
Morphological and Genetic Diversity of Scutellaria tuvensis Juz., an Endemic of Desert Steppes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Analysis
2.3. Molecular Analysis
2.4. Data Analysis
3. Results and Discussion
3.1. Morphological Diversity
3.2. Evaluation of Intrapopulation Polymorphism
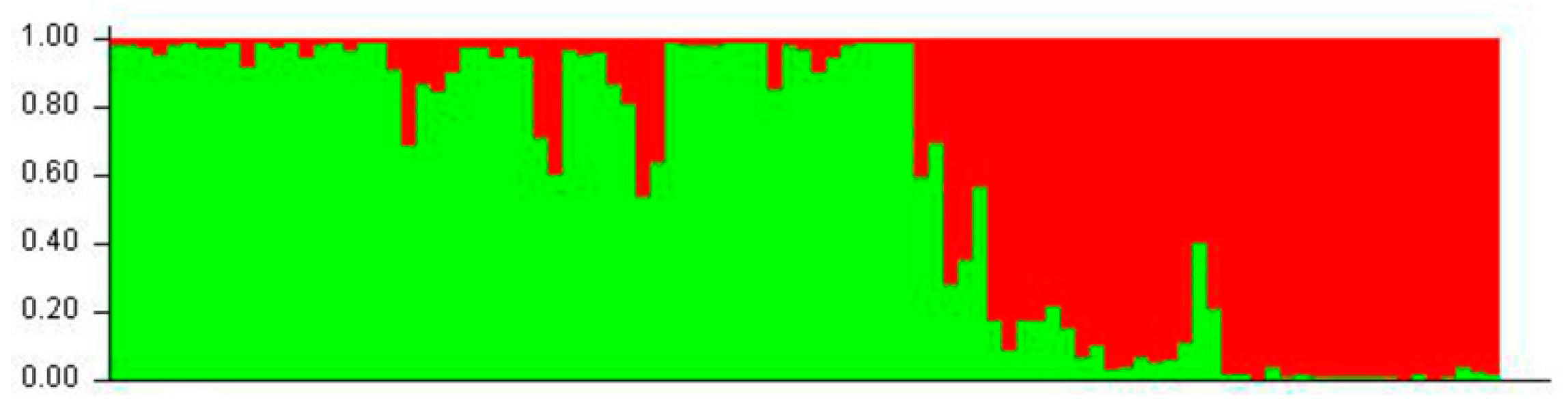
3.3. Quantitation of Interpopulation Differences
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frankel, O.H. Genetic Conservation: Our Evolutionary Responsibility. Genetics 1974, 78, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.J. The phytogeography of Scutellaria L. Notes R. Bot. Gard. 1990, 46, 345–359. [Google Scholar]
- Yuzepchuk, S.V. Genus Scutellaria L. In Flora SSSR; Komarov, V.L., Ed.; AN SSSR: Saint Petersburg, Russia, 1954; Volume 20, pp. 183–184. (In Russian) [Google Scholar]
- Kamelin, R.V.; Gubanov, I.A. Scutellaria grandiflora Sims S. L. in Mongolia. Byull. MOIP. Otd. Biol. 1989, 94, 109–111. (In Russian) [Google Scholar]
- Zuev, V.V. Scutellaria L. Flora of Siberia; Malyshev, L.I., Ed.; Nauka: Novosibirsk, Russia, 1997; Volume 11, pp. 161–165. (In Russian) [Google Scholar]
- Guseva, A.A.; Cheryomushkina, V.A. Morphogenesis and state of coenopopulations of the endemic species Scutellaria tuvensis (Lamiaceae). Byull. MOIP. Otd. Biol. 2017, 122, 68–77. (In Russian) [Google Scholar]
- Guseva, A.; Cheryomushkina, V. Polyvariation of the development Scutellaria L. species and the structure of their populations in Siberia. BIO Web Conf. 2019, 16, 00010. [Google Scholar] [CrossRef] [Green Version]
- Awad, R.; Arnason, J.; Trudeau, V.; Bergeron, C.; Budzinski, J.; Foster, B.; Merali, Z. Phytochemical and biological analysis of Skullcap (Scutellaria lateriflora L.): A medicinal plant with anxiolytic properties. Phytomedicine 2003, 10, 640–649. [Google Scholar] [CrossRef]
- Sonoda, M.; Nishiyama, T.; Matsukawa, Y.; Moriyasu, M. Cytotoxic activities of flavonoids from two Scutellaria plants in Chinese medicine. J. Ethnopharmacol. 2004, 91, 65–68. [Google Scholar] [CrossRef]
- Bardakci, H.; Skaltsa, H.; Milosevic-Ifantis, T.; Lazari, D.; Hadjipavlou-Litina, D.; Yeşilada, E.; Kırmızıbekmez, H. Antioxidant activities of several Scutellaria taxa and bioactive phytoconstituents from Scutellaria hastifolia L. Ind. Crops Prod. 2015, 77, 196–203. [Google Scholar] [CrossRef]
- Han, Q.-T.; Ren, Y.; Li, G.-S.; Xiang, K.-L.; Dai, S.-J. Flavonoid alkaloids from Scutellaria moniliorrhiza with anti-inflammatory activities and inhibitory activities against aldose reductase. Phytochemistry 2018, 152, 91–96. [Google Scholar] [CrossRef]
- Nukhimovskiy, E.L. Osnovy Biomorfologii Semennykh Rasteniy; Overlay: Moscow, Russia, 2002; Volume 2, p. 458. (In Russian) [Google Scholar]
- Nedosekina, T.V.; Agafonov, V.A. Morphogenesis and age stages of Scutellaria supina L. (Lamiaceae) in the conditions of middle-russia upland. Byull. MOIP. Otd. Biol. 2006, 111, 89–94. (In Russian) [Google Scholar]
- Cheryomushkina, V.A.; Guseva, A.A. Life forms of Scutellaria supina L. (Lamiaceae). Contemp. Probl. Ecol. 2015, 8, 624–635. [Google Scholar] [CrossRef]
- Cheryomushkina, V.A.; Guseva, A.A. Morhogenesis of Scutellaria grandiflora (Lamiaceae) and ontogenetic structure of its coenopopulations. Rastit. Resur. 2017, 53, 380–393. (In Russian) [Google Scholar]
- Guseva, A.A.; Cheryomushkina, V.A. Special development and condition of the Scutellaria sieversii (Lamiaceae) coenopopulations in Chu-Ilyisk mountains (Medium Asia). Plant Life Asian Russ. 2019, 3, 47–52. (In Russian) [Google Scholar]
- Cheryomushkina, V.A.; Astashenkov, A.Y. Scutellaria oreophila (Lamiaceae): Life form, shoot formation, ontogenesis. Bot. Zhurnal 2021, 106, 229–238. (In Russian) [Google Scholar]
- Olmstead, R. Phylogeny, Phenotypic Evolution, and Biogeography of the Scutellaria angustifolia Complex (Lamiaceae): Inference from Morphological and Molecular Data. Syst. Bot. 1989, 14, 320. [Google Scholar] [CrossRef]
- Olmstead, R.G. Biological and Historical Factors Influencing Genetic Diversity in the Scutellaria angustifolia complex (labiatae). Evolution 1990, 44, 54–70. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.-J.; Zhang, Z.-Y.; Hu, J.; Guo, L.-P.; Shao, A.-J.; Huang, L.-Q. Impacts of recent cultivation on genetic diversity pattern of a medicinal plant, Scutellaria baicalensis (Lamiaceae). BMC Genet. 2010, 11, 29. [Google Scholar] [CrossRef]
- Chiang, Y.-C.; Huang, B.-H.; Liao, P.-C. Diversification, Biogeographic Pattern, and Demographic History of Taiwanese Scutellaria Species Inferred from Nuclear and Chloroplast DNA. PLoS ONE 2012, 7, e50844. [Google Scholar] [CrossRef] [Green Version]
- Bai, C.; Wen, M.; Zhang, L.; Li, G. Genetic diversity and sampling strategy of Scutellaria baicalensis germplasm resources based on ISSR. Genet. Resour. Crop Evol. 2013, 60, 1673–1685. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome Fingerprinting by Simple Sequence Repeat (SSR)-Anchored Polymerase Chain Reaction Amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Nagaoka, T.; Ogihara, Y. Applicability of inter-simple sequence repeat polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD markers. Theor. Appl. Genet. 1997, 94, 597–602. [Google Scholar] [CrossRef]
- Geng, L.; Xu, J.; Ye, L.; Zhou, W.; Zhou, K. Connections with Nature and Environmental Behaviors. PLoS ONE 2015, 10, e0127247. [Google Scholar] [CrossRef] [PubMed]
- Zvyagina, N.S.; Dorogina, O.V.; A Krasnikov, A. Genetic differentiation and karyotype variation in Hedysarum chaiyrakanicum, an endemic species of Tuva Republic, Russia. Indian J. Exp. Biol. 2016, 54, 338–344. [Google Scholar] [PubMed]
- Bozchaloyi, S.E.; Sheidai, M. Molecular diversity and genetic relationships among Geranium pusillum and G. pyrenaicum with inter simple sequence repeat (ISSR) regions. Caryologia 2018, 71, 457–470. [Google Scholar] [CrossRef]
- dos Santos, J.O.; Mayo, S.J.; Bittencourt, C.B.; de Andrade, I.M. Genetic diversity in wild populations of the restinga ecotype of the cashew (Anacardium occidentale) in coastal Piauí, Brazil. Oesterreichische Bot. Z. 2019, 305, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Valuyskikh, O.E.; Shadrin, D.M. Population and molecular datasets for Gymnadenia conopsea (Orchidaceae). Data Brief 2019, 25, 104161. [Google Scholar] [CrossRef]
- Gatsuk, L.E.; Smirnova, O.V.; Vorontzova, L.I.; Zhukova, L.B.Z.A. Age States of Plants of Various Growth Forms: A Review. J. Ecol. 1980, 68, 675. [Google Scholar] [CrossRef]
- Smirnova, O.V.; Palenova, M.M.; Komarov, A.S. Ontogeny of Different Life Forms of Plants and Specific Features of Age and Spatial Structure of Their Populations. Russ. J. Dev. Biol. 2002, 33, 1–10. [Google Scholar] [CrossRef]
- Muraseva, D.S.; Guseva, A.A. ISSR primer screening for analysis of genetic diversity among Scutellaria tuvensis (Lamiaceae) populations. BIO Web Conf. 2021, 38, 00082. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Peakall, R.O.D.; Smouse, P.E. GENAlEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Yeh, F.; Yang, R.; Boyle, T. Popgene Version 1.32 Microsoft Windows-Based Freeware for Populations Genetic Analysis; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. Structure Harvester: A website and program for visualizing structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Artyukova, E.V.; Kholina, A.B.; Kozyrenko, M.M.; Zhuravlev, Y.N. Analysis of Genetic Variation in Rare Endemic Species Oxytropis chankaensis Jurtz. (Fabaceae) Using RAPD Markers. Russ. J. Genet. 2004, 40, 710–716. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J. Allozyme diversity in plant species. In Plant Population Genetics, Breeding and Germplasm Resources; Brown, A.H.D., Clegg, M.T., Kahler, A.L., Weir, B.S., Eds.; Sinauer: Sunderland, MA, USA, 1989; pp. 43–63. [Google Scholar]
- Boronnikova, S.V.; Kokaeva, Z.G.; Gostimsky, S.A.; Dribnokhodova, O.P.; Tikhomirova, N.N. Analysis of DNA polymorphism in a relict Uralian species, large-flowered foxglove (Digitalis grandiflora mill.), using RAPD and ISSR markers. Russ. J. Genet. 2007, 43, 530–535. [Google Scholar] [CrossRef]
- Selyutina, I.Y.; Konichenko, E.S.; Dorogina, O.V. Variability and interpopulation differentiation of the rare species Gueldenstaedtia monophylla Fisch. (Fabaceae). Vavilovskii Zhurnal Genet. Sel. 2017, 21, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Sözen, E.; Ozaydın, B. A study of genetic variation in endemic plant Centaurea wiedemanniana by using RAPD markers. Ekoloji 2010, 19, 1–8. [Google Scholar] [CrossRef]
- Mucciarelli, M.; Ferrazzini, D.; Belletti, P. Genetic Variability and Population Divergence in the Rare Fritillaria tubiformis subsp. moggridgei Rix (Liliaceae) as Revealed by RAPD Analysis. PLoS ONE 2014, 9, e101967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Shi, S.; Chang, E.; Yang, W.; Jiang, Z. Genetic Diversity of the Critically Endangered Thuja sutchuenensis Revealed by ISSR Markers and the Implications for Conservation. Int. J. Mol. Sci. 2013, 14, 14860–14871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gholami, S.; Vafaee, Y.; Nazari, F.; Ghorbani, A. Molecular characterization of endangered Iranian terrestrial orchids using ISSR markers and association with floral and tuber-related phenotypic traits. Physiol. Mol. Biol. Plants 2021, 27, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M. Gene Flow and the Geographic Structure of Natural Populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Trait | Habitat | ||||
|---|---|---|---|---|---|
| Beach Gravel | Detrital Cone | Steep Slopes with Coarse Rock Fragments | |||
| P 1 | P 2 | P 3 | P 4 | P 5 | |
| Primary-shoot length, cm | 1.9 ± 0.1 | 1.6 ± 0.1 | 5.1 ± 0.3 | 11.3 ± 0.4 | 10.9 ± 0.3 |
| Number of primary-shoot metameres | 10.4 ± 0.3 | 9.6 ± 0.3 | 7.9 ± 0.2 | 9.5 ± 0.3 | 12.1 ± 0.5 |
| Length of primary-shoot remainder, cm | 1.4 ± 0.1 | 1.1 ± 0.1 | 0.2 ± 0.01 | 8.6 ± 0.5 | 9.5 ± 0.3 |
| Number of metameres of primary-shoot remainder | 5.9 ± 0.4 | 8.5 ± 0.2 | 1.4 ± 0.1 | 7.3 ± 0.3 | 10.2 ± 0.6 |
| Number of generative shoots | 12.9 ± 0.6 | 13 ± 0.5 | 22.7 ± 2.0 | 56.2 ± 5.2 | 42.1 ± 3.8 |
| Length of generative shoots, cm | 6.5 ± 0.4 | 6 ± 0.2 | 6.9 ± 0.3 | 6.9 ± 0.6 | 11.1 ± 0.3 |
| Number of axes | 3.3 ± 0.5 | 4.1 ± 0.3 | 1.8 ± 0.1 | 6.7 ± 0.6 | 5.2 ± 0.4 |
| Length of axis, cm | 2.0 ± 0.2 | 2.0 ± 0.1 | 5.1 ± 0.5 | 8.2 ± 0.8 | 10.5 ± 0.5 |
| Age of axis, y | 3.3 ± 0.1 | 3.5 ± 0.1 | 8.4 ± 0.4 | 5 ± 0.3 | 4.1 ± 0.2 |
| Bush diameter, cm | 17.4 ± 0.9 | 18.4 ± 0.8 | 13.3 ± 1.1 | 22.6 ± 1.4 | 18.4 ± 0.9 |
| Primer | Sequence (5′→3′) | Band Size, bp | Number of Loci in Population | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P 1 | P 2 | P 3 | P 4 | P 5 | Total Study Population | |||||||||
| 1 * | 2 ** | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |||
| UBC 811 | (GA)8C | 450–1650 | 11 | 9 | 13 | 13 | 11 | 9 | 14 | 13 | 14 | 13 | 16 | 16 (100) |
| UBC 825 | (AC)8T | 300–1300 | 17 | 14 | 14 | 11 | 15 | 11 | 15 | 13 | 11 | 9 | 17 | 16 (94) |
| UBC 826 | (AC)8C | 470–2000 | 17 | 14 | 17 | 16 | 18 | 16 | 17 | 16 | 17 | 14 | 20 | 19 (95) |
| UBC 830 | (TG)8G | 250–1600 | 11 | 10 | 11 | 9 | 11 | 7 | 11 | 10 | 15 | 14 | 16 | 15 (94) |
| UBC 834 | (AG)8Y*T | 300–1400 | 13 | 9 | 13 | 9 | 12 | 7 | 13 | 11 | 15 | 15 | 15 | 15 (100) |
| UBC 836 | (AG)8YA | 300–1200 | 6 | 4 | 7 | 5 | 6 | 3 | 7 | 6 | 7 | 4 | 7 | 6 (86) |
| UBC 840 | (GA)8YT | 300–900 | 5 | 3 | 6 | 4 | 6 | 3 | 6 | 3 | 6 | 5 | 6 | 6 (100) |
| UBC 855 | (AC)8YT | 400–1500 | 13 | 8 | 13 | 9 | 15 | 11 | 12 | 7 | 15 | 10 | 15 | 13 (87) |
| UBC 856 | (AC)8YA | 450–1700 | 14 | 8 | 13 | 11 | 13 | 9 | 13 | 9 | 12 | 6 | 14 | 13 (93) |
| UBC 857 | (AC)8YG | 350–1650 | 14 | 10 | 12 | 9 | 14 | 9 | 14 | 13 | 13 | 13 | 15 | 15 (100) |
| Total | 121 | 90 (74) | 119 | 96 (81) | 121 | 85 (70) | 121 | 102 (84) | 125 | 112 (90) | 141 | 134 (95) | ||
| Population | HE | na | ne |
|---|---|---|---|
| P 1 | 0.202 | 1.468 | 1.342 |
| P 2 | 0.213 | 1.518 | 1.354 |
| P 3 | 0.189 | 1.461 | 1.321 |
| P 4 | 0.236 | 1.582 | 1.407 |
| P 5 | 0.266 | 1.638 | 1.459 |
| Population | P 1 | P 2 | P 3 | P 4 | P 5 |
|---|---|---|---|---|---|
| P 1 | 2 | 115 | 110 | 154 | |
| P 2 | 0.071 | 117 | 110 | 154 | |
| P 3 | 0.049 | 0.098 | 137 | 153 | |
| P 4 | 0.115 | 0.099 | 0.128 | 48 | |
| P 5 | 0.163 | 0.136 | 0.173 | 0.102 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guseva, A.; Muraseva, D.; Cheryomushkina, V. Morphological and Genetic Diversity of Scutellaria tuvensis Juz., an Endemic of Desert Steppes. Int. J. Plant Biol. 2022, 13, 473-484. https://doi.org/10.3390/ijpb13040038
Guseva A, Muraseva D, Cheryomushkina V. Morphological and Genetic Diversity of Scutellaria tuvensis Juz., an Endemic of Desert Steppes. International Journal of Plant Biology. 2022; 13(4):473-484. https://doi.org/10.3390/ijpb13040038
Chicago/Turabian StyleGuseva, Alexandra, Dinara Muraseva, and Vera Cheryomushkina. 2022. "Morphological and Genetic Diversity of Scutellaria tuvensis Juz., an Endemic of Desert Steppes" International Journal of Plant Biology 13, no. 4: 473-484. https://doi.org/10.3390/ijpb13040038