New Soil, Old Plants, and Ubiquitous Microbes: Evaluating the Potential of Incipient Basaltic Soil to Support Native Plant Growth and Influence Belowground Soil Microbial Community Composition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Seed Information
2.3. Plant and Soil Measurements
2.4. Soil Microbial Community Analysis
2.5. Functional Annotation of Sequence Data
2.6. Data Analyses
3. Results and Discussion
3.1. Germination, Plant Height and Aboveground Biomass
3.2. Physico-Chemical Properties of the Studied Soil Materials
3.3. Composition of Microbial Communities
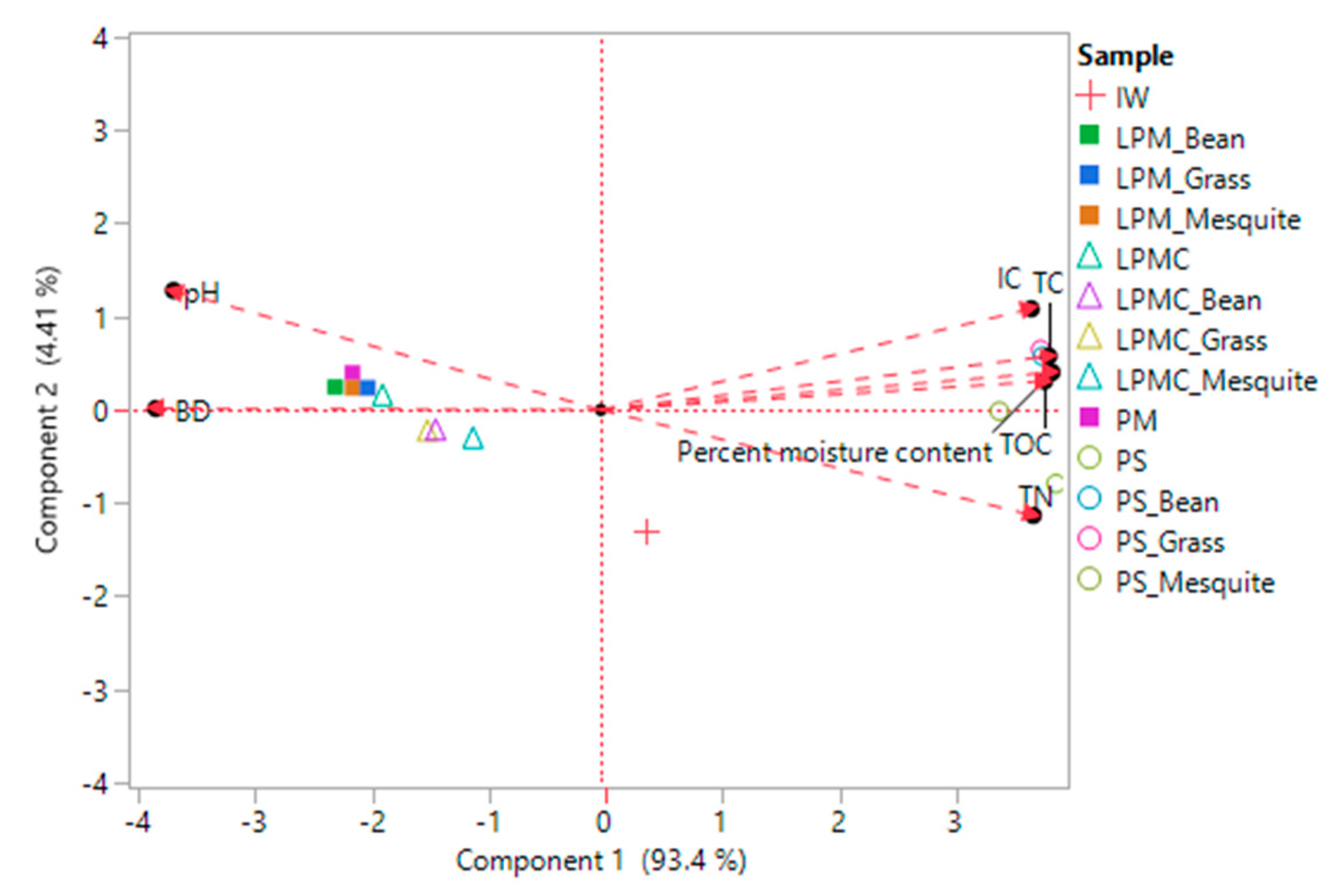
3.4. Microbial Communities and Environmental Variables
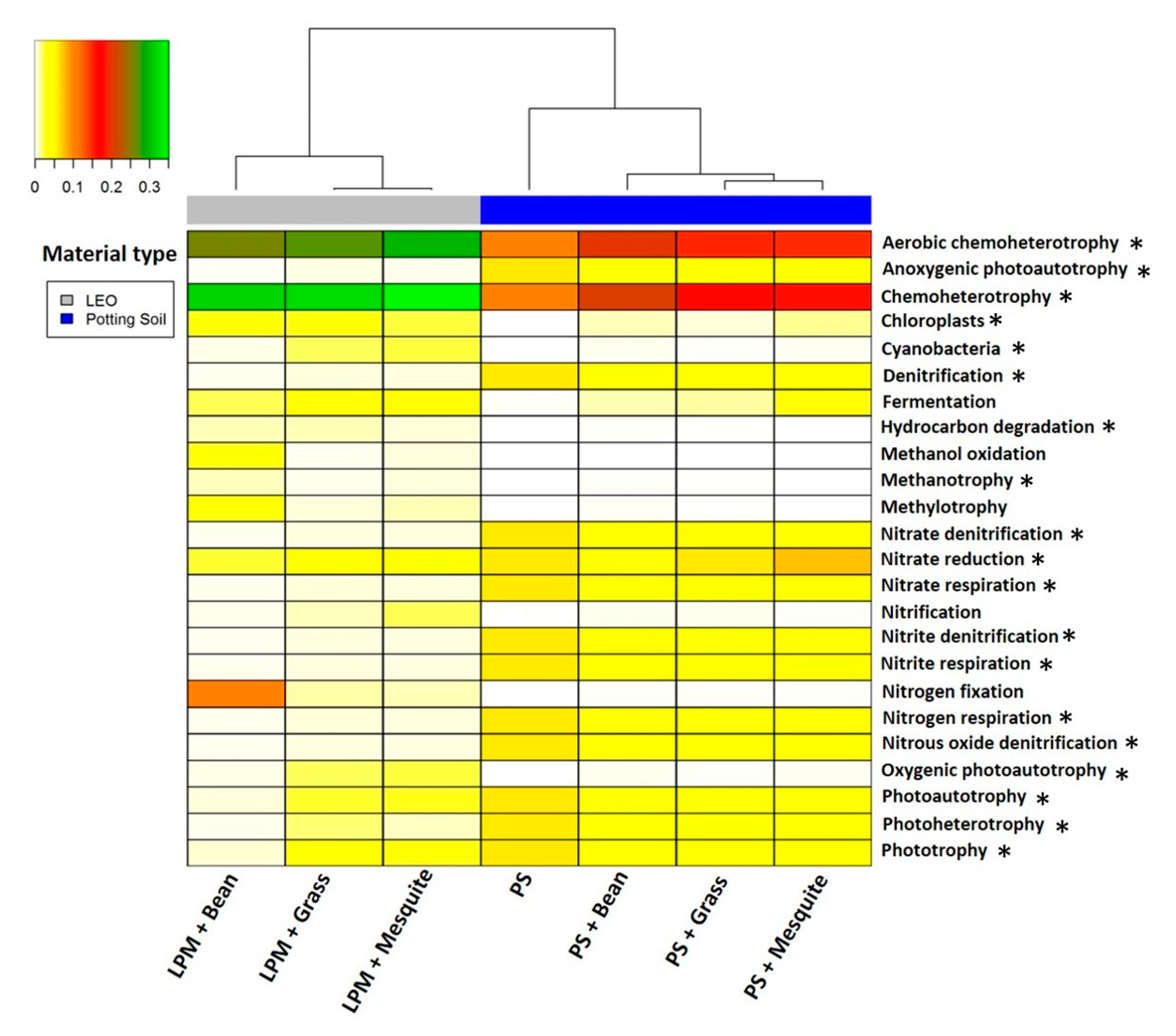
3.5. Predicted Functional Potential of Microbial Community
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils, 15th ed.; Pearson: London, UK, 2017; ISBN 0133254488. [Google Scholar]
- Pimentel, D. Soil erosion: A food and environmental threat. Environ. Dev. Sustain. 2006, 8, 119–137. [Google Scholar] [CrossRef]
- O’geen, A.T.; Schwankl, L.J. University of California Davis, Division of Agriculture and Natural Resources, Publication 8196; University of California: Davis, CA, USA, 2006. [Google Scholar]
- World Health Organization. Land Degradation and Desertification. 2012. Available online: http://www.who.int/globalchange/ecosystems/desert/en/ (accessed on 6 April 2018).
- UNCCD/Science-Policy Interface. Land in Balance the Scientific Conceptual Framework for Land Degradation Neutrality Conceptual Framework for Land Degradation Neutrality (LDN); United Nations Convention to Combat Desertification: Bonn, Germany, 2016. [Google Scholar]
- Orr, B.J.; Cowie, A.L.; Sanchez, V.M.C.; Chasek, P.; Crossman, N.D.; Erlewein, A.; Louwagie, G.; Maron, M.; Metternicht, G.I.; Minelli, S.A.E.; et al. Scientific Conceptual Framework for Land Degradation Neutrality: A Report of the Science-Policy Interface; United Nations Covention to Combat Desertification: Bonn, Germany, 2017. [Google Scholar]
- Montoya-Tangarife, C.; de la Barrera, F.; Salazar, A.; Inostroza, L. Monitoring the effects of land cover change on the supply of ecosystem services in an urban region: A study of Santiago-Valparaíso, Chile. PLoS ONE 2017, 12, e0188117. [Google Scholar] [CrossRef] [PubMed]
- Valentín-Vargas, A.; Root, R.A.; Neilson, J.W.; Chorover, J.; Maier, R.M. Environmental factors influencing the structural dynamics of soil microbial communities during assisted phytostabilization of acid-generating mine tailings: A mesocosm experiment. Sci. Total Environ. 2014, 500–501, 314–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honeker, L.K.; Neilson, J.W.; Root, R.A.; Gil-Loaiza, J.; Chorover, J.; Maier, R.M. Bacterial rhizoplane colonization patterns of buchloe dactyloides growing in metalliferous mine tailings reflect plant status and biogeochemical conditions. Microb. Ecol. 2017, 74, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Valentín-Vargas, A.; Neilson, J.W.; Root, R.A.; Chorover, J.; Maier, R.M. Treatment impacts on temporal microbial community dynamics during phytostabilization of acid-generating mine tailings in semiarid regions. Sci. Total Environ. 2018, 618, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Grandlic, C.J.; Mendez, M.O.; Chorover, J.; Machado, B.; Maier, R.M. Plant growth-promoting bacteria for phytostabilization of mine tailings. Environ. Sci. Technol. 2008, 42, 2079–2084. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.O.; Maier, R.M. Phytoremediation of mine tailings in temperate and arid environments. Rev. Environ. Sci. Biotechnol. 2008, 7, 47–59. [Google Scholar] [CrossRef]
- Smith, S.W.; Ross, K.; Karlsson, S.; Bond, B.; Upson, R.; Davey, A. Going native, going local: Revegetating eroded soils on the Falkland Islands using native seeds and farmland waste. Restor. Ecol. 2017, 26, 134–144. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef]
- Debenport, S.J.; Assigbetse, K.; Bayala, R.; Chapuis-Lardy, L.; Dick, R.P.; McSpadden Gardener, B.B. Shifting populations in the root-zone microbiome of millet associated with enhanced crop productivity in the Sahel. Appl. Environ. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tittonell, P.; Scopel, E.; Andrieu, N.; Posthumus, H.; Mapfumo, P.; Corbeels, M.; van Halsema, G.E.; Lahmar, R.; Lugandu, S.; Rakotoarisoa, J.; et al. Agroecology-based aggradation-conservation agriculture (ABACO): Targeting innovations to combat soil degradation and food insecurity in semi-arid Africa. Field Crops Res. 2012, 132, 168–174. [Google Scholar] [CrossRef]
- Abd-Elmabod, S.; Bakr, N.; Muñoz-Rojas, M.; Pereira, P.; Zhang, Z.; Cerdà, A.; Jordán, A.; Mansour, H.; De la Rosa, D.; Jones, L. Assessment of soil suitability for improvement of soil factors and agricultural management. Sustainability 2019, 11, 1588. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, R.R.; Debenport, S.J.; Leewis, M.C.C.E.; Ndoye, F.; Nkenmogne, K.I.E.; Soumare, A.; Thuita, M.; Gueye, M.; Miambi, E.; Chapuis-Lardy, L.; et al. The native shrub, Piliostigma reticulatum, as an ecological “resource island” for mango trees in the Sahel. Agric. Ecosyst. Environ. 2015, 204, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Nunes, J.S.; Araujo, A.S.F.; Nunes, L.A.P.L.; Lima, L.M.; Carniero, R.F.V.; Salviano, A.A.C.; Tsai, S.M. Impact of land degradation on soil microbial biomass and activity in Northeast Brazil. Pedosphere 2012, 22, 88–95. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Chen, S.; Qi, G.; He, Z.; Zhao, X. Microbial taxa and functional genes shift in degraded soil with bacterial wilt. Sci. Rep. 2017, 7, 39911. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.S.F.; Borges, C.D.; Tsai, S.M.; Cesarz, S.; Eisenhauer, N. Soil bacterial diversity in degraded and restored lands of Northeast Brazil. Antonie Van Leeuwenhoek 2014, 106, 891–899. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Jenny, H.K. Factors of Soil Formation: A System of Quantitative Pedology; Tata McGraw Hill: New York, NY, USA, 1941. [Google Scholar]
- Dauber, J.; Brown, C.; Fernando, A.L.; Finnan, J.; Krasuska, E.; Ponitka, J.; Styles, D.; Thrän, D.; Jan, K.; Groenigen, V.; et al. Bioenergy from “surplus” land: Environmental and socio-economic implications. BioRisk 2012, 7, 5–50. [Google Scholar] [CrossRef]
- Mendez, M.O.; Neilson, J.W.; Maier, R.M. Characterization of a bacterial community in an abandoned semiarid lead-zinc mine tailing site. Appl. Environ. Microbiol. 2008, 74, 3899–3907. [Google Scholar] [CrossRef] [Green Version]
- Ciria, C.; Sanz, M.; Carrasco, J.; Ciria, P. Identification of arable marginal lands under rainfed conditions for bioenergy purposes in Spain. Sustainability 2019, 11, 1833. [Google Scholar] [CrossRef] [Green Version]
- Volkmann, T.H.M.; Sengupta, A.; Pangle, L.A.; Dontsova, K.; Barron-Gafford, G.A.; Harman, C.J.; Niu, G.-Y.; Abramson, N.; Meira-Neto, A.A.; Wang, Y.; et al. Controlled experiments of hillslope coevolution at the biosphere 2 landscape evolution observatory: Toward prediction of coupled hydrological, biogeochemical, and ecological change. In Hydrology of Artificial and Controlled Experiments; Intech Open Limiited: London, UK, 2017; pp. 25–74. [Google Scholar]
- Sengupta, A.; Pangle, L.A.; Volkmann, T.H.M.; Dontsova, K.; Troch, P.A.; Meira-neto, A.A.; Neilson, J.W.; Hunt, E.A.; Chorover, J.; Zeng, X.; et al. Advancing understanding of hydrological and biogeochemical interactions in evolving landscapes through controlled experimentation at the landscape evolution observatory. In Terrestrial Ecosystems Research Infrastructure: Challenges and Opportunities; Chabbi, A., Loescher, H.W., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2017; pp. 83–118. [Google Scholar]
- Pangle, L.A.; DeLong, S.B.; Abramson, N.; Adams, J.; Barron-Gafford, G.A.; Breshears, D.D.; Brooks, P.D.; Chorover, J.; Dietrich, W.E.; Dontsova, K.; et al. The landscape evolution observatory: A large-scale controllable infrastructure to study coupled Earth-surface processes. Geomorphology 2015, 244, 190–203. [Google Scholar] [CrossRef] [Green Version]
- Pohlmann, M.; Dontsova, K.; Root, R.; Ruiz, J.; Troch, P.; Chorover, J. Pore water chemistry reveals gradients in mineral transformation across a model basaltic hillslope. Geochem. Geophys Geosystems 2016, 17, 2054–2069. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, A.; Stegen, J.C.; Nielson, J.W.; Meira-Neto, A.; Wang, Y.; Troch, P.A.; Chorover, J.; Maier, R.M. Assessment of microbial community patterns under incipient conditions in a basalt soil system. J. Geophys. Res. Biogeosci. 2019, 124, 941–958. [Google Scholar] [CrossRef]
- Native Seeds/SEARCH—Home. Available online: https://www.nativeseeds.org/ (accessed on 29 August 2017).
- Sengupta, A.; Wang, Y.; Meira Neto, A.A.; Matos, K.A.; Dontsova, K.; Root, R.; Neilson, J.W.; Maier, R.M.; Chorover, J.; Troch, P.A. Soil lysimeter excavation for coupled hydrological, geochemical, and microbiological investigations. J. Vis. Exp. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Jim, A.; Sengupta, A. Assessing the ability of incipient basaltic soil to support plants native to Southwestern United States. J. Undergrad. Res. 2018, IX, 30–33. [Google Scholar]
- Duke, J.A. Phaseolus vulgaris L. In Handbook of Energy Crops; Available online: https://www.hort.purdue.edu/newcrop/duke_energy/Phaseolus_vulgaris.html (accessed on 15 July 2017).
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laubitz, D.; Harrison, C.A.; Midura-Kiela, M.T.; Ramalingam, R.; Larmonier, C.B.; Chase, J.H.; Caporaso, J.G.; Besselsen, D.G.; Ghishan, F.K.; Kiela, P.R. Reduced epithelial Na+/H+ exchange drives gut microbial dysbiosis and promotes inflammatory response in T cell-mediated murine colitis. PLoS ONE 2016, 11, e0152044. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: https://www.R-project.org/ (accessed on 21 May 2020).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Warenes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. gplots: Various R Programming Tools for Plotting Data; R Package Version; 2016; Available online: https://rdrr.io/cran/gplots/ (accessed on 21 May 2020).
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan tutorial; 2015; pp. 1–40. Available online: https://www.mooreecology.com/uploads/2/4/2/1/24213970/vegantutor.pdf (accessed on 21 May 2020).
- Dray, S.; Dufor, A.-B.; Thioulouse, J. Analysis of Ecological Data: Exploratory and Euclidean Methods in Environmental Sciences. 2018. Available online: https://rdrr.io/cran/ade4/ (accessed on 21 May 2020).
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Solís-Domínguez, F.A.; Valentín-Vargas, A.; Chorover, J.; Maier, R.M. Effect of arbuscular mycorrhizal fungi on plant biomass and the rhizosphere microbial community structure of mesquite grown in acidic lead/zinc mine tailings. Sci. Total Environ. 2011, 409, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Romero, E.; Segovia, L.; Mercante, F.M.; Franco, A.A.; Graham, P.; Pardo, M.A. Rhizobium tropici, a novel species nodulating phaseolus vulgaris l. beans and leucaena sp. trees. Int. J. Syst. Bacteriol. 1991, 41, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Romero, E. Diversity of rhizobium-phaseolus vulgaris symbiosis: Overview and perspectives. Plant Soil 2003, 252, 11–23. [Google Scholar] [CrossRef]
- Gtari, M.; Ghodhbane-Gtari, F.; Nouioui, I.; Beauchemin, N.; Tisa, L.S. Phylogenetic perspectives of nitrogen-fixing actinobacteria. Arch. Microbiol. 2012, 194, 3–11. [Google Scholar] [CrossRef]
- Neilson, J.W.; Quade, J.; Ortiz, M.; Nelson, W.M.; Legatzki, A.; Tian, F.; LaComb, M.; Betancourt, J.L.; Wing, R.A.; Soderlund, C.A.; et al. Life at the hyperarid margin: Novel bacterial diversity in arid soils of the Atacama Desert, Chile. Extremophiles 2012, 16, 553–566. [Google Scholar] [CrossRef]
- Kardol, P.; De Deyn, G.B.; Laliberté, E.; Mariotte, P.; Hawkes, C.V. Biotic plant-soil feedbacks across temporal scales. J. Ecol. 2013, 101, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Kelly, L.C.; Cockell, C.S.; Herrera-Belaroussi, A.; Piceno, Y.; Andersen, G.; DeSantis, T.; Brodie, E.; Thorsteinsson, T.; Marteinsson, V.; Poly, F.; et al. Bacterial diversity of terrestrial crystalline volcanic rocks, Iceland. Microb. Ecol. 2011, 62, 69–79. [Google Scholar] [CrossRef]
- Akob, D.M.; Küsel, K. Where microorganisms meet rocks in the Earth’s Critical Zone. Biogeosciences 2011, 8, 3531–3543. [Google Scholar] [CrossRef] [Green Version]
- Knelman, J.E.; Legg, T.M.; O’Neill, S.P.; Ashenberger, C.L.W.; Gonzalez, A.G.; Cleveland, C.C.; Nemergut, D.R. Bacterial community structure and function change in association with colonizer plants during early primary succession in a glacier forefield. Soil Biol. Biochem. 2012, 46, 172–180. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PERMANOVA | Mantel Test | |||
|---|---|---|---|---|
| Environmental variables | R2 | P | r | P |
| pH | 0.46 | 0.002 | 0.91 | 0.002 |
| TC | 0.47 | 0.007 | 0.87 | 0.012 |
| TN | 0.45 | 0.02 | 0.9 | 0.002 |
| TOC | 0.47 | 0.002 | 0.85 | 0.006 |
| TIC | 0.42 | 0.02 | 0.69 | 0.023 |
| Moisture content | 0.47 | 0.018 | 0.86 | 0.039 |
| Bulk density | 0.43 | 0.03 | 0.81 | 0.017 |
| Soil material type | 0.47 | 0.035 | 0.87 | 0.034 |
| Plant attributes | R2 | P | r | P |
| Germination rate | 0.15 | 0.474 | 0.02 | 0.413 |
| Plant height | 0.21 | 0.232 | 0.10 | 0.310 |
| Wet aboveground biomass | 0.17 | 0.429 | −0.16 | 0.660 |
| Dry aboveground biomass | 0.17 | 0.416 | −0.14 | 0.605 |
| Plant type | 0.39 | 0.799 | 0.00 | 0.452 |
| Predicted Functions | F | P |
|---|---|---|
| Aerobic chemoheterotrophy | 28.6 | 0.003 |
| Anoxygenic photoautotrophy | 87.8 | 0.0002 |
| Chemoheterotrophy | 40.2 | 0.001 |
| Chloroplasts | 26.5 | 0.004 |
| Cyanobacteria | 7.6 | 0.040 |
| Denitrification | 84.1 | 0.0003 |
| Fermentation | 4.3 | 0.094 |
| Hydrocarbon degradation | 38.1 | 0.002 |
| Methanol oxidation | 2.8 | 0.157 |
| Methanotrophy | 10.7 | 0.022 |
| Methylotrophy | 3.9 | 0.106 |
| Nitrate denitrification | 84.1 | 0.0003 |
| Nitrate reduction | 12.0 | 0.018 |
| Nitrate respiration | 82.8 | 0.0003 |
| Nitrification | 4.2 | 0.096 |
| Nitrite denitrification | 84.1 | 0.0003 |
| Nitrite respiration | 84.4 | 0.0003 |
| Nitrogen fixation | 2.4 | 0.185 |
| Nitrogen respiration | 82.8 | 0.0003 |
| Nitrous oxide denitrification | 84.1 | 0.0003 |
| Oxygenic photoautotrophy | 7.6 | 0.040 |
| Photoautotrophy | 20.3 | 0.006 |
| Photoheterotrophy | 45.1 | 0.001 |
| Phototrophy | 7.9 | 0.037 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sengupta, A.; Kushwaha, P.; Jim, A.; Troch, P.A.; Maier, R. New Soil, Old Plants, and Ubiquitous Microbes: Evaluating the Potential of Incipient Basaltic Soil to Support Native Plant Growth and Influence Belowground Soil Microbial Community Composition. Sustainability 2020, 12, 4209. https://doi.org/10.3390/su12104209
Sengupta A, Kushwaha P, Jim A, Troch PA, Maier R. New Soil, Old Plants, and Ubiquitous Microbes: Evaluating the Potential of Incipient Basaltic Soil to Support Native Plant Growth and Influence Belowground Soil Microbial Community Composition. Sustainability. 2020; 12(10):4209. https://doi.org/10.3390/su12104209
Chicago/Turabian StyleSengupta, Aditi, Priyanka Kushwaha, Antonia Jim, Peter A. Troch, and Raina Maier. 2020. "New Soil, Old Plants, and Ubiquitous Microbes: Evaluating the Potential of Incipient Basaltic Soil to Support Native Plant Growth and Influence Belowground Soil Microbial Community Composition" Sustainability 12, no. 10: 4209. https://doi.org/10.3390/su12104209