Netherlands, 2018. [Google Scholar]
- Boxin, O.U.; Dejian, H.; Maureen, A.F.; Elizabeth, K.D. Analysis of antioxidant activities of common vegetables employing oxygen radical Absorbance Capacity (ORAC) and Ferric Reducing Antioxidant Power (FRAP) Assays: A comparative study. J. Agric. Food Chem. 2002, 5, 223–228. [Google Scholar]
- Burk, R.F.; Hill, K.E.; Motley, A.K. Selenoprotein Metabolism and Function: Evidence for More than One Function for Selenoprotein. Pak. J. Nutr. 2003, 133, 1517S–1520S. [Google Scholar] [CrossRef] [PubMed]
- Mezzetti, A.; Ilio, C.D.; Calafiore, A.M.; Aceto, A.; Marzio, L.; Frederici, G.; Cuccurullo, F. Glutathione peroxidase, glutathione reductase and glutathione transferase activities in the human artery, vein and heart. J. Mol. Cardiol. 1990, 22, 935–938. [Google Scholar] [CrossRef]
- Grasso, P.J.; Scholz, R.W.; Erskine, R.J.; Eberhart, R.J. Phagocytosis, bactericidal activity, and oxidative metabolism of mammary neutrophils from dairy cows fed selenium-adequate and selenium-deficient diets. Am. J. Vet. Res. 1990, 51, 269–274. [Google Scholar] [PubMed]
- Gunter, S.A.; Beck, P.A.; Phillips, J.M. Effects of supplementary selenium source on the performance and blood measurements in beef cows and their calves. J. Anim. Sci. 2003, 81, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, J.R.; Boyne, R. Superoxide dismutase and glutathione peroxidase activities in neutrophils from selenium deficient and copper deficient cattle. Life Sci. 1985, 36, 1569–1575. [Google Scholar] [CrossRef]
- Awadeh, F.T.; Kincaid, R.L.; Johnson, K.A. Effect of level and sources of dietary selenium on concentrations of thyroid hormones and immunoglobulins in beef cows and calves. J. Anim. Sci. 1998, 76, 1204–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, W.R. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 2000, 60–61, 449–457. [Google Scholar] [CrossRef]
- Rabiee, A.R.; Lean, I.J.; Stevenson, M.A.; Socha, M.T. Effects of feeding organic trace minerals on milk production and reproductive performance in lactating dairy cows: A meta-analysis. J. Dairy Sci. 2010, 93, 4239–4251. [Google Scholar] [CrossRef] [Green Version]
- Oltramari, C.E.; Pinheiro, M.G.; Miranda, M.S.; Arcaro, J.R.P.; Castelani, L.; Toledo, L.C.; Ambrósio, L.A.; Leme, P.R.; Manella, M.Q.; Júnior, I.A. Selenium Sources in the Diet of Dairy Cows and Their Effects on Milk Production and Quality, on Udder Health and on Physiological Indicators of Heat Stress. Ital. J. Anim. Sci. 2014, 13, 2921. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.B. Serum selenium concentrations and glutathione peroxidase activities in cattle grazing forages of various selenium concentrations. Am. J. Vet. Res. 1985, 46, 1556–1561. [Google Scholar]
- Tufarelli, V.; Laudadio, V. Dietary supplementation with selenium and vitamin E improves milk yield, composition and rheological properties of dairy Jonica goats. J. Dairy Res. 2011, 78, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Salman, S.; Khol-Parisini, A.; Schafft, H.; Lahrssen-Wiederholt, M.; Hulan, H.W.; Dinse, D.; Zentek, J. The role of dietary selenium in bovine mammary gland health and immune function. Anim. Health Res. Rev. 2009, 10, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Souza, F.N.; Blagitz, M.G.; Latorre, A.O.; Mori, C.S.; Sucupira, M.C.A.; Libera, A.M.M.P.D. Effects of in vitro selenium supplementation on blood and milk neutrophils from dairy cows. Pesqui. Vet. Bras. 2012, 32, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, H.H.; Safwat, M.A.; El-Sysy, M.A.I.; Al-Metwaly, M.A. Effect of selenium and vitamin E supplementation as a nutritional treatment for some physiological and productive traits of Holstein dairy cows under Egyptian summer conditions. J. Egypt. Acad. Soc. Environ. Dev. 2016, 17, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Cordova-Izquierdo, C.G.R.L.; Campos, V.M.X.; Suarez, S.C.; Cordova-Jimenez, C.A.; Cordova-Jimenez, M.S. Effect of gonadotophin releasing factor and antioxidants on the rate of estrous repetition dairy cows. Aust. J. Basic Appl. Sci. 2010, 408, 1282–1287. [Google Scholar]
- Wang, C.; Liu, Q.; Yang, W.Z.; Dong, Q.; Yang, X.M.; He, D.C.; Huang, Y.X. Effects of selenium yeast on rumen fermentation, lactation performance and feed digestibilities in lactating dairy cows. Livest. Sci. 2009, 126, 239–244. [Google Scholar] [CrossRef]
- Wilde, D. Influence of macro and micro minerals in the peri-parturient period on fertility in dairy cattle. Anim. Reprod. Sci. 2006, 96, 240–249. [Google Scholar] [CrossRef]
- Ullah, H.; Khan, R.U.; Mobashar, M.; Ahmad, S.; Sajid, A.; Khan, N.U.; Usman, T.; Khattak, I.; Hamayun, K. Effect of yeast-based selenium on blood progesterone, metabolites and milk yield in Achai dairy cows. Ital. J. Anim. Sci. 2019, 18, 1445–1450. [Google Scholar] [CrossRef] [Green Version]
- Eulogio, G.L.J.; Alberto, S.O.J.; Hugo, C.V.; Antonio, C.N.; Alejandro, C.; Juan, M.Q. Effects of Selenium and vitamin E in the production, physiochemical composition and somatic cell count in milk of Ayrshire cows. J. Anim. Vet. Adv. 2012, 11, 687–691. [Google Scholar]
- Chauhana, S.S.; Ponnampalama, E.N.; Celi, P.; Hopkins, D.L.; Leurya, B.J.; Dunsheaa, F.R. High dietary vitamin E and selenium improves feed intake and weight gain of finisher lambs and maintains redox homeostasis under hot conditions. Small Rumin. Res. 2016, 137, 17–23. [Google Scholar] [CrossRef]
- Smith, J.F.; Brouk, M.J. Factors affecting dry matter intake by lactating dairy cows. Kans. Agric. Exp. Stn. Res. Rep. 2000, 2, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Alhidary, I.; Shini, S.; Al Jassim, R.; Gaughan, J. Effect of various doses of injected selenium on performance and physiological responses of sheep to heat load. J. Anim. Sci. 2012, 90, 2988–2994. [Google Scholar] [CrossRef] [PubMed]
- Ganie, A.A.; Baghel, R.P.S.; Mudgal, V.; Sheikh, G.G. Effect of Selenium Supplementation on Growth and Nutrient Utilization in Buffalo Heifers. Anim. Nutr. Feed Technol. 2010, 10, 255–259. [Google Scholar]
- Surai, P.F. Selenium in poultry nutrition-1. Antioxidant properties, deficiency and toxicity. World Poult. Sci. J. 2002, 58, 333–347. [Google Scholar] [CrossRef]
- Willshire, J.A.; Payne, J.H. Selenium and vitamin E in dairy cows—A review. Cattle Pract. 2011, 19, 22–30. [Google Scholar]
- Rooke, J.A.; Robinson, J.J.; Arthur, J.R. Effects of vitamin E and selenium on the performance and immune status of ewes and lambs. J. Agric. Sci. 2004, 142, 253–262. [Google Scholar] [CrossRef]
- Gartner, R.; Gasnier, B.; Dietrich, J.; Krebs, B.; Angstwurm, M. Selenium supplementation in patients with autoimmune thyroiditis decreases thyroid peroxidase antibodies concentrations. J. Clin. Endocrinol. Metab. 2007, 87, 1687–1691. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, L.; Riese, C.; Michaelis, M.; Griebert, E.; Klein, M.; Sapin, R.; Schweizer, U.; Köhrle, J. Synthesis and metabolism of thyroid hormones is preferentially maintained in seleniumdeficient transgenic mice. Endocrinology 2007, 147, 1306–1313. [Google Scholar] [CrossRef] [Green Version]
- Taheri, Z.; Karimi, S.; Mehrban, H.; Moharrery, A. Supplementation of different selenium sources during early lactation of native goats and their effects on nutrient digestibility, nitrogen and energy status. J. Appl. Anim. Res. 2018, 46, 64–68. [Google Scholar] [CrossRef]
- Weiss, W.P. Selenium sources for dairy cattle. In Proceedings of the Tri–State Dairy Nutrition Conference, Fort Wayne, IN, USA, 2–3 May 2005. [Google Scholar]
- Alimohamady, R.; Aliarabi, H.; Bahari, A.; Dezfoulian, A.H. Influence of Different Amounts and Sources of Selenium Supplementation on Performance, Some Blood Parameters, and Nutrient Digestibility in Lambs. Biol. Trace Elem. Res. 2013, 154, 45–54. [Google Scholar] [CrossRef]
- Do, C.; Wasana, N.; Cho, K.; Choi, Y.; Choi, T.; Park, B.; Lee, D. The Effect of Age at First Calving and Calving Interval on Productive Life and Lifetime Profit in Korean Holsteins. Asian Australas. J. Anim. Sci. 2013, 26, 1511–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedaoo, M.; Khllare, K.; Meshram, M.; Sahatpure, S.; Patil, M. Study of some serum trace minerals in cyclic and noncyclic surti buffaloes. Vet. World 2008, 1, 71–72. [Google Scholar]
- Kamada, H. Effects of selenium-rich yeast supplementation on the plasma progesterone levels of postpartum dairy cows. Asian Australas. J. Anim. Sci. 2017, 30, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pursley, J.R.; Martins, J.P. Impact of circulating concentrations of progesterone and antral age of the ovulatory follicle on fertility of high-producing lactating dairy cows. Reprod. Fertil. Dev. 2011, 24, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inskeep, E.K. Preovulatory, and postmaternal recognition effects of concentrations of progesterone on embryonic survival in the cow. J. Anim. Sci. 2004, 82, 24–39. [Google Scholar] [CrossRef]
- Kamada, H.; Ikumo, H. Effect of selenium on cultured bovine luteal cells. Anim. Reprod. Sci. 1997, 46, 203–211. [Google Scholar] [CrossRef]
- Bajaj, N.K.; Sharma, N. Endocrine Causes of Early Embryonic Death: An Overview. Curr. Res. Dairy Sci. 2011, 3, 1–24. [Google Scholar] [CrossRef]
- Ganie, A.A.; Baghel, R.P.S.; Mudgal, V.; Aarif, O.; Sheikh, G.G. Effect of selenium supplementation on reproductive performance and hormonal profile in buffalo heifers. Ind. J. Anim. Res. 2014, 48, 27–30. [Google Scholar] [CrossRef]
- Thatcher, W.; Santos, J.E.P.; Staples, C.R. Dietary manipulations to improve embryonic survival in cattle. Theriogenology 2011, 76, 1619–1631. [Google Scholar] [CrossRef]
- Dimri, U.; Ranjan, R.; Sharma, M.C.; Varshney, V.P. Effect of vitamin E and selenium supplementation on oxidative stress indices and cortisol level in blood in water buffaloes during pregnancy and early postpartum period. Trop. Anim. Health Prod. 2010, 42, 405–410. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, R.; Malevu, T.D.; Sochor, J.; Baron, M.; Melcova, M.; Zidkova, J.; et al. A Summary of New Findings on the Biological Effects of Selenium in Selected Animal Species—A Critical Review. Int. J. Mol. Sci. 2017, 18, 2209. [Google Scholar] [CrossRef] [PubMed]
- Mézes, M.; Balogh, K. Prooxidant mechanisms of selenium toxicity—A review. Acta Biol. Szeged. 2009, 53, 15–18. [Google Scholar]
- Hartikainen, H. Biogeochemistry of selenium and its impact on food chain quality and human health. J. Trace Elem. Med. Biol. 2005, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A.; Pendias, H. Biogeochemia Pierwiastk’ow‘sladowych; PWN Press: Warsaw, Poland, 1999. [Google Scholar]
- Mihajlović, M. Toksicnost selena kod domaćih zivotinja [Selenium toxicity in domestic animals]. Glas Srp Akad Nauka Med. 1992, 42, 131–144. [Google Scholar]
- Neve, J.; Favier, A. Selenium in medecine and biology. In Proceedings of the Second International Congress on Trace Elements in Medecine and Biology, Avoriaz, France, March 1988. [Google Scholar]
- Claude, J.B. Introduction à la Nutrition des Animaux Domestiques; Tec & Doc/EM Inter: Paris, France, 2002. [Google Scholar]
- Rogers, P.A.M.; Arora, S.P.; Fleming, G.A.; Crinion, R.A.P.; McLaughlin, J.G. Selenium toxicity in farm animals: Treatment and prevention. Ir. Vet. J. 1990, 43, 151–153. [Google Scholar]
- James, L.F.; James, L. Shupe. Selenium Poisoning in Livestock. Rangelands 1984, 6, 64–67. [Google Scholar]
- Nair, M.P.N.; Schwartz, S.A. Immunoregulation of natural and lymphokine-activated killer cells by selenium. Immunopharmacology 1990, 19, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Abuelo, A.; Alves-Nores, V.; Hernandez, J.; Muiño, R.; Benedito, J.L.; Castillo, C. Effect of parenteral antioxidant supplementation during the dry period on postpartum glucose tolerance in dairy cows. J. Vet. Intern. Med. 2016, 30, 892–898. [Google Scholar]
- Rizzo, A.; Pantaleo, M.; Mutinati, M.; Minoia, G.; Trisolini, C.; Ceci, E.; Sciorsci, R.L. Blood and milk oxidative status after administration of different antioxidants during early postpartum in dairy cows. Res. Vet. Sci. 2013, 95, 1171–1174. [Google Scholar] [CrossRef]
- Szpunar, J.; Lobinski, R.; Prange, A. Hyphenated techniques for elemental speciation in biological systems. Appl. Spectrosc. 2003, 57, 102–112. [Google Scholar] [CrossRef]
- Najafnejad, B.; Aliarabi, H.; Tabatabaei, M.M.; Alimohamady, R. Effects of different forms of Selenium supplementation on production performance and nutrient digestibility in lactating dairy cattle. In Proceedings of the Second International Conference on Agriculture and Natural Resources, Kermanshah, Iran, 25–26 December 2013. [Google Scholar]
- Kamada, H.; Hondate, K. Effect of dietary Selenium supplementation on plasma progesterone concentration in cows. J. Vet. Med. Sci. 1998, 60, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.A.; Bobe, G.; Vorachek, W.R.; Kasper, K.; Traber, M.G.; Mosher, W.D.; Pirelli, G.J.; Gamroth, M. Effect of supranutritional organic selenium supplementation on postpartum blood micronutrients, antioxidants, metabolites, and inflammation biomarkers in selenium-replete dairy cows. Biol. Trace Elem. Res. 2014, 161, 272–287. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Parameter | Form of Se | Dose of Se | Effect | Reference |
|---|---|---|---|---|
| Milk production | Organic | 0.15 mg/kg DMI | Increased | [61] |
| Organic | 0.3 mg/kg DMI | Increased | [61,63,64] | |
| Organic | 0.4 mg/kg DMI | Decreased | [63] | |
| Neutrophil function | Inorganic/sodium selenite | 10 μM | Improved | [58] |
| Organic | 0.3 mg/kg DMI | Increased | [33] | |
| Adaptive immunity | Organic | 0.3 mg/kg DMI | Improved | [33] |
| Somatic cell count | Organic | 0.3 mg/kg DMI | Decreased | [33] |
| Milk fat | Organic | 0.3 mg/kg DMI | Increased | [100] |
| Nano particles | 0.3 mg/kg DMI | Increased | [100] | |
| Organic | 0.3 mg/kg DMI | Increased | [33] | |
| Milk protein | Organic | 0.3 mg/kg DMI | Increased | [100] |
| Nano particles | 0.3 mg/kg DMI | Increased | [100] | |
| Digestibility (DM, OM, NDF, Ether extract, NFC) | Organic | 0.3 mg/kg DMI | Increased | [100] |
| Nano particles | 0.3 mg/kg DMI | Increased | [100] | |
| Plasma P4 | Inorganic | 0.5 mg/kg DMI | Increased | [101] |
| Serum Se | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Plasma Se | Organic | 0.3 mg/kg DMI | Increased | [33] |
| Whole blood Se concentration | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Serum amyloid A | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Erythrocyte glutathione | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Serum albumin concentration | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Serum cholesterol concentration | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Decreased | [102] |
| α-tocopherol/cholesterol ratio | Organic + Inorganic | 105 mg/week + 0.3 mg/kg DMI | Increased | [102] |
| Uterine health | Organic | 0.3 mg/kg DMI | Improved | [33] |
| Second service pregnancy rate | Organic | 0.3 mg/kg DMI | increased | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, H.; Khan, R.U.; Tufarelli, V.; Laudadio, V. Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production. Sustainability 2020, 12, 10693. https://doi.org/10.3390/su122410693
Ullah H, Khan RU, Tufarelli V, Laudadio V. Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production. Sustainability. 2020; 12(24):10693. https://doi.org/10.3390/su122410693
Chicago/Turabian StyleUllah, Hammad, Rifat Ullah Khan, Vincenzo Tufarelli, and Vito Laudadio. 2020. "Selenium: An Essential Micronutrient for Sustainable Dairy Cows Production" Sustainability 12, no. 24: 10693. https://doi.org/10.3390/su122410693