Biotreatment of Poultry Waste Coupled with Biodiesel Production Using Suspended and Attached Growth Microalgal-Based Systems

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of Poultry Litter Extract (PLE)
2.2. The Microbial Consortium Used in the PLE Treatment and Treatment Conditions
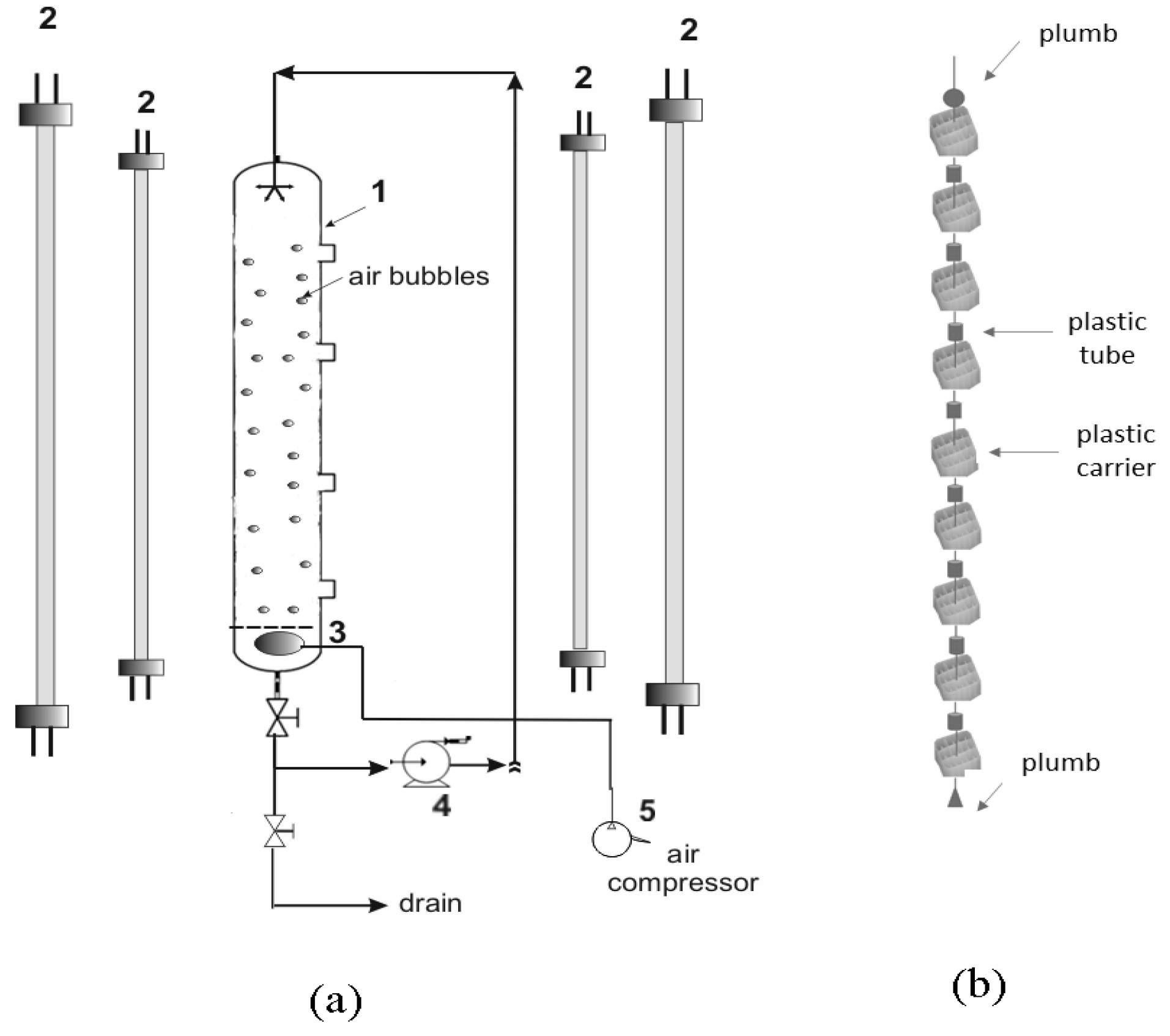
2.3. Experimental Set up
2.3.1. Laboratory-Scale Photobioreactors
2.3.2. Pilot-Scale Photobioreactors
2.4. Analytical Procedures
2.5. Statistical Analysis
3. Results and Discussion
3.1. Laboratory-Scale Suspended Growth System
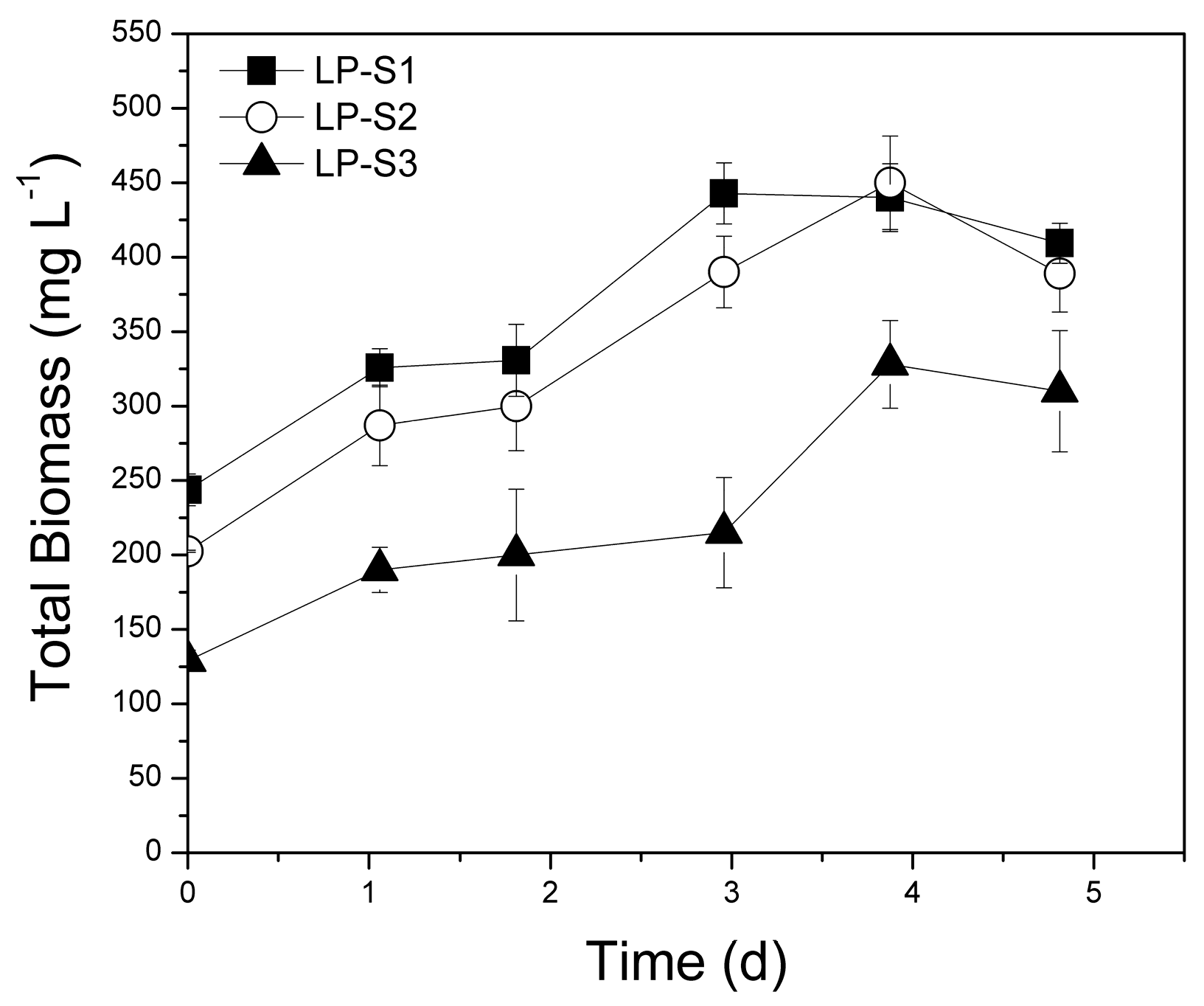
3.1.1. Biomass Production
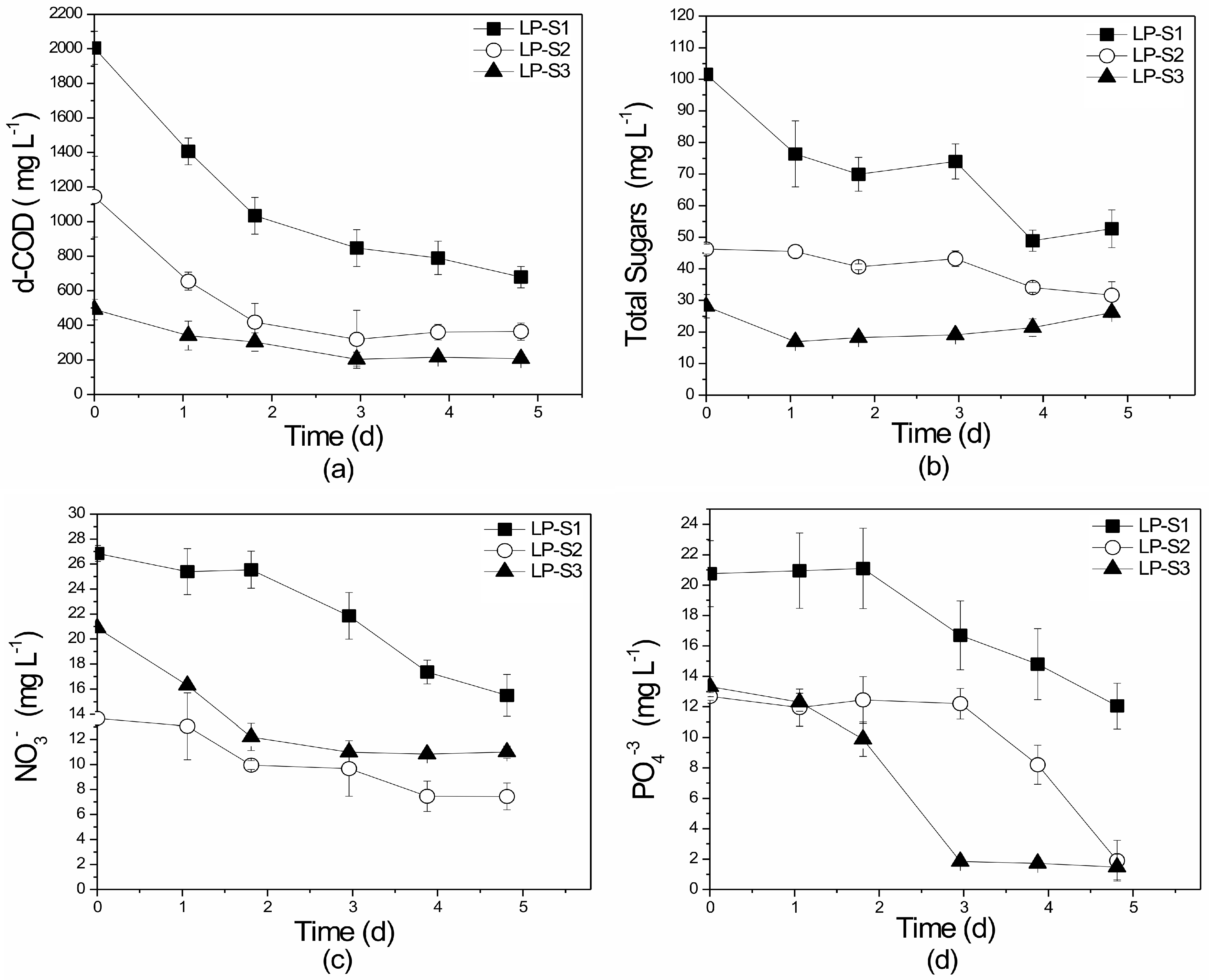
3.1.2. Nutrient and Organic Load Removal
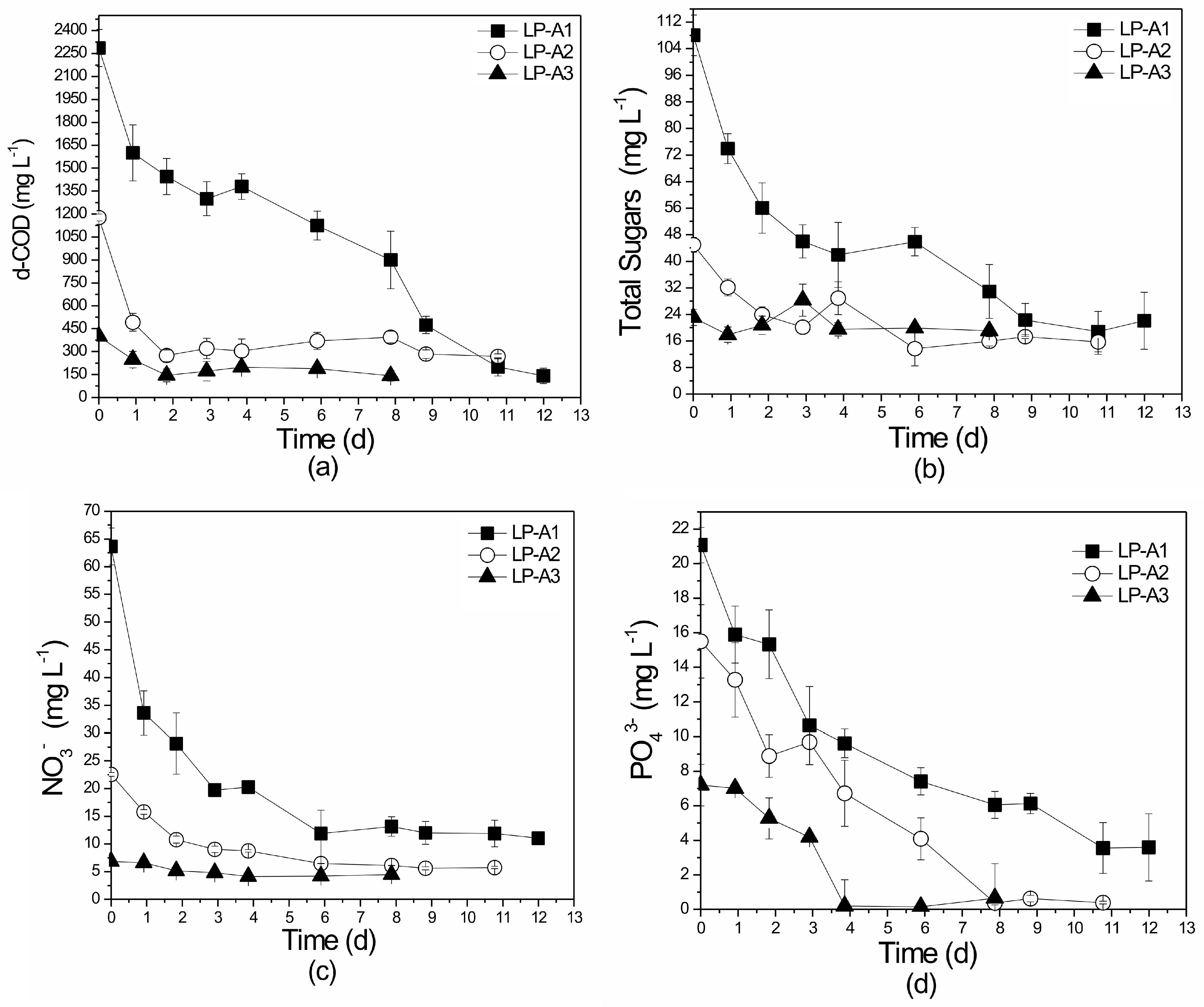
3.2. Laboratory-Scale Attached Growth System
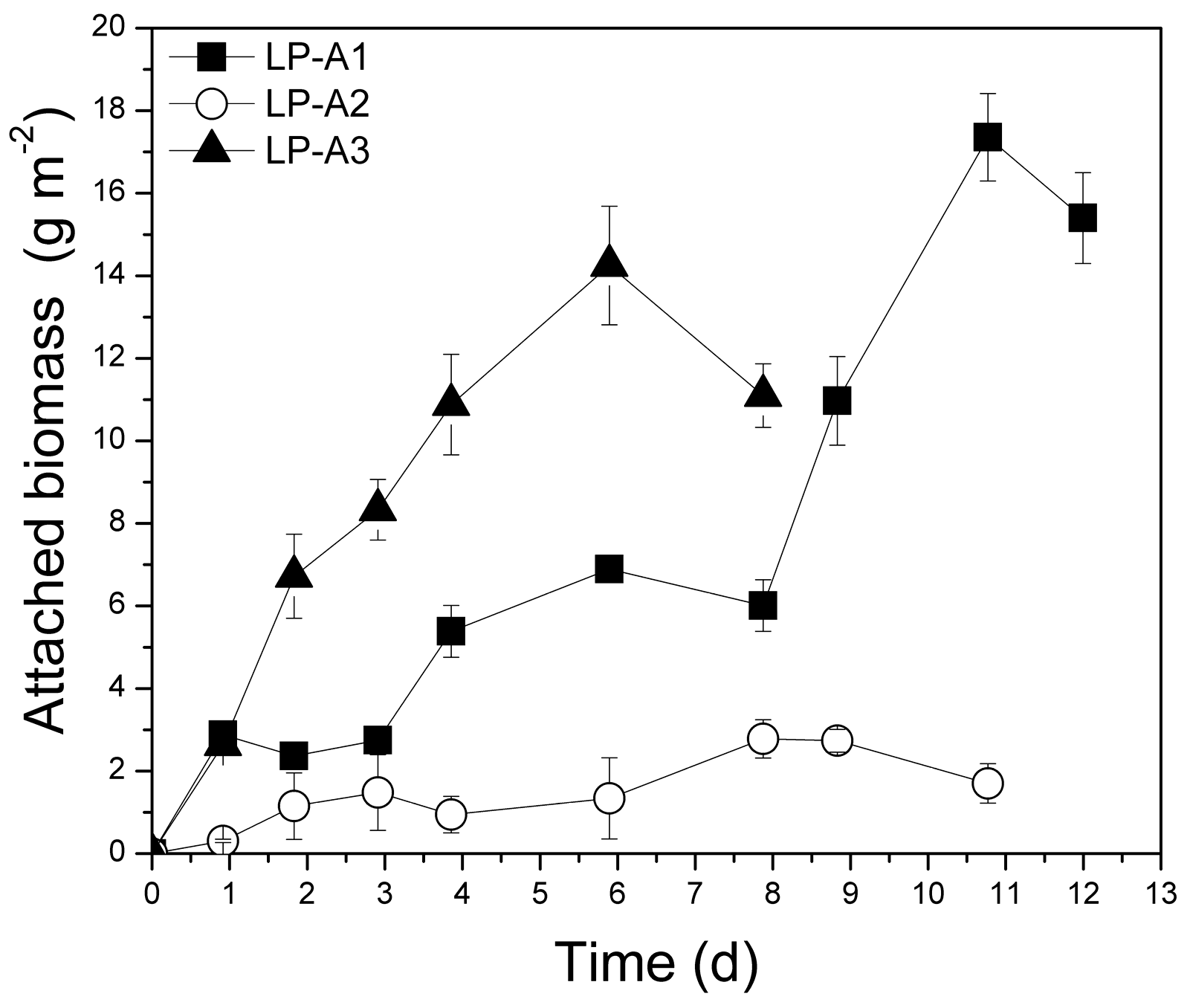
3.2.1. Biomass Production
3.2.2. Nutrient and Organic Load Removal
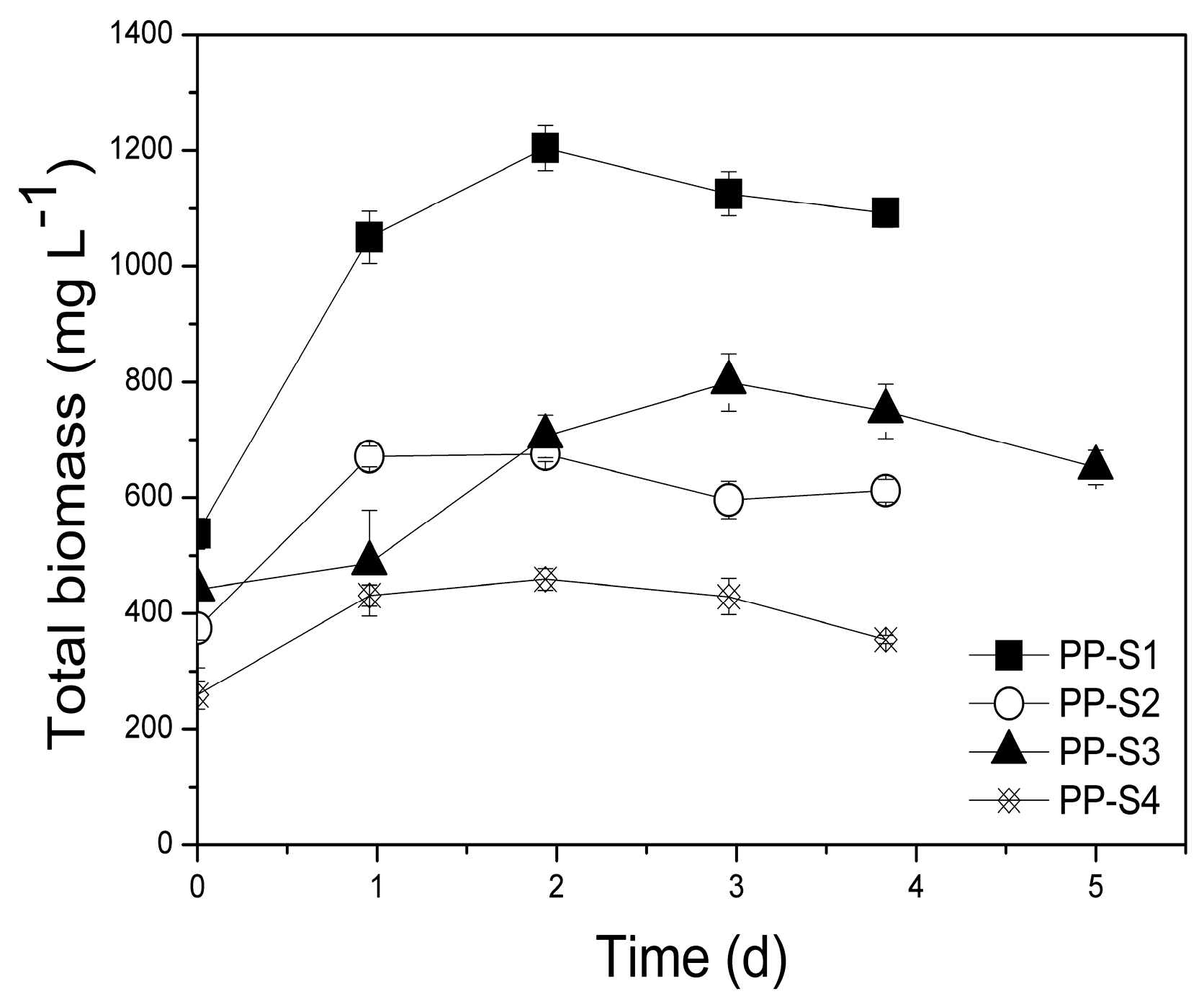
3.3. Pilot-Scale Suspended Growth System
3.3.1. Biomass Production
3.3.2. Nutrient and Organic Load Removal
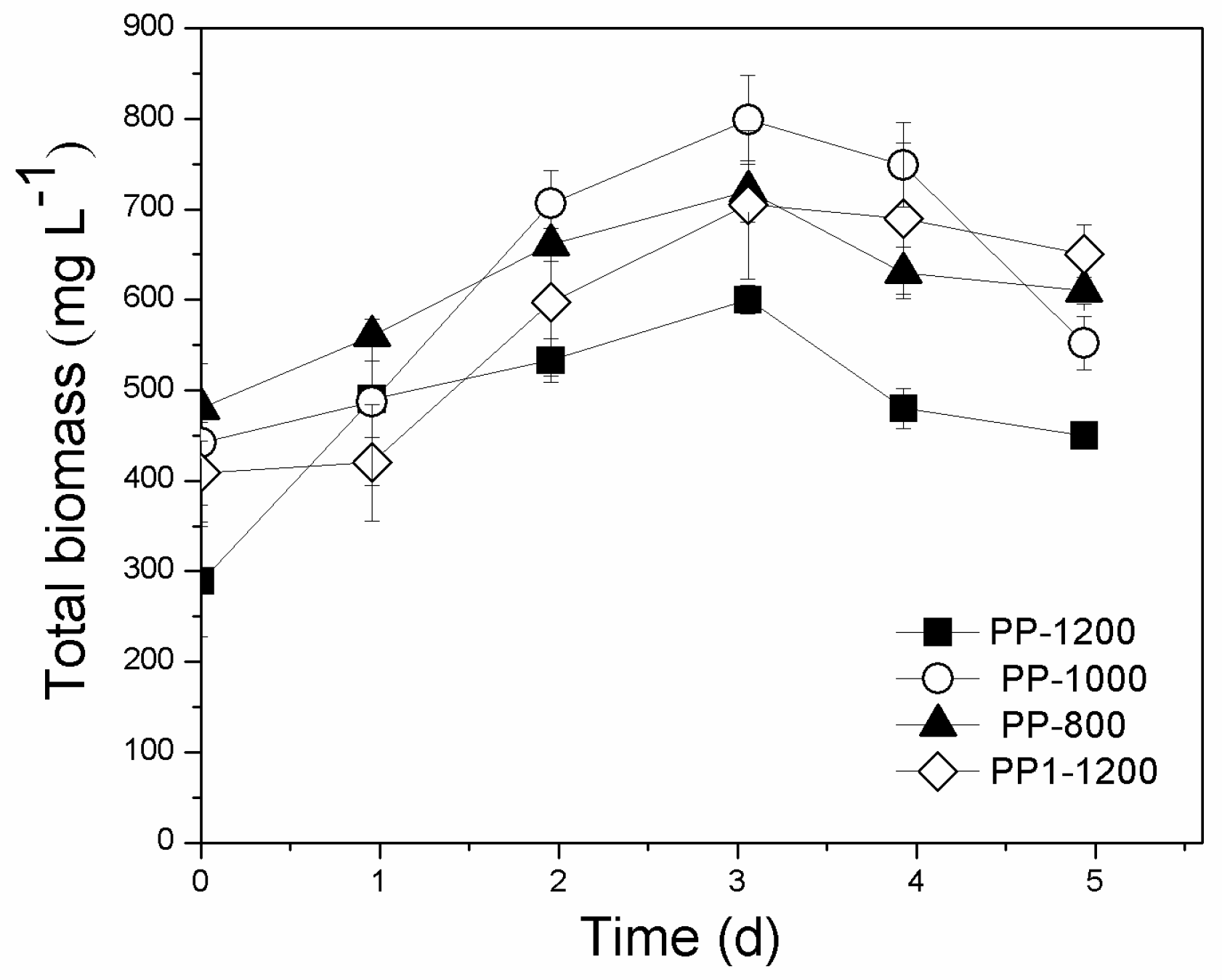
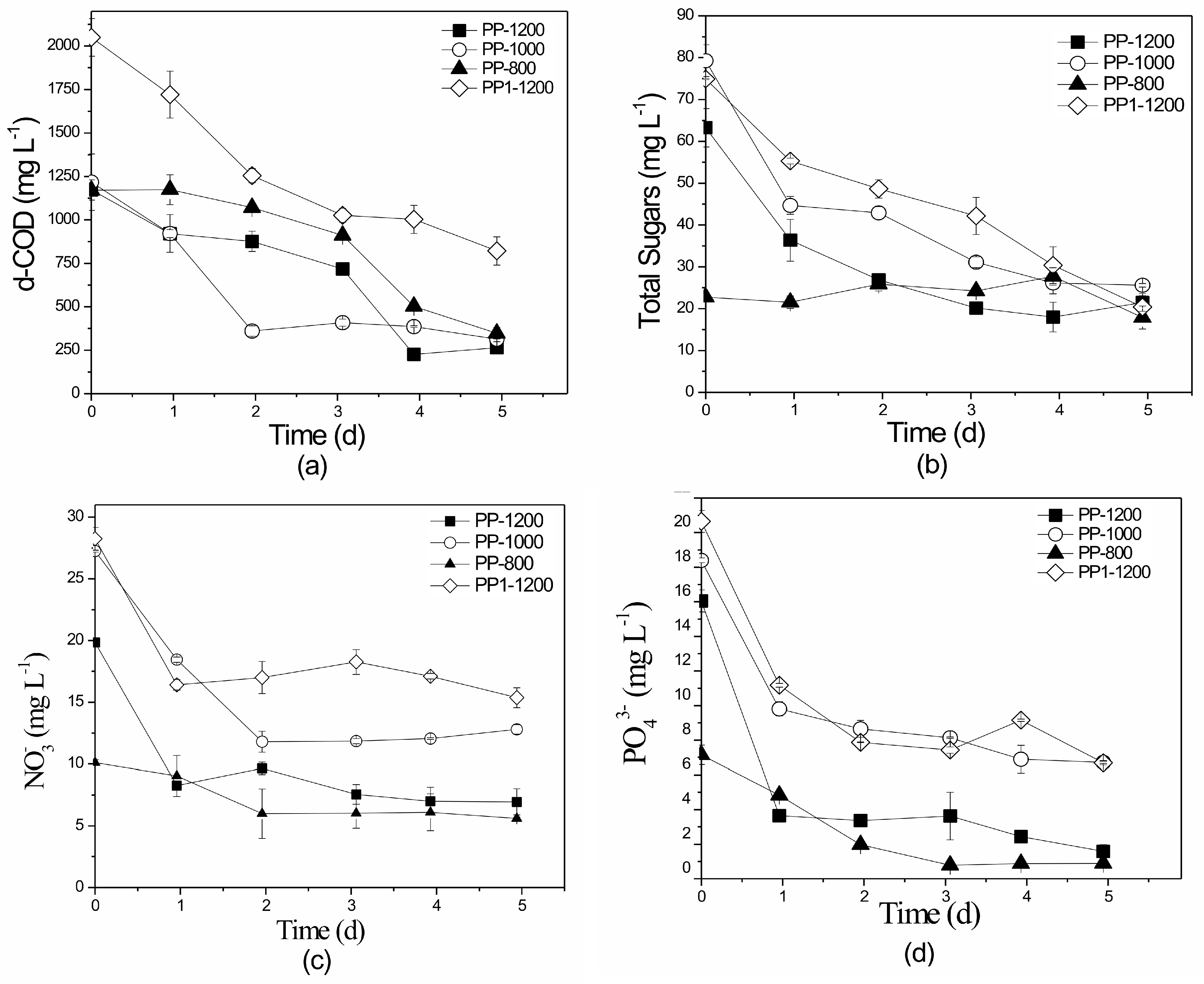
3.4. Pilot-Scale Attached Growth System
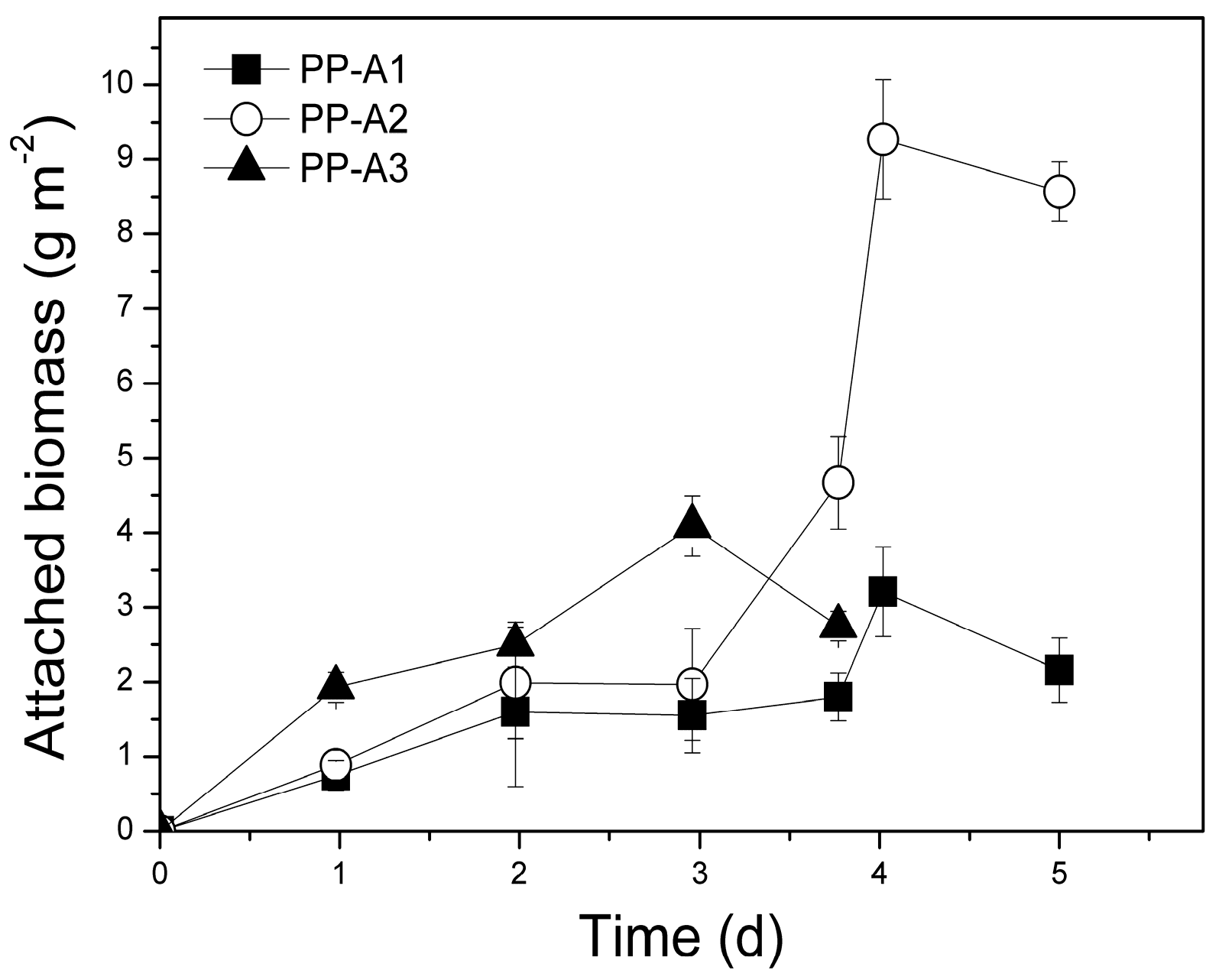
3.4.1. Biomass Production
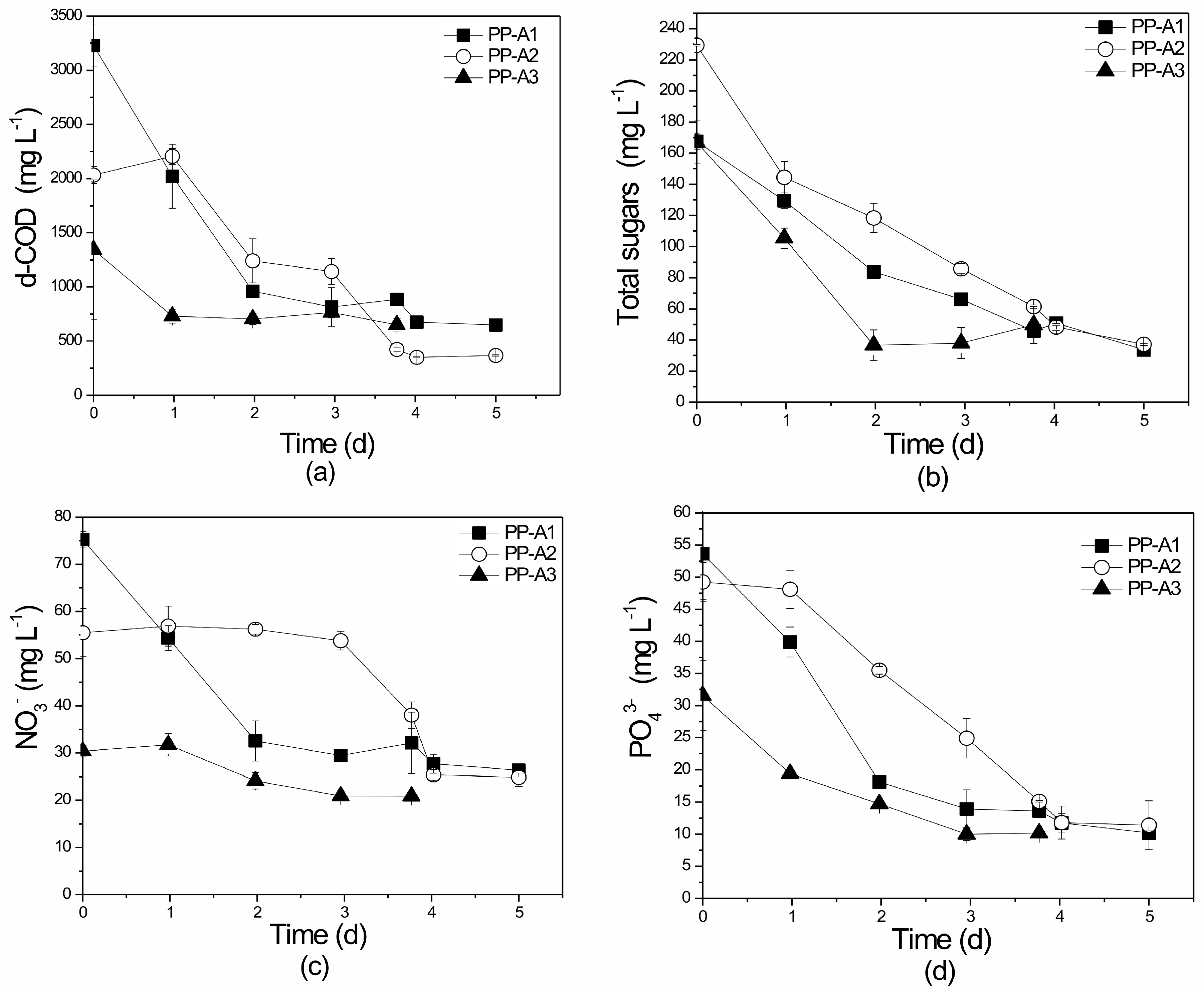
3.4.2. Nutrient and Organic Load Removal
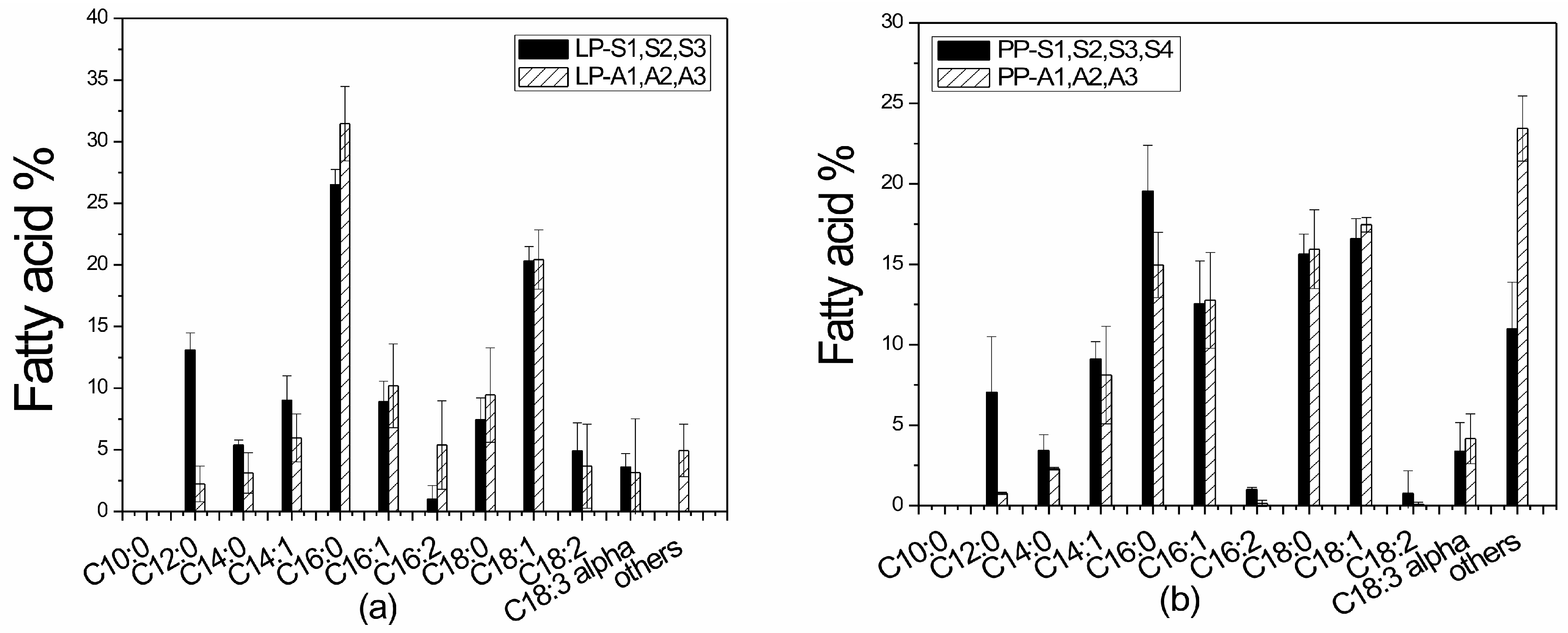
3.5. Lipid Production and Fatty Acid Profile
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nicholson, F.A.; Chambers, B.J.; Smith, K.A. Nutrient composition of poultry manures in England and Wales. Bioresour. Technol. 1996, 58, 279–284. [Google Scholar] [CrossRef]
- Markou, G.; Iconomou, D.; Muylaert, K. Applying raw poultry litter leachate for the cultivation of Arthrospira platensis and Chlorella vulgaris. Algal Res. 2016, 13, 79–84. [Google Scholar] [CrossRef]
- Markou, G. Fed-batch cultivation of Arthrospira and Chlorella in ammonia-rich wastewater: Optimization of nutrient removal and biomass production. Bioresour. Technol. 2015, 193, 35–41. [Google Scholar] [CrossRef]
- Singh, M.; Reynolds, D.L.; Das, K.C. Microalgal system for treatment of effluent from poultry litter anaerobic digestion. Bioresour. Technol. 2011, 102, 10841–10848. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and wastewater treatment. Saudi J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chen, G.; Tao, Y.; Wang, J. Application of effluent from WWTP in cultivation of four microalgae for nutrients removal and lipid production under the supply of CO2. Renew. Energy 2020, 149, 708–715. [Google Scholar] [CrossRef]
- Shabana, E.F.; Taofik, F.E.; Shalaby, H.S. Biodiesel production and nutrient removal by microalgae cultured on poultry waste. Egyptian J. Phycol. 2019, 20. Available online: https://scholar.cu.edu.eg/sites/default/files/hanaas/files/ejphycology-2019-08-09_1567622747.pdf (accessed on 15 May 2020).
- Bhatnagar, A.; Chinnasamy, S.; Singh, M.; Das, K.C. Renewable biomass production by mixotrophic algae in the presence of various carbon sources and wastewaters. Appl. Energy 2011, 88, 3425–3431. [Google Scholar] [CrossRef]
- Mandal, S.; Mallick, N. Biodiesel Production by the Green Microalga Scenedesmus obliquus in a Recirculatory Aquaculture System. Appl. Environ. Microbiol. 2012, 78, 5929–5934. [Google Scholar] [CrossRef] [Green Version]
- Bhati, R.; Mallick, N. Carbon dioxide and poultry waste utilization for production of polyhydroxyalkanoate biopolymers by Nostoc muscorum Agardh: A sustainable approach. J. Appl. Phycol. 2016, 28, 161–168. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Sarmah, A.K.; Sen, R. Performance evaluation of an outdoor algal biorefinery for sustainable production of biomass, lipid and lutein valorizing flue-gas carbon dioxide and wastewater cocktail. Bioresour. Technol. 2019, 283, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Mallick, N. Waste Utilization and Biodiesel Production by the Green Microalga Scenedesmus obliquus. Appl. Environ. Microbiol. 2011, 77, 374–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lin, L.; Lu, H.; Liu, Z.; Duan, N.; Dong, T.; Xiao, H.; Li, B.; Xu, P. Microalgae cultivation and culture medium recycling by a two-stage cultivation system. Front. Environ. Sci. Eng. 2018, 12, 14. [Google Scholar] [CrossRef]
- Mahadevaswamy, M.; Venkataraman, L.V. Bioconversion of Poultry Droppings for Biogas and Algal Production. Agric. Wastes 1986, 18, 93–101. [Google Scholar] [CrossRef]
- Iyovo, G.D.; Du, G.; Chen, J. Poultry Manure Digestate Enhancement of Chlorella Vulgaris Biomass under Mixotrophic Condition for Biofuel Production. J. Microb. Biochem. Technol. 2010, 2, 51–57. [Google Scholar] [CrossRef]
- Wang, M.; Wu, Y.; Li, B.; Dong, R.; Lu, H.; Zhou, H.; Cao, W. Pretreatment of poultry manure anaerobic-digested effluents by electrolysis, centrifugation and autoclaving process for Chlorella vulgaris growth and pollutants removal. Environ. Technol. 2015, 36, 837–843. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, M.; Cao, W.; Li, B.; Liu, Z.; Lu, H. Optimization of Chlorella pyrenoidosa Y3 biomass production in poultry waste anaerobic-digested effluents using a response surface methodology. Desalin. Water Treat. 2016, 57, 8711–8719. [Google Scholar] [CrossRef]
- Markou, G.; Iconomou, D.; Sotiroudis, T.; Israilides, C.; Muylaert, K. Exploration of using stripped ammonia and ash from poultry litter for the cultivation of the cyanobacterium Arthrospira platensis and the green microalga Chlorella vulgaris. Bioresour. Technol. 2015, 196, 459–468. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotehnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Lowrey, J.; Brooks, M.S.; McGinn, P.J. Heterotrophic and mixotrophic cultivation of microalgae for biodiesel production in agricultural wastewaters and associated challenges—A critical review. J. Appl. Phycol. 2015, 27, 485–1498. [Google Scholar] [CrossRef]
- Kothari, R.; Prasad, R.; Kumar, V.; Singh, D.P. Production of biodiesel from microalgae Chlamydomonas polypyrenoideum grown on dairy industry wastewater. Bioresour. Technol. 2013, 144, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Bellou, S.; Baeshen, M.N.; Elazzazy, A.M.; Aggeli, D.; Sayegh, F.; Aggelis, G. Microalgal lipids biochemistry and biotechnological perspectives. Biotechnol. Adv. 2014, 32, 1476–1493. [Google Scholar] [CrossRef] [PubMed]
- Dourou, M.; Tsolcha, O.; Tekerlekopoulou, A.; Bokas, D.; Aggelis, G. Fish farm effluents are suitable growth media for Nannochloropsis gaditana, a polyunsaturated fatty acid producing microalga. Eng. Life Sci. 2018, 18, 851–860. [Google Scholar] [CrossRef] [Green Version]
- Kothari, R.; Pathak, V.V.; Kumar, V.; Singh, D.P. Experimental study for growth potential of unicellular alga Chlorella pyrenoidosa on dairy wastewater: An integrated approach for treatment and biofuel production. Bioresour. Technol. 2012, 116, 466–470. [Google Scholar] [CrossRef]
- Gross, M.; Jarboe, D.; Wen, Z. Biofilm-based algal cultivation systems. Appl. Microbiol. Biotechnol. 2015, 99, 5781–5789. [Google Scholar] [CrossRef]
- Mantzorou, A.; Ververidis, F. Microalgal biofilms: A further step over current microalgal cultivation techniques. Sci. Total Environ. 2019, 651, 3187–3201. [Google Scholar] [CrossRef]
- Tsolcha, O.N.; Tekerlekopoulou, A.G.; Akratos, C.S.; Aggelis, G.; Genitsaris, S.; Moustaka-Gouni, M.; Vayenas, D.V. Agro-industrial wastewater treatment with simultaneous biodiesel production in attached growth systems using a mixed microbial culture. Water 2018, 10, 1693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.L.; Tay, J.H. Microalgal-bacterial consortia: From interspecies interactions to biotechnological applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [Google Scholar] [CrossRef]
- Shen, Y.; Zhu, W.; Chen, C.; Nie, Y.; Lin, X. Biofilm formation in attached microalgal reactors. Bioprocess Biosyst. Eng. 2016, 39, 1281–1288. [Google Scholar] [CrossRef]
- Katarzyna, L.; Gu, S.; Singh, O.A. Non-enclosure methods for non-suspended micro- algae cultivation: Literature review and research needs. Renew. Sustain. Energy Rev. 2015, 42, 1418–1427. [Google Scholar] [CrossRef] [Green Version]
- Hoh, D.; Watson, S.; Kan, E. Algal biofilm reactors for integrated wastewater treatment and biofuel production: A review. Chem. Eng. J. 2016, 287, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Oh, H.M.; Jo, B.H.; Lee, S.A.; Shin, S.Y.; Kim, H.S.; Lee, S.H.; Ahn, C.Y. Higher biomass productivity of microalgae in an attached growth system, using wastewater. J. Microbiol. Biotechnol. 2014, 24, 1566–1573. [Google Scholar] [CrossRef]
- Johnson, M.B.; Wen, Z. Development of an attached microalgal growth system for biofuel production. Appl. Microbiol. Biotechnol. 2010, 85, 525–534. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Hernandez, J.P.; Morey, T.; Bashan, Y. Microalgae growth-promoting bacteria as “helpers” for microalgae: A novel approach for removing ammonium and phosphorus from municipal wastewater. Water Res. 2004, 38, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.P. Wastewater treatment with suspended and non-suspended algae. J. Phycol. 1998, 34, 757–763. [Google Scholar] [CrossRef]
- Boelee, N.C.; Temmink, H.; Janssen, M.; Buisman, C.J.N.; Wijffels, R.H. Nitrogen and phosphorus removal from municipal wastewater effluent using microalgal biofilms. Water Res. 2011, 45, 5925–5933. [Google Scholar] [CrossRef] [PubMed]
- Tsolcha, O.N.; Tekerlekopoulou, A.G.; Akratos, C.S.; Bellou, S.; Aggelis, G.; Katsiapi, M.; Vayenas, D.V. Treatment of second cheese whey effluents using a Choricystis-based system with simultaneous lipid production. J. Chem. Technol. Biotechnol. 2016, 91, 2349–2359. [Google Scholar] [CrossRef]
- Tsolcha, O.N.; Tekerlekopoulou, A.G.; Akratos, C.S.; Aggelis, G.; Genitsaris, S.; Moustaka-Gouni, M.; Vayenas, D.V. Biotreatment of raisin and winery wastewaters and simultaneous biodiesel production using a Leptolyngbya-based microbial consortium. J. Clean. Prod. 2017, 148, 185–193. [Google Scholar] [CrossRef]
- Tsolcha, O.N.; Tekerlekopoulou, A.G.; Akratos, C.S.; Antonopoulou, G.; Aggelis, G.; Genitsaris, S.; Moustaka-Gouni, M.; Vayenas, D.V. A Leptolyngbya-based microbial consortium for agro-industrial wastewaters treatment and biodiesel production. Environ. Sci. Pollut. Res. 2018, 25, 17957–17966. [Google Scholar] [CrossRef]
- Economou, C.N.; Marinakis, N.; Moustaka-Gouni, M.; Kehayias, G.; Aggelis, G.; Vayenas, D.V. Lipid production by the filamentous cyanobacterium Limnothrix sp. growing in synthetic wastewater in suspended- and attached-growth photobioreactor systems. Ann. Microb. 2015, 65, 1941–1948. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998.
- Gonçalves, A.L.; Pires, J.C.M.; Simões, M. Wastewater polishing by consortia of Chlorella vulgaris and activated sludge native bacteria. J. Clean. Prod. 2016, 133, 348–357. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005.
- Folch, J.; Lees, M.; Sloane-Stanley, G.A. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Economou, C.N.; Aggelis, G.; Pavlou, S.; Vayenas, D.V. Single cell oil production from rice hulls hydrolysate. Bioresour. Technol. 2011, 102, 9737–9742. [Google Scholar] [CrossRef] [PubMed]
- Association Francaise de Normalisation. Collection of French Standards for Fats, Oleaginous Grains and Derived Products, 3rd ed.; French Association for Standardization, Paris (AFNOR): Paris, France, 1984; p. 95.
- Choi, H.J.; Lee, S.M. Effect of the N/P ratio on biomass productivity and nutrient removal from municipal wastewater. Bioprocess Biosyst. Eng. 2015, 38, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Shashirekha, V.; Sivakumar, M.; Seshadri, S. Effective C–N–P ratio for growth and nutrient removal efficiency of Scenedesmus obliquus in sugar mill effluent. Energy Ecol. Environ. 2016, 1, 283–295. [Google Scholar] [CrossRef]
- Geider, R.; La Roche, J. Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis. Eur. J. Phycol. 2002, 37, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Khemka, A.; Saraf, M. Phycoremediation of dairy wastewater coupled with biomass production using Leptolyngbya sp. J. Environ. Sci. Water Res. 2015, 4, 104–111. [Google Scholar]
- Cho, H.U.; Park, J.M.; Kim, Y.M. Enhanced microalgal biomass and lipid production from a consortium of indigenous microalgae and bacteria present in municipal wastewater under gradually mixotrophic culture conditions. Bioresour. Technol. 2017, 228, 290–297. [Google Scholar] [CrossRef]
- Solmaz, A.; Işık, M. Polishing the secondary effluent and biomass production by microalgae submerged membrane photo bioreactor. Sustain. Energy Technol. Assess. 2019, 34, 1–8. [Google Scholar] [CrossRef]
- Van Den Hende, S.; Carré, E.; Cocaud, E.; Beelen, V.; Boon, N.; Vervaeren, H. Treatment of industrial wastewaters by microalgal bacterial flocs in sequencing batch reactors. Bioresour. Technol. 2014, 161, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhou, W.; Fu, Z.; Cheng, Y.; Min, M.; Liu, Y.; Zhang, Y.; Chen, P.; Ruan, R. Effect of wastewater-borne bacteria on algal growth and nutrients removal in wastewater-based algae cultivation system. Bioresour. Technol. 2014, 167, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Lee, S.A.; Ko, S.R.; Oh, H.M.; Ahn, C.Y. Effects of photoperiod on nutrient removal, biomass production, and algal-bacterial population dynamics in lab-scale photobioreactors treating municipal wastewater. Water Res. 2015, 68, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Ansari, F.A.; Shriwastav, A.; Sahoo, N.K.; Rawat, I.; Bux, F. Dual role of Chlorella sorokiniana and Scenedesmus obliquus for comprehensive wastewater treatment and biomass production for bio-fuels. J. Clean. Prod. 2016, 115, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Tricolici, O.; Bumbac, C.; Postolache, C. Microalgae—Bacteria System for Biological Wastewater Treatment. J. Environ. Prot. Ecol. 2015, 15, 268–276. [Google Scholar]
- Su, Y.; Mennerich, A.; Urban, B. Municipal wastewater treatment and biomass accumulation with a wastewater-born and settleable algal-bacterial culture. Water Res. 2011, 45, 3351–3358. [Google Scholar] [CrossRef]
- Singh, J.; Tripathi, R.; Thakur, I.S. Characterization of endolithic cyanobacterial strain, Leptolyngbya sp. ISTCY101, for prospective recycling of CO2 and biodiesel production. Bioresour. Technol. 2014, 166, 345–352. [Google Scholar] [CrossRef]
- Singh, J.; Thakur, I.S. Evaluation of cyanobacterial endolith Leptolyngbya sp. ISTCY101, for integrated wastewater treatment and biodiesel production: A toxicological perspective. Algal Res. 2015, 11, 294–303. [Google Scholar] [CrossRef]
- Gutzeit, G.; Lorch, D.; Weber, A.; Engels, M.; Neis, U. Bioflocculent algal–bacterial biomass improves low-cost wastewater treatment. Water Sci. Technol. 2005, 52, 9–18. [Google Scholar] [CrossRef]
- Zamalloa, C.; Boon, N.; Verstraete, W. Bioresource Technology Decentralized two-stage sewage treatment by chemical—Biological flocculation combined with microalgae biofilm for nutrient immobilization in a roof installed parallel plate reactor. Bioresour. Technol. 2013, 130, 152–160. [Google Scholar] [CrossRef]
- González, C.; Marciniak, J.; Villaverde, S.; León, C.; García, P.A.; Muñoz, R. Efficient nutrient removal from swine manure in a tubular biofilm photo-bioreactor using algae-bacteria consortia. Water Sci. Technol. 2008, 58, 95–102. [Google Scholar] [CrossRef]
- Rai, J.; Kumar, D.; Pandey, L.K.; Yadav, A.; Gaur, J.P. Potential of cyanobacterial biofilms in phosphate removal and biomass production. J. Environ. Manag. 2016, 177, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Posadas, E.; García-Encina, P.; Soltau, A.; Domínguez, A.; Díaz, I.; Muñoz, R. Carbon and nutrient removal from centrates and domestic wastewater using algal—Bacterial biofilm bioreactors. Bioresour. Technol. 2013, 139, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Adey, W.H.; Kangas, P.C.; Mulbry, W. Algal Turf Scrubbing: Cleaning Surface Waters with Solar Energy while Producing a Biofuel. BioScience 2011, 61, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Travieso, L.; Benítez, F.; Sánchez, E.; Borja, R.; Colmenarejo, M.F. Production of Biomass (Algae-Bacteria) by Using a Mixture of Settled Swine and Sewage as Substrate. J. Environ. Sci. Health Part A Tox. Hazard Subst. Environ. Eng. 2006, 41, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, C.; Yang, Z.H.; Zeng, G.M.; Feng, L.J.; Liu, J.; Liu, M.; Cai, H. Continuous microalgae cultivation in aquaculture wastewater by a membrane photobioreactor for biomass production and nutrients removal. Ecol. Eng. 2016, 92, 55–61. [Google Scholar] [CrossRef]
- Anbalagan, A.; Toledo-Cervantes, A.; Posadas, E.; Rojo, E.M.; Lebrero, R.; González-Sánchez, A.; Nehrenheim, E.; Muñoz, R. Continuous photosynthetic abatement of CO2 and volatile organic compounds from exhaust gas coupled to wastewater treatment: Evaluation of tubular algal-bacterial photobioreactor. J. CO2 Util. 2017, 21, 353–359. [Google Scholar] [CrossRef]
- Viruela, A.; Robles, Á.; Durán, F.; Ruano, M.V.; Barat, R.; Ferrer, J.; Seco, A. Performance of an outdoor membrane photobioreactor for resource recovery from anaerobically treated sewage. J. Clean. Prod. 2018, 178, 665–674. [Google Scholar] [CrossRef]
- Van Den Hende, S.; Beelen, V.; Julien, L.; Lefoulon, A.; Vanhoucke, T.; Coolsaet, C.; Sonnenholzner, S.; Vervaeren, H.; Rousseau, D.P.L. Technical potential of microalgal bacterial floc raceway ponds treating food-industry effluents while producing microalgal bacterial biomass: An outdoor pilot-scale study. Bioresour. Technol. 2016, 218, 969–979. [Google Scholar] [CrossRef]
- Ferrero, E.M.; de Godos, I.; Rodríguez, E.M.; García-Encina, P.A.; Muñoz, R.; Bécares, E. Molecular characterization of bacterial communities in algal-bacterial photobioreactors treating piggery wastewaters. Ecol. Eng. 2012, 40, 121–130. [Google Scholar] [CrossRef]
- De Godos, I.; González, C.; Becares, E.; García-Encina, P.A.; Muñoz, R. Simultaneous nutrients and carbon removal during pretreated swine slurry degradation in a tubular biofilm photobioreactor. Appl. Microbiol. Biotechnol. 2009, 82, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Mennerich, A.; Urban, B. Synergistic cooperation between wastewater-born algae and activated sludge for wastewater treatment: Influence of algae and sludge inoculation ratios. Bioresour. Technol. 2012, 105, 67–73. [Google Scholar] [CrossRef]
- Christenson, L.B.; Sims, R.C. Rotating Algal Biofilm Reactor and Spool Harvester for Wastewater Treatment with. Biotechnol. Bioeng. 2012, 109, 1674–1684. [Google Scholar] [CrossRef] [PubMed]
- Sebestyén, P.; Blanken, W.; Bacsa, I.; Tóth, G.; Martin, A.; Bhaiji, T.; Dergez, A.; Kesseru, P.; Koos, A.; Kiss, I. Upscale of a laboratory rotating disk biofilm reactor and evaluation of its performance over a half-year operation period in outdoor conditions. Algal Res. 2016, 18, 266–272. [Google Scholar] [CrossRef]
- Boelee, N.C.; Janssen, M.; Temmink, H.; Taparavičiute, L.; Khiewwijit, R.; Jánoska, Á.; Busiman, C.J.N.; Wijffels, R.H. The effect of harvesting on biomass production and nutrient removal in phototrophic biofilm reactors for effluent polishing. J. Appl. Phycol. 2014, 26, 1439–1452. [Google Scholar] [CrossRef]
- Gross, M.; Henry, W.; Michael, C.; Wen, Z. Development of a rotating algal biofilm growth system for attached microalgae growth with in situ biomass harvest. Bioresour. Technol. 2013, 150, 195–201. [Google Scholar] [CrossRef]
- Derakhshan, Z.; Mahvi, A.H.; Ehrampoush, M.H.; Mazloomi, S.M.; Faramarzian, M.; Dehghani, M.; Yousefinejad, S.; Ghaneian, M.T.; Abtahi, S.M. Studies on influence of process parameters on simultaneous biodegradation of atrazine and nutrients in aquatic environments by a membrane photobioreactor. Environ. Res. 2018, 161, 599–608. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Xue, G. Algal-based immobilization process to treat the effluent from a secondary wastewater treatment plant (WWTP). J. Hazard. Mater. 2010, 178, 895–899. [Google Scholar] [CrossRef]
- Wei, Q.; Hu, Z.; Li, G.; Xiao, B.; Sun, H.; Tao, M. Removing nitrogen and phosphorus from simulated wastewater using algal biofilm technique. Front. Environ. Sci. Eng. China 2008, 2, 446–451. [Google Scholar] [CrossRef]
- Orfanos, A.G.; Manariotis, I.D. Algal biofilm ponds for polishing secondary effluent and resource recovery. J. Appl. Phycol. 2019, 31, 1765–1772. [Google Scholar] [CrossRef]
- Shayan, S.I.; Agblevor, F.A.; Bertin, L.; Sims, R.C. Hydraulic retention time effects on wastewater nutrient removal and bioproduct production via rotating algal biofilm reactor. Bioresour. Technol. 2016, 211, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Podola, B.; Melkonian, M. Application of a prototype-scale twin-layer photobioreactor for effective N and P removal from different process stages of municipal wastewater by immobilized microalgae. Bioresour. Technol. 2014, 154, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Wahidin, S.; Idris, A.; Raehanah, S.; Shaleh, M. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, N.; Tredici, M.R. Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Talebi, A.F.; Tabatabaei, M.; Chisti, Y. BiodieselAnalyzer©: A user-friendly software for predicting the properties of prospective biodiesel. Biofuel Res. J. 2015, 1, 55–57. [Google Scholar] [CrossRef]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Pollutant Concentrations (mg L−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Experimental Substrate | d-COD | NO3−-N | Total Nitrogen | PO43− | Total Sugars | Total Biomass | N:P | C:N |
| LP-S1 a | 2005.5 ± 95.5 | 26.9 ± 0.63 | 122.5 ± 0.7 | 20.8 ± 4.1 | 101.6 ± 0.1 | 243.7 ± 10.6 | 3.52 | 16.4 |
| LP-S2 a | 1145.0 ± 133.2 | 13.7 ± 0.5 | 73.2 ± 5.4 | 12.7 ± 0.3 | 46.3 ± 1.5 | 202.5 ± 0.7 | 5.76 | 15.6 |
| LP-S3 a | 491.6 ± 59.4 | 20.9 ± 0.2 | 127.8 ± 2.4 | 13.3 ± 0.7 | 28.1 ± 3.6 | 129.0 ± 7.1 | 9.60 | 3.85 |
| LC-S b | N/D i | 68.0 ± 3.1 | 68.1± 1.4 | 11.7 ± 2.3 | 4.9 ± 2.9 | 70.0 ± 22.6 | 5.82 | - |
| LP-A1 c | 2287.0 ± 122.0 | 63.7 ± 3.3 | 101.3 ± 10.5 | 21.1 ± 1.1 | 108.1 ± 6.1 | 480.0 ± 40.5 | 4.80 | 22.8 |
| LP-A2 c | 1177.0 ± 22.5 | 22.6 ± 0.4 | 92.2 ± 8.6 | 15.5 ± 2.1 | 45.0 ± 2.2 | 246.4 ± 83.8 | 5.95 | 12.8 |
| LP-A3 c | 400.0 ± 38.0 | 6.8 ± 0.5 | 38.4 ± 5.5 | 7.2 ± 0.2 | 23.1 ± 2.4 | 365.0 ± 20.9 | 5.33 | 10.42 |
| LC-A d | N/D i | 99.6 ± 2.7 | 100.8 | 17.8 ± 0.3 | 3.4 ± 0.1 | 73.5 ± 42.0 | 5.66 | - |
| PP-S1 e | 3232.5 ± 226.0 | 41.7 ± 4.4 | 48.7 ± 2.2 | 43.6 ± 9.5 | 111.8 ± 3.8 | 538.0 ± 26.5 | 1.11 | 66.4 |
| PP-S2 e | 1873.3 ± 10.5 | 35.2 ± 0.9 | 42.5 ± 0.5 | 28.2 ± 5.5 | 86.1 ± 5.2 | 374.0 ± 67.0 | 1.51 | 44.1 |
| PP-S3 e | 1217.0 ± 162.0 | 27.2 ± 0.1 | 48.5 ± 2.3 | 18.4 ± 0.1 | 79.3 ± 3.8 | 442.0 ± 67.0 | 2.63 | 25.1 |
| PP-S4 e | 638.8 ± 25.2 | 20.1 ± 1.9 | 21.7 ± 4.6 | 15.3 ± 0.1 | 39.0 ± 1.1 | 259.0 ± 24.0 | 1.42 | 29.4 |
| PC-S f | N/D i | 67.0 ± 4.1 | 67.1 ± 0.0 | 42.1 ± 1.5 | 30.05 ± 0.1 | 88.0 ± 56.5 | 1.60 | - |
| PP-A1 g | 3229.3 ± 197.0 | 75.3 ± 1.6 | 152.4 ± 1.1 | 53.6 ± 7.1 | 167.5 ± 4.8 | 446.3 ± 36.0 | 2.84 | 21.2 |
| PP-A2 g | 2032.0 ± 78.8 | 55.5 ± 5.1 | 199.0 ± 5.3 | 49.3 ± 3.0 | 229.4 ± 0.6 | 454.0 ± 48.0 | 4.04 | 10.2 |
| PP-A3 g | 1343.0 ± 44.0 | 30.6 ± 1.2 | 65.9 ± 1.2 | 31.6 ± 5.4 | 167.1 ± 13.7 | 316.0 ± 62.2 | 2.08 | 20.4 |
| PC-A h | N/D i | 75.8 ± 0.1 | 75.85 ± 0.0 | 40.0 ± 2.0 | 7.5 ± 0.1 | 98.0 ± 16.0 | 1.90 | - |
| Experimental Set | % Removal Rate | % Maximum Lipid Content | Biomass Productivity | Specific Growth Rate (d−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NO3−-N | Total Nitrogen | PO43− | Total Sugars | d-COD | Total | Attached | Total mg L−1 d−1 | Attached g m−2d−1 | ||
| LP-S1 a | 42.2 ± 0.7 | 74.0 ± 3.4 | 42.0 ± 2.1 | 48.0 ± 6.0 | 66.1 ± 3.5 | 8.3 ± 1.0 | - | 153.7 | - | 0.260 |
| LP-S2 a | 45.5 ± 0.5 | 51.2 ± 5.0 | 85.0 ± 0.5 | 31.5 ± 3.0 | 68.0 ± 7.8 | 10.3 ± 1.5 | - | 198.9 | - | 0.206 |
| LP-S3 a | 47.5 ± 0.2 | 80.5 ± 3.3 | 88.8 ± 0.4 | 7.0 ± 2.5 | 58.0 ± 5.0 | 15.5 ± 3.5 | - | 111.0 | - | 0.184 |
| LC-S b | 75.2 ± 3.0 | N/D i | 94.5 ± 1.2 | - | N/D i | 16.6 ± 1.2 | - | 47.2 | - | 0.111 |
| LP-A1 c | 82.5 ± 3.3 | 88.2 ± 9.0 | 83.0 ± 2.0 | 79.3 ± 1.2 | 94.0 ± 1.3 | 14.8 ± 2.5 | 17.7 ± 3.3 | 152.0 | 5.2 | 0.187 |
| LP-A2 c | 74.3 ± 0.4 | 82.3 ± 4.2 | 97.4 ± 1.1 | 65.1 ± 1.8 | 77.0 ± 19.5 | 9.8 ± 2.4 | 14.3 ± 1.2 | 169.6 | 2.9 | 0.153 |
| LP-A3 c | 35.0 ± 5.6 | 79.8 ± 5.0 | 91.0 ± 0.1 | 17.3 ± 2.2 | 64.5 ± 0.4 | 19.6 ± 0.1 | 18.4 ± 2.9 | 109.5 | 4.1 | 0.140 |
| LC-A d | 93.7 ± 1.4 | N/D i | 95.0 ± 0.4 | - | N/D i | 15.1 ± 4.0 | 15.9 ± 2.2 | 119.0 | 1.6 | 0.246 |
| PP-S1 e | 46.3 ± 2.2 | 54.0 ± 1.2 | 62.3 ± 4.8 | 73.8 ± 2.6 | 88.4 ± 3.7 | 6.5 ± 0.07 | - | 157.3 | - | 0.287 |
| PP-S2 e | 54.2 ± 0.5 | 61.6 ± 2.8 | 67.8 ± 3.2 | 59.1 ± 3.6 | 88.4 ± 12.8 | 8.6 ± 1.1 | - | 339.4 | - | 0.291 |
| PP-S3 e | 53.0 ± 0.2 | 59.2 ± 3.1 | 63.5 ± 0.1 | 67.7 ± 1.8 | 74.2 ± 14.0 | 8.0 ± 2.5 | - | 210.6 | - | 0.203 |
| PP-S4 e | 44.0 ± 1.23 | 36.4 ± 0.1 | 51.0 ± 0.2 | 65.4 ± 1.7 | 74.0 ± 12.0 | 5.7 ± 2.2 | - | 175.7 | - | 0.224 |
| PC-S f | 59.0 ± 2.6 | N/D i | 98.8 ± 1.1 | - | N/D i | 18.2 ± 3.7 | - | 150.6 | - | 0.130 |
| PP-A1 g | 65.0 ± 1.1 | 68.5 ± 0.6 | 81.0 ± 3.8 | 80.0 ± 3.7 | 80.0 ± 11.3 | 13.0 ± 1.8 | 7.5 ± 0.1 | 227.0 | 1.5 | 0.292 |
| PP-A2 g | 55.2 ± 3.0 | 69.4 ± 0.9 | 76.8 ± 2.8 | 83.8 ± 0.5 | 82.0 ± 6.2 | 10.1 ± 0.8 | 7.3 ± 1.3 | 316.5 | 3.4 | 0.380 |
| PP-A3 g | 31.0 ± 2.3 | 63.5 ± 3.5 | 67.8 ± 2.8 | 70.2 ± 3.7 | 51.5 ± 4.4 | 9.1 ± 1.2 | 9.5 ± 0.1 | 335.3 | 2.0 | 0.232 |
| PC-A h | 67.0 ± 0.2 | N/D i | 87.6 ± 1.1 | - | N/D i | 16.1 ± 0.2 | 15.3 ± 0.3 | 112.0 | 0.5 | 0.112 |
| Experimental Set—Recirculation Rate (mL min−1 ) | N:P | C:N | Initial NO3−-N (mg L−1) | NO3−-N Removal (%) | Initial Total Nitrogen (mg L−1) | Total Nitrogen Removal (%) | Initial PO43− (mg L−1) | PO43− Removal (%) | Initial d-COD (mg L−1) | d-COD Removal (%) | Initial Total Sugars (mg L−1) | Total Sugars Removal (%) | Maximum Lipid Content (mg L−1) | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PP-800 a | 2.9 | 48.9 | 10.1 ± 0.2 | 44.5 | 24.0 ± 0.3 | 100 | 8.2 ± 0.6 | 87.4 | 1173.4 ± 226.0 | 70.0 | 22.7 ± 1.03 | 21.4 | 7.5 | 111.0 | 0.07 |
| PP-1000 b | 2.6 | 25.1 | 27.2 ± 0.1 | 53.0 | 48.5 ± 0.4 | 59.2 | 18.4 ± 0.1 | 63.5 | 1217.0 ± 162.0 | 74.2 | 79.3 ± 3.8 | 67.7 | 8.0 | 210.6 | 0.203 |
| PP-1200 c | 2.9 | 24.5 | 19.8 ± 0.1 | 65.0 | 47.9 ± 2.2 | 54.2 | 16.1 ± 0.6 | 90.0 | 1172.3 ± 57.0 | 77.4 | 63.3 ± 4.5 | 66.0 | 6.0 | 197.1 | 0.225 |
| PP1-1200 d | 1.36 | 22.5 | 28.2 ± 0.9 | 45.6 | 91.4 ± 1.6 | 83.2 | 20.7 ± 0.6 | 67.6 | 2050.0 ± 108.0 | 60.0 | 74.9 ± 0.14 | 72.7 | 7.7 | 177.0 | 0.19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patrinou, V.; Tsolcha, O.N.; Tatoulis, T.I.; Stefanidou, N.; Dourou, M.; Moustaka-Gouni, M.; Aggelis, G.; Tekerlekopoulou, A.G. Biotreatment of Poultry Waste Coupled with Biodiesel Production Using Suspended and Attached Growth Microalgal-Based Systems. Sustainability 2020, 12, 5024. https://doi.org/10.3390/su12125024
Patrinou V, Tsolcha ON, Tatoulis TI, Stefanidou N, Dourou M, Moustaka-Gouni M, Aggelis G, Tekerlekopoulou AG. Biotreatment of Poultry Waste Coupled with Biodiesel Production Using Suspended and Attached Growth Microalgal-Based Systems. Sustainability. 2020; 12(12):5024. https://doi.org/10.3390/su12125024
Chicago/Turabian StylePatrinou, Vasiliki, Olga N. Tsolcha, Triantafyllos I. Tatoulis, Natassa Stefanidou, Marianna Dourou, Maria Moustaka-Gouni, George Aggelis, and Athanasia G. Tekerlekopoulou. 2020. "Biotreatment of Poultry Waste Coupled with Biodiesel Production Using Suspended and Attached Growth Microalgal-Based Systems" Sustainability 12, no. 12: 5024. https://doi.org/10.3390/su12125024