Effect of Soil Microbiome from Church Forest in the Northwest Ethiopian Highlands on the Growth of Olea europaea and Albizia gummifera Seedlings under Glasshouse Conditions

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Field Soil Sampling
2.3. Experimental Design
2.4. Growth and Survival Data
2.5. Soil Analysis
2.6. Plant and Soil Sample Collection
2.7. Soil DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and Sequencing
2.8. Sequence Data Processing
2.9. Statistical Analysis
3. Results
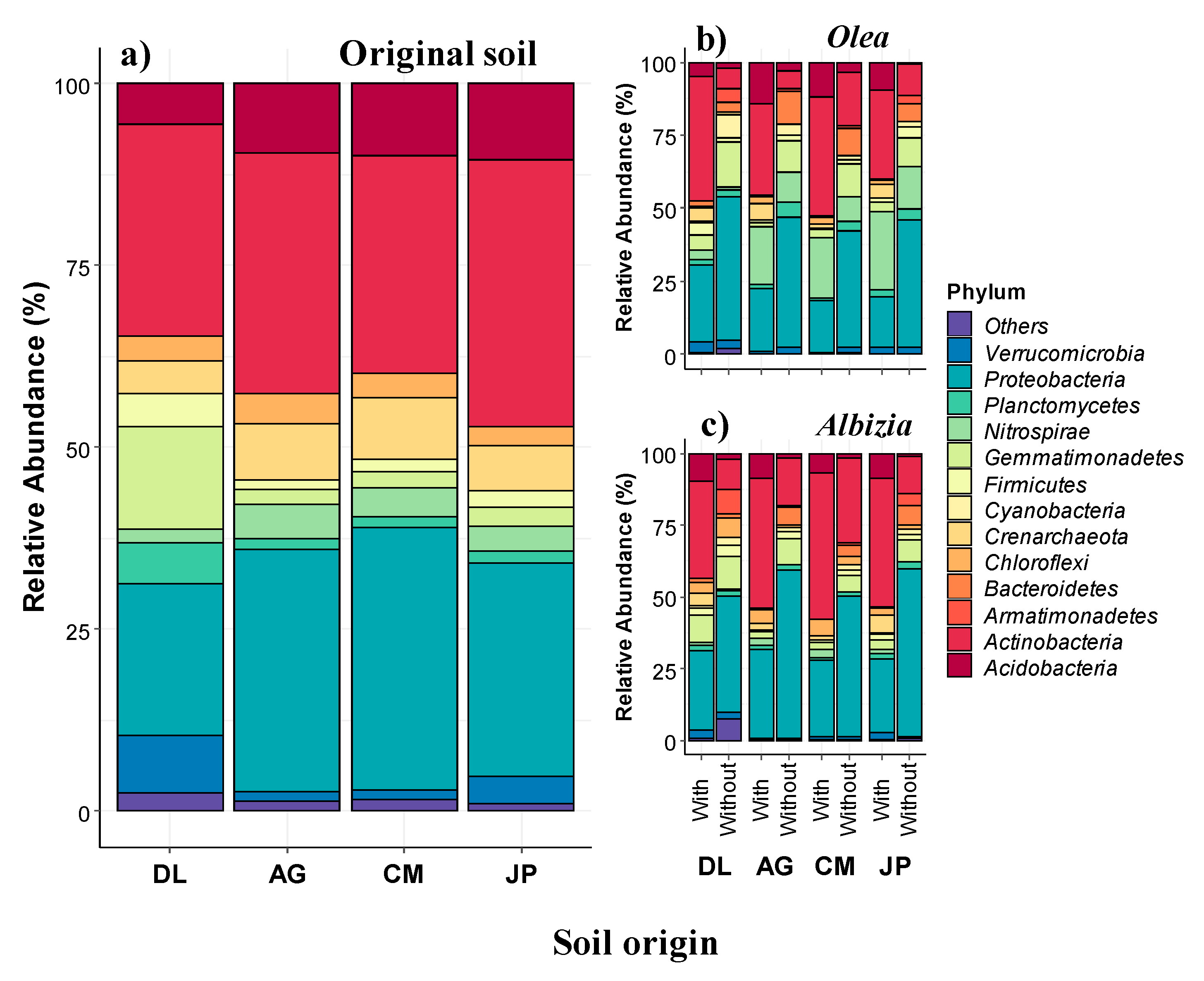
3.1. Soil Microbial Community
3.1.1. Soil Bacterial Community Composition, Abundance, and Relationship with Soil Properties
3.1.2. Soil Fungal Community Composition, Abundance, and Relationship with Soil Properties
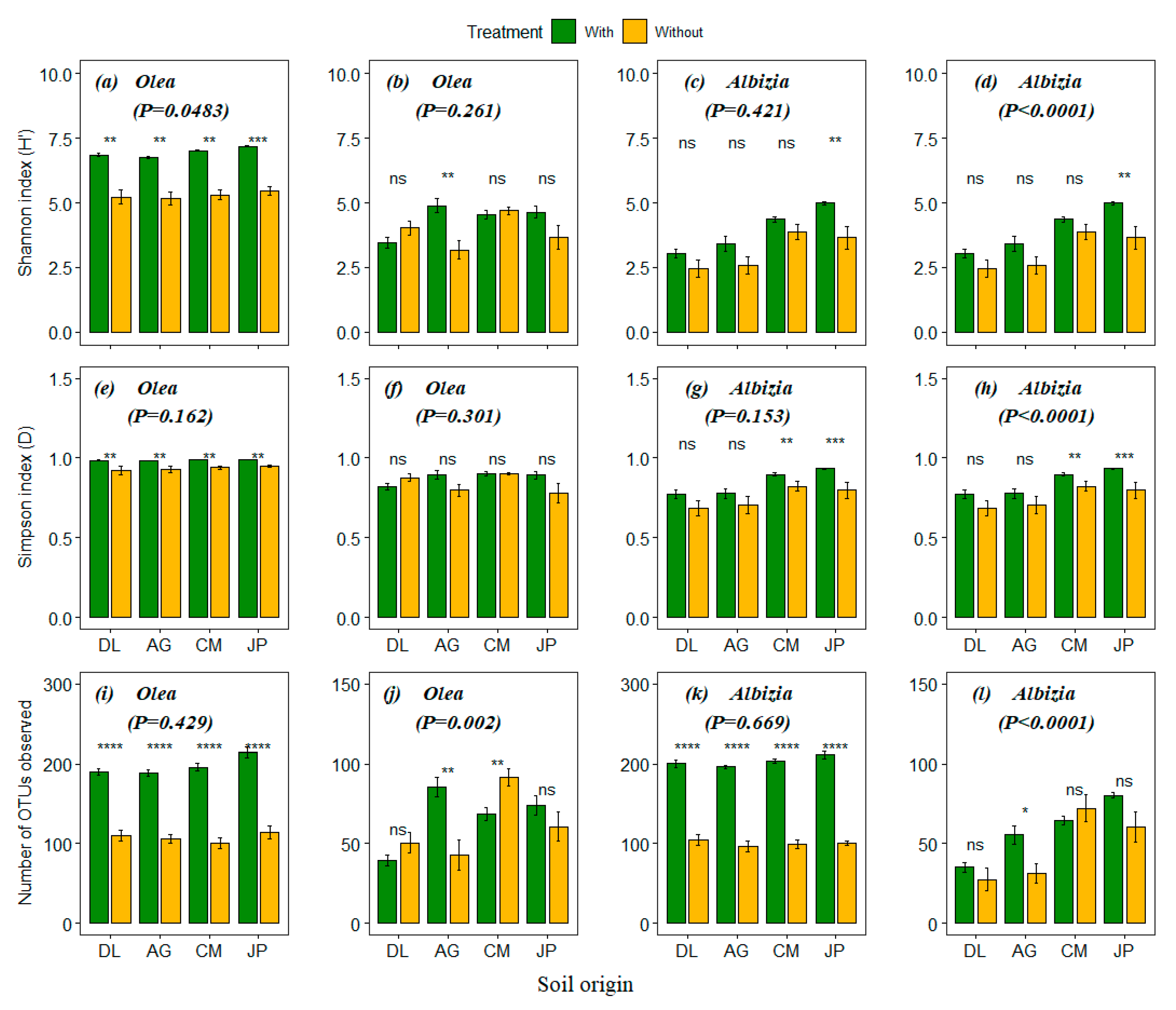
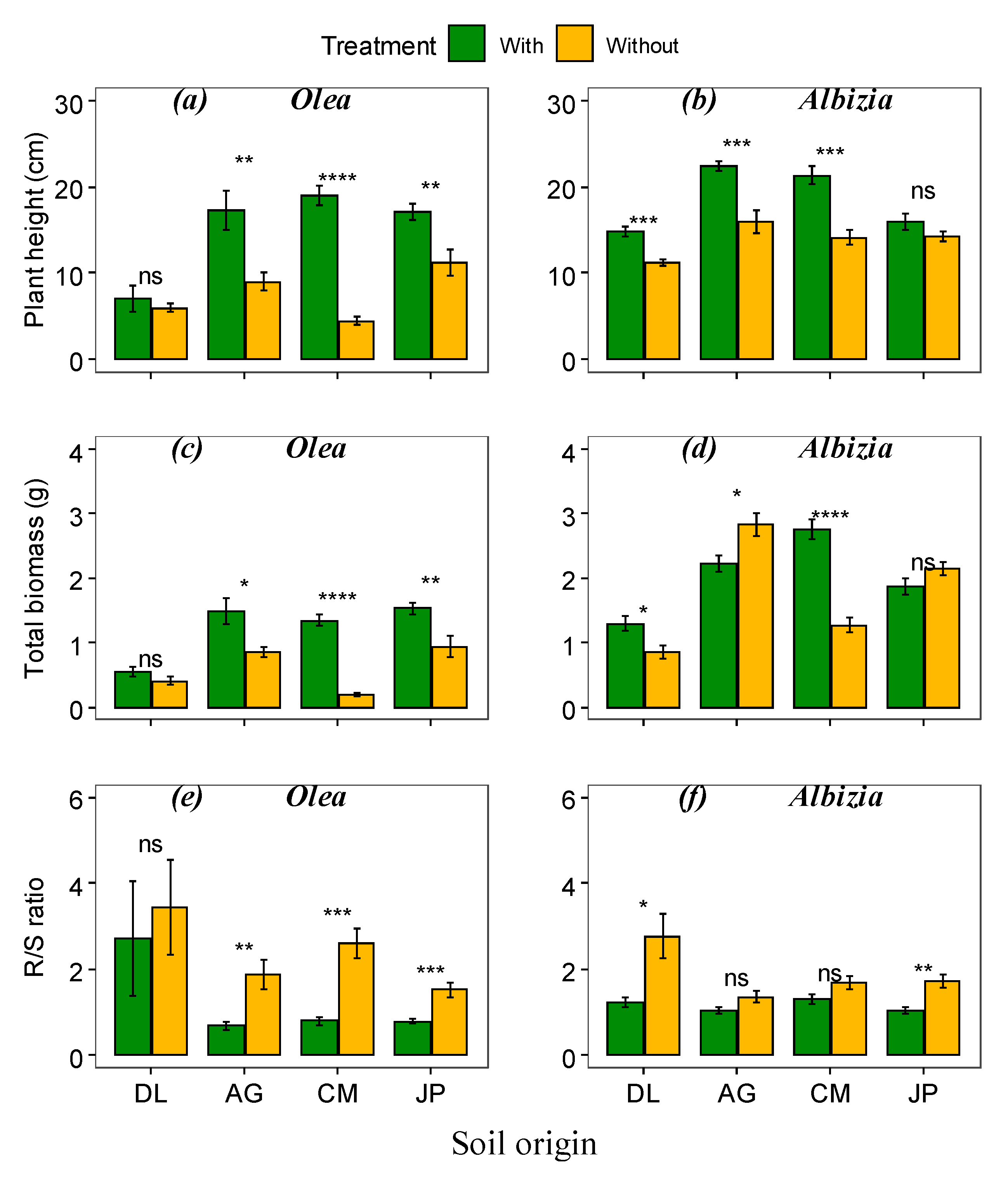
3.2. Effect of Soil Origins and Soil Treatments on Plant Growth and Survival
3.3. Soil Chemical and Physical Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nkonya, E.; Johnson, T.; Kwon, H.Y.; Kato, E. Economics of land degradation in sub-Saharan Africa. In Economics of Land Degradation and Improvement–A Global Assessment for Sustainable Development; Springer: Cham, Switzerland, 2016; pp. 215–259. [Google Scholar]
- Mekuria, W.; Wondie, M.; Amare, T.; Wubet, A.; Feyisa, T.; Yitaferu, B. Restoration of degraded landscapes for ecosystem services in North-Western Ethiopia. Heliyon 2018, 4, e00764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, J.S.; Araujo, A.S.F.; Nunes, L.; Lima, L.M.; Carneiro, R.F.V.; Salviano, A.A.C.; Tsai, S.M. Impact of Land Degradation on Soil Microbial Biomass and Activity in Northeast Brazil. Pedosphere 2012, 22, 88–95. [Google Scholar] [CrossRef]
- Meshesha, D.T.; Tsunekawa, A.; Tsubo, M.; Ali, S.A.; Haregeweyn, N. Land-use change and its socio-environmental impact in Eastern Ethiopia’s highland. Reg. Environ. Chang. 2014, 14, 757–768. [Google Scholar] [CrossRef]
- Araujo, A.S.; Borges, C.D.; Tsai, S.M.; Cesarz, S.; Eisenhauer, N. Soil bacterial diversity in degraded and restored lands of Northeast Brazil. Antonie Van Leeuwenhoek 2014, 106, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S. Microbes: The chief ecological engineers in reinstating equilibrium in degraded ecosystems. Agric. Ecosyst. Environ. 2015, 203, 80–82. [Google Scholar] [CrossRef]
- Zhang, P.; Li, L.; Pan, G.; Ren, J. Soil quality changes in land degradation as indicated by soil chemical, biochemical and microbiological properties in a karst area of southwest Guizhou, China. Environ. Geol. 2006, 51, 609–619. [Google Scholar] [CrossRef]
- Neuenkamp, L.; Prober, S.M.; Price, J.N.; Zobel, M.; Standish, R.J. Benefits of mycorrhizal inoculation to ecological restoration depend on plant functional type, restoration context and time. Fungal Ecol. 2019, 40, 140–149. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Horton, T.R. Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. J. Ecol. 2009, 97, 1139–1150. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Classen, A.T.; Sundqvist, M.K.; Henning, J.A.; Newman, G.S.; Moore, J.A.M.; Cregger, M.A.; Moorhead, L.C.; Patterson, C.M. Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 2015, 6, 21. [Google Scholar] [CrossRef]
- Wehner, J.; Antunes, P.M.; Powell, J.R.; Mazukatow, J.; Rillig, M.C. Plant pathogen protection by arbuscular mycorrhizas: A role for fungal diversity? Pedobiologia 2010, 53, 197–201. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Bergmann, J.; Verbruggen, E.; Veresoglou, S.D.; Lehmann, A. Plant root and mycorrhizal fungal traits for understanding soil aggregation. New Phytol. 2015, 205, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Calderon, K.; Spor, A.; Breuil, M.C.; Bru, D.; Bizouard, F.; Violle, C.; Barnard, R.L.; Philippot, L. Effectiveness of ecological rescue for altered soil microbial communities and functions. ISME J. 2017, 11, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosini, A.; de Souza, R.; Passaglia, L.M.P. Ecological role of bacterial inoculants and their potential impact on soil microbial diversity. Plant Soil 2015, 400, 193–207. [Google Scholar] [CrossRef]
- SER. The SER international primer on ecological restoration. Ecol. Restor 2004, 2, 206–207. [Google Scholar]
- Sultan, D.; Tsunekawa, A.; Haregeweyn, N.; Adgo, E.; Tsubo, M.; Meshesha, D.T.; Masunaga, T.; Aklog, D.; Fenta, A.A.; Ebabu, K. Impact of Soil and Water Conservation Interventions on Watershed Runoff Response in a Tropical Humid Highland of Ethiopia. Environ. Manag. 2018, 61, 860–874. [Google Scholar] [CrossRef]
- Damene, S.; Tamene, L.; Vlek, P.L.G. Performance of exclosure in restoring soil fertility: A case of Gubalafto district in North Wello Zone, northern highlands of Ethiopia. CATENA 2013, 101, 136–142. [Google Scholar] [CrossRef]
- Aerts, R.; Negussie, A.; Maes, W.; November, E.; Hermy, M.; Muys, B. Restoration of dry Afromontane forest using pioneer shrubs as nurse-plants for Olea europaea ssp. cuspidata. Restor. Ecol. 2007, 15, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Delelegn, Y.T.; Purahong, W.; Sanden, H.; Yitaferu, B.; Godbold, D.L.; Wubet, T. Transition of Ethiopian highland forests to agriculture-dominated landscapes shifts the soil microbial community composition. BMC Ecol. 2018, 18, 58. [Google Scholar] [CrossRef] [Green Version]
- Wassie, A.; Sterck, F.J.; Bongers, F. Species and structural diversity of church forests in a fragmented Ethiopian Highland landscape. J. Veg. Sci. 2010, 21, 938–948. [Google Scholar] [CrossRef]
- Abiyu, A.; Teketay, D.; Glatzel, G.; Gratzer, G. Seed production, seed dispersal and seedling establishment of two afromontane tree species in and around a church forest: Implications for forest restoration. For. Ecosyst. 2016, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Berhane, A.; Totland, Ø.; Moe, S.R. Woody plant assemblages in isolated forest patches in a semiarid agricultural matrix. Biodivers. Conserv. 2013, 22, 2519–2535. [Google Scholar] [CrossRef]
- Aerts, R.; Overtveld, K.; November, E.; Wassie, A.; Abiyu, A.; Demissew, S.; Daye, D.D.; Giday, K.; Haile, M.; TewoldeBerhan, S.; et al. Conservation of the Ethiopian church forests: Threats, opportunities and implications for their management. Sci. Total Environ. 2016, 551. [Google Scholar] [CrossRef] [Green Version]
- Wassie, A.; Sterck, F.J.; Teketay, D.; Bongers, F. Tree Regeneration in Church Forests of Ethiopia: Effects of Microsites and Management. Biotropica 2009, 41, 110–119. [Google Scholar] [CrossRef]
- Wassie, A.; Sterck, F.J.; Teketay, D.; Bongers, F. Effects of livestock exclusion on tree regeneration in church forests of Ethiopia. For. Ecol. Manag. 2009, 257, 765–772. [Google Scholar] [CrossRef]
- Friis, I. Forests and Forest Trees of Northeast Tropical Africa: Their Natural Habitats and Distribution Patterns in Ethiopia, Djibouti and Somalia; Her Majesty’s Stationery Office: London, UK, 1992. [Google Scholar]
- Tesemma, A.B. Useful Trees and Shrubs for Ethiopia: Identification, Propagation and Management for 17 Agro-Climatic Zones; Regional Land Management Unit.: Nairobi, Kenya, 2007. [Google Scholar]
- Aynekulu, E.; Denich, M.; Tsegaye, D.; Aerts, R.; Neuwirth, B.; Boehmer, H.J. Dieback affects forest structure in a dry Afromontane forest in northern Ethiopia. J. Arid Environ. 2011, 75, 499–503. [Google Scholar] [CrossRef]
- Yibeltal, M.; Tsunekawa, A.; Haregeweyn, N.; Adgo, E.; Meshesha, D.T.; Aklog, D.; Masunaga, T.; Tsubo, M.; Billi, P.; Vanmaercke, M.; et al. Analysis of long-term gully dynamics in different agro-ecology settings. Catena 2019, 179, 160–174. [Google Scholar] [CrossRef]
- Cardelús, C.; Scull, P.; Hair, J.; Baimas-George, M.; Lowman, M.; Eshete, A. A Preliminary Assessment of Ethiopian Sacred Grove Status at the Landscape and Ecosystem Scales. Diversity 2013, 5, 320–334. [Google Scholar] [CrossRef]
- Mekonnen, G. Soil Characterization Classification and Mapping of Three Twin Watersheds in the Upper Blue Nile basin, Ethiopia; Final Project Report; Amhara Design and Supervision Works Enterprise: Bahir Dar, Ethiopia, 2016. [Google Scholar]
- McNamara, N.P.; Black, H.I.J.; Beresford, N.A.; Parekh, N.R. Effects of acute gamma irradiation on chemical, physical and biological properties of soils. Appl. Soil Ecol. 2003, 24, 117–132. [Google Scholar] [CrossRef]
- Yibeltal, M.; Tsunekawa, A.; Haregeweyn, N.; Adgo, E.; Meshesha, D.T.; Masunaga, T.; Tsubo, M.; Billi, P.; Ebabu, K.; Fenta, A.A.; et al. Morphological characteristics and topographic thresholds of gullies in different agro-ecological environments. Geomorphology 2019, 341, 15–27. [Google Scholar] [CrossRef]
- Orlowsky, B. iki. dataclim: Consistency, Homogeneity and SummaryStatistics of Climatological Data. R Package Version 1.0. 2014. Available online: https://rdrr.io/cran/iki.dataclim/ (accessed on 4 September 2019).
- Chandra, N.; Kumar, S. Antibiotics Producing Soil Microorganisms. In Antibiotics and Antibiotics Resistance Genes in Soils: Monitoring, Toxicity, Risk Assessment and Management; Hashmi, M.Z., Strezov, V., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–18. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Levy, G.J.; Mamedov, A.I. High-Energy-Moisture-Characteristic Aggregate Stability as a Predictor for Seal Formation. Soil Sci. Soc. Am. J. 2002, 66, 1603–1609. [Google Scholar] [CrossRef]
- Bouyoucos, G.j. Directions For Making Mechanical Analyses Of Soils By The Hydrometer Method. Soil Sci. 1936, 42, 225–230. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.; Taniguchi, T.; Shi, W.Y.; Li, G.; Yamanaka, N.; Du, S. Land-use types and soil chemical properties influence soil microbial communities in the semiarid Loess Plateau region in China. Sci. Rep. 2017, 7, 45289. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- De Mendiburu, M.F. Package ‘agricolae’. R Package, Version 2019, 1.2-1. Available online: https://tarwi.lamolina.edu.pe/~fmendiburu/ (accessed on 4 September 2019).
- Allignol, A.; Latouche, A. CRAN Task View: Survival Analysis. Version 2017-04-25. Available online: https://CRAN.R-project.org/view=Survival.2017 (accessed on 4 September 2019).
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P. vegan: Community Ecology Package. R Package Version 2.5-2. 2018. Available online: https://rdrr.io/cran/vegan/ (accessed on 4 September 2019).
- Brady, N.; Ray, W. The Nature and Properties of Soils, 15th ed.; Pearson Education: London, UK, 2017. [Google Scholar]
- Wubet, T.; Kottke, I.; Teketay, D.; Oberwinkler, F. Mycorrhizal status of indigenous trees in dry Afromontane forests of Ethiopia. For. Ecol. Manag. 2003, 179, 387–399. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, L.; Weiner, J.; Tang, J.; Chen, X. Arbuscular mycorrhizal fungi alter plant allometry and biomass-density relationships. Ann. Bot. 2011, 107, 407–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, D.; Stief, P.; Amann, R.; De Beer, D.; Schramm, A. In situ distribution and activity of nitrifying bacteria in freshwater sediment. Environ. Microbiol. 2003, 5, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chapman, S.J.; Nicol, G.W.; Yao, H. Nitrification and nitrifiers in acidic soils. Soil Biol. Biochem. 2018, 116, 290–301. [Google Scholar] [CrossRef]
- Deacon, J. Fungal Biology. Fungal Biol. 2005. [Google Scholar] [CrossRef]
- Jia, T.; Wang, R.; Fan, X.; Chai, B. A Comparative Study of Fungal Community Structure, Diversity and Richness between the Soil and the Phyllosphere of Native Grass Species in a Copper Tailings Dam in Shanxi Province, China. Appl. Sci. 2018, 8, 1297. [Google Scholar] [CrossRef] [Green Version]
- Hewelke, E.; Górska, E.B.; Gozdowski, D.; Korc, M.; Olejniczak, I.; Prędecka, A. Soil Functional Responses to Natural Ecosystem Restoration of a Pine Forest Peucedano-Pinetum after a Fire. Forests 2020, 11, 286. [Google Scholar] [CrossRef] [Green Version]
- Garau, G.; Morillas, L.; Roales, J.; Castaldi, P.; Mangia, N.P.; Spano, D.; Mereu, S. Effect of monospecific and mixed Mediterranean tree plantations on soil microbial community and biochemical functioning. Appl. Soil Ecol. 2019, 140, 78–88. [Google Scholar] [CrossRef]
- Ushio, M.; Kitayama, K.; Balser, T.C. Tree species-mediated spatial patchiness of the composition of microbial community and physicochemical properties in the topsoils of a tropical montane forest. Soil Biol. Biochem. 2010, 42, 1588–1595. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Haichar, F.E.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Jakobsen, I.; Gronlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- St-Denis, A.; Kneeshaw, D.; Bélanger, N.; Simard, S.; Laforest-Lapointe, I.; Messier, C. Species-specific responses to forest soil inoculum in planted trees in an abandoned agricultural field. Appl. Soil Ecol. 2017, 112, 1–10. [Google Scholar] [CrossRef]
- Abiyu, A.; Teketay, D.; Glatzel, G.; Aerts, R.; Gratzer, G. Restoration of degraded ecosystems in the Afromontane highlands of Ethiopia: Comparison of plantations and natural regeneration. South. For. 2017, 79, 103–108. [Google Scholar] [CrossRef]
- Asmelash, F.; Bekele, T.; Birhane, E. The Potential Role of Arbuscular Mycorrhizal Fungi in the Restoration of Degraded Lands. Front. Microbiol. 2016, 7, 1095. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Loveys, B.R.; Atkinson, L.J.; Pons, T.L. Phenotypic plasticity and growth temperature: Understanding interspecific variability. J. Exp. Bot. 2006, 57, 267–281. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, L. Pattern and control of biomass allocation across global forest ecosystems. Ecol. Evol. 2017, 7, 5493–5501. [Google Scholar] [CrossRef]
- Eziz, A.; Yan, Z.; Tian, D.; Han, W.; Tang, Z.; Fang, J. Drought effect on plant biomass allocation: A meta-analysis. Ecol. Evol. 2017, 7, 11002–11010. [Google Scholar] [CrossRef] [PubMed]
- Zandavalli, R.B.; Dillenburg, L.R.; de Souza, P.V.D. Growth responses of Araucaria angustifolia (Araucariaceae) to inoculation with the mycorrhizal fungus Glomus clarum. Appl. Soil Ecol. 2004, 25, 245–255. [Google Scholar] [CrossRef]
- Delelegn, Y.T.; Purahong, W.; Blazevic, A.; Yitaferu, B.; Wubet, T.; Göransson, H.; Godbold, D.L. Changes in land use alter soil quality and aggregate stability in the highlands of northern Ethiopia. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caravaca, F.; Lax, A.; Albaladejo, J. Aggregate stability and carbon characteristics of particle-size fractions in cultivated and forested soils of semiarid Spain. Soil Tillage Res. 2004, 78, 83–90. [Google Scholar] [CrossRef]
- Daynes, C.N.; Field, D.J.; Saleeba, J.A.; Cole, M.A.; McGee, P.A. Development and stabilisation of soil structure via interactions between organic matter, arbuscular mycorrhizal fungi and plant roots. Soil Biol. Biochem. 2013, 57, 683–694. [Google Scholar] [CrossRef]
- Obalum, S.E.; Chibuike, G.U.; Peth, S.; Ouyang, Y. Soil organic matter as sole indicator of soil degradation. Environ. Monit. Assess. 2017, 189, 176. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Fonte, S.J.; Six, J.; Schimel, J.P. Plant versus microbial controls on soil aggregate stability in a seasonally dry ecosystem. Geoderma 2016, 272, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Bearden, B. Influence of arbuscular mycorrhizal fungi on soil structure and soil water characteristics of vertisols. Plant Soil 2001, 229, 245–258. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | pH | MC (%) | SOC (%) | TN (%) | C/N | Avail. P (mg kg−1) | Sand (%) | Silt (%) | Clay (%) | Texture Class (USDA) a |
|---|---|---|---|---|---|---|---|---|---|---|
| DL | 5.6 (0.2) b | 11.7 (1.1) c | 6.0 (0.2) b | 0.8 (0.1) b | 10.2 (0.3) b | 29.4 (1.1) b | 68 | 11 | 21 | Sandy clay loam |
| AG | 6.1 (0.1) ab | 26.7 (2.0) a | 12.0 (1.2) a | 1.0 (0.1) a | 11.8 (1.8) a | 88.2 (5.9) a | 68 | 12 | 20 | Sandy loam |
| CM | 6.5 (0.2) ab | 23.3 (1.5) ab | 8.1 (1.1) ab | 0.7 (0.1) b | 11.3 (0.4) a | 39.8 (4.2) b | 64 | 17 | 19 | Sandy loam |
| JP | 7.0 (0.3) a | 17.0 (1.6) bc | 8.7 (0.4) ab | 0.8 (0.1) b | 10.8 (0.1) b | 76.3 (5.1) a | 67 | 14 | 19 | Sandy loam |
| Species | Soil Origin | pH | SOC (%) | TN (%) | C/N | Avail.P (mg kg−1) | SAS (SI, cm−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| with | without | with | without | with | without | with | without | with | without | with | without | ||
| Olea | DL | 5.9 | 5.9 | 4.5 | 7.9 | 0.4 | 0.7 | 11 | 11 | 28.4 | 30.7 | 0.02 | 0.01 |
| (0.1) cA | (0.1) cA | −1.3 | −0.8 | −0.1 | −0.1 | −0.2 | −0.1 | (0.6) d | (0.4) d | (0.0) c | (0.0) c | ||
| ns | ns | ns | * | * | * | ns | ns | ns | ns | ||||
| AG | 6.7 | 7.2 | 6.7 | 10.1 | 0.6 | 0.92 | 11.1 | 11.4 | 88 | 86.1 | 0.05 | 0.04 | |
| (0.1) aB | (0.1) aA | −1.8 | −1.5 | −0.2 | −0.1 | −0.5 | −0.2 | (3.3) a | (4.7) a | (0.0) aA | (0.0) aB | ||
| * | * | * | * | * | ns | ns | ns | ns | ns | ||||
| CM | 6.6 | 6.6 | 6.3 | 5.7 | 0.6 | 0.5 | 10.9 | 42.2 | 42.5 | 0.04 | 0.03 | ||
| (0.1) aA | (0.1) aA | −1.4 | −1.5 | −0.1 | −0.1 | −0.2 | −0.2 | (0.7) c | (0.7) c | (0.0) bA | (0.0) bB | ||
| ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ||||
| JP | 6.3 | 5.9 | 8.3 | 6.8 | 0.7 | 0.62 | 11.3 | 10.9 | 77.9 | 74.8 | 0.04 | 0.03 | |
| (0.1) bA | (0.1) bB | −1.7 | −0.7 | −0.1 | −0.1 | −0.1 | −0.1 | (2.2) b | (2.0) b | (0.0) b | (0.0) b | ||
| * | * | ns | ns | ns | ns | ns | ns | ns | ns | ||||
| Albizia | DL | 5.6 | 5.9 | 3.9 | 3.7 | 0.4 | 0.3 | 10.1 | 11.4 | 27.3 | 28.6 | 0.02 | 0.01 |
| (0.1) dB | (0.0) dA | (0.0) d | (0.0) d | (0.0) d | (0.0) c | (0.1) a | (0.2) a | (0.4) dB | (0.6) dA | (0.0) d | (0.0) c | ||
| ns | ns | * | * | * | * | ns | * | ns | ns | ||||
| AG | 7 | 7.4 | 12.1 | 11.9 | 1 | 1 | 11.6 | 11.6 | 74.1 | 86.9 | 0.05 | 0.04 | |
| (0.1) aB | (0.0) aA | (0.2) a | (0.7) a | (0.0) a | (0.1) a | (0.1) a | (0.1) a | (2.0) bB | (3.9) bA | (0.0) aA | (0.0) aB | ||
| * | * | ns | ns | ns | ns | * | * | * | ns | ||||
| CM | 6.5 | 6.6 | 8.3 | 8.6 | 0.8 | 0.8 | 10.7 | 10.8 | 34.5 | 47.3 | 0.04 | 0.04 | |
| (0.0) bA | (0.1) bA | (0.1) b | (0.4) b | (0.0) b | (0.0) b | (0.1) b | (0.1) b | (0.8) c | (1.1) c | (0.0) b | (0.0) a | ||
| ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ||||
| JP | 6 | 6.3 | 7.3 | 7.7 | 0.7 | 0.7 | 10.6 | 10.7 | 74.2 | 94.8 | 0.03 | 0.03 | |
| (0.0) cB | (0.0) cA | (0.1) c | (0.1) c | (0.0) c | (0.0) b | (0.1) b | (0.0) b | (3.7) aB | (1.5) aA | (0.0) c | (0.0) b | ||
| * | * | * | ns | ns | ns | ns | ns | ns | * | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abebe, G.; Tsunekawa, A.; Haregeweyn, N.; Taniguchi, T.; Wondie, M.; Adgo, E.; Masunaga, T.; Tsubo, M.; Ebabu, K.; Mamedov, A.; et al. Effect of Soil Microbiome from Church Forest in the Northwest Ethiopian Highlands on the Growth of Olea europaea and Albizia gummifera Seedlings under Glasshouse Conditions. Sustainability 2020, 12, 4976. https://doi.org/10.3390/su12124976
Abebe G, Tsunekawa A, Haregeweyn N, Taniguchi T, Wondie M, Adgo E, Masunaga T, Tsubo M, Ebabu K, Mamedov A, et al. Effect of Soil Microbiome from Church Forest in the Northwest Ethiopian Highlands on the Growth of Olea europaea and Albizia gummifera Seedlings under Glasshouse Conditions. Sustainability. 2020; 12(12):4976. https://doi.org/10.3390/su12124976
Chicago/Turabian StyleAbebe, Getu, Atsushi Tsunekawa, Nigussie Haregeweyn, Takeshi Taniguchi, Menale Wondie, Enyew Adgo, Tsugiyuki Masunaga, Mitsuru Tsubo, Kindiye Ebabu, Amrakh Mamedov, and et al. 2020. "Effect of Soil Microbiome from Church Forest in the Northwest Ethiopian Highlands on the Growth of Olea europaea and Albizia gummifera Seedlings under Glasshouse Conditions" Sustainability 12, no. 12: 4976. https://doi.org/10.3390/su12124976