
Cryptococcus gattii Can Use the Cactus Pilosocereus spp. to Grow and Develop a Capsule and Produce Melanin In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cactus Media
2.3. Growth Curve
2.4. Microscopy and Capsule Measurements
2.5. Inoculation of Cryptococcus gattii into Pilosocereus spp.
2.6. Melanin Ghosts
3. Results
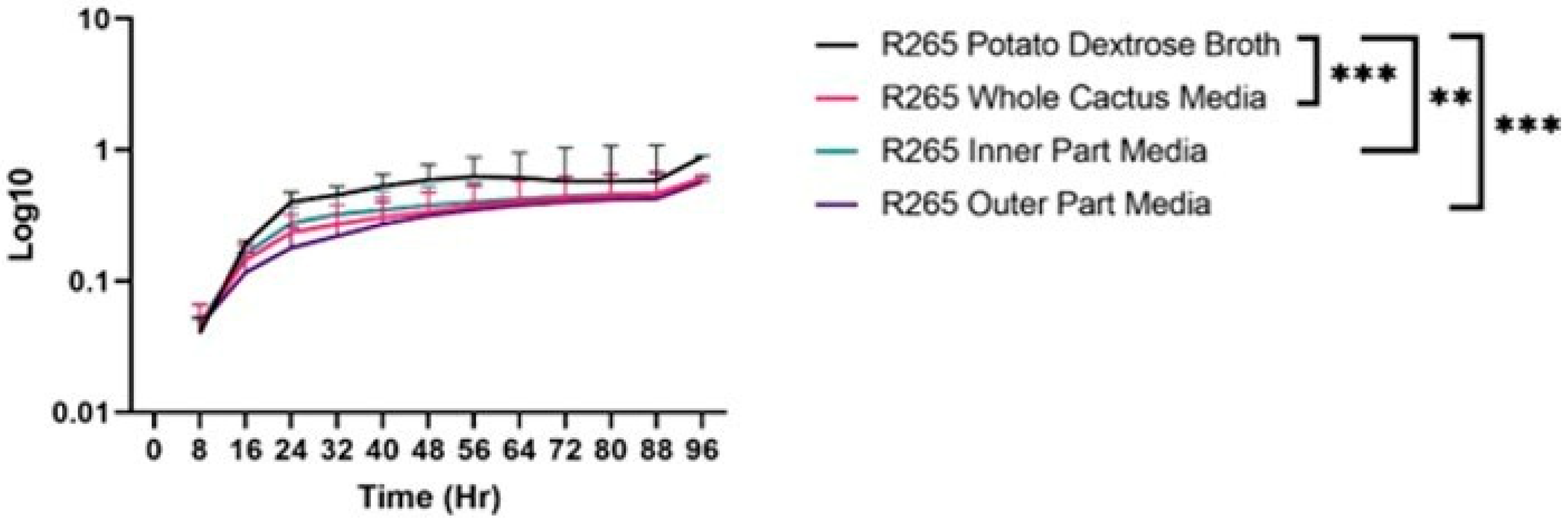
3.1. Growth Differences of C. gattii in Cactus Media/Broth
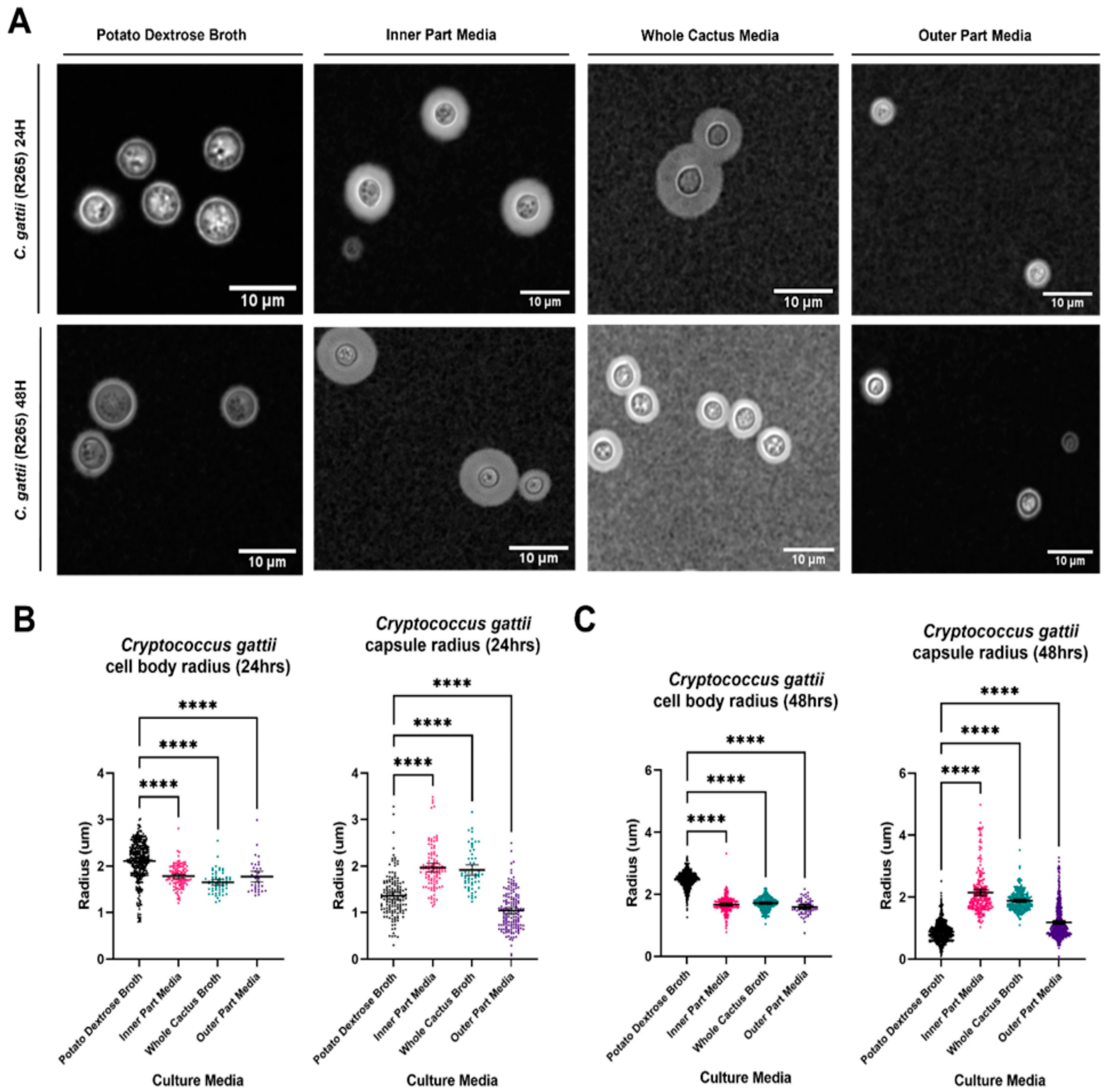
3.2. Analysis of Cell Body and Capsule Radius of C. gattii in Cactus Media
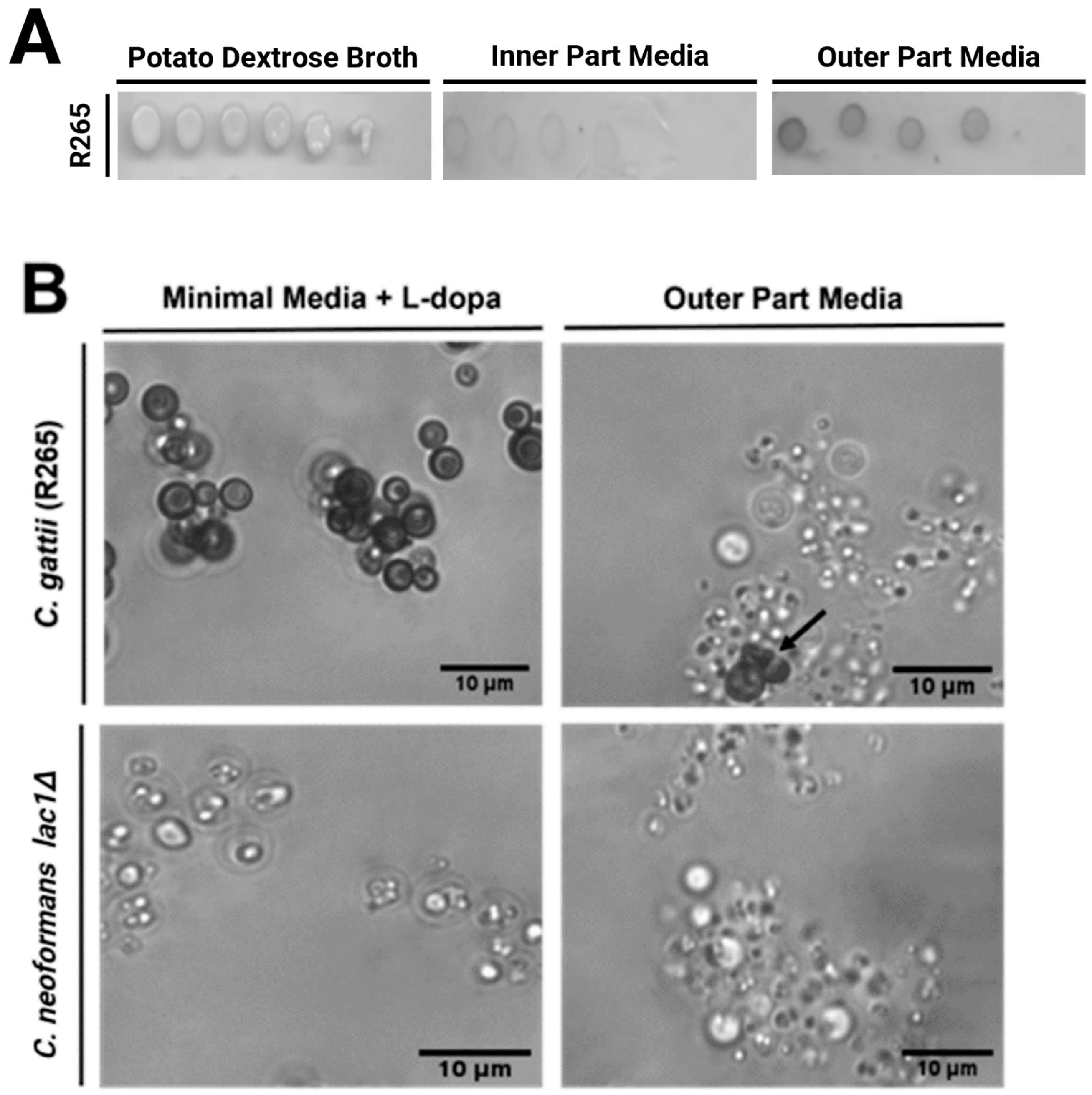
3.3. Pigment Produced by C. gattii When Grown in Outer Part Broth Is Laccase-Dependent
3.4. Melanin Is Produce in Cactus When C. gattii Is Directly Inoculated
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, T.G.; Perfect, J.R. Cryptococcosis in the era of AIDS--100 years after the discovery of Cryptococcus neoformans. Clin. Microbiol. Rev. 1995, 8, 515–548. [Google Scholar] [CrossRef]
- Doering, T.L.; Nosanchuk, J.D.; Roberts, W.K.; Casadevall, A. Melanin as a potential cryptococcal defence against microbicidal proteins. Med. Mycol. 1999, 37, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Doering, T.L. How sweet it is! Cell wall biogenesis and polysaccharide capsule formation in Cryptococcus neoformans. Annu. Rev. Microbiol. 2009, 63, 223–247. [Google Scholar] [CrossRef] [Green Version]
- O’Meara, T.R.; Alspaugh, J.A. The Cryptococcus neoformans capsule: A sword and a shield. Clin. Microbiol. Rev. 2012, 25, 387–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.A.; Janbon, G.; Idnurm, A.; Bahn, Y.S. Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harb. Perspect. Med. 2014, 4, a019760. [Google Scholar] [CrossRef] [Green Version]
- Escandón, P.; Sánchez, A.; Martínez, M.; Meyer, W.; Castañeda, E. Molecular epidemiology of clinical and environmental isolates of the Cryptococcus neoformans species complex reveals a high genetic diversity and the presence of the molecular type VGII mating type a in Colombia. FEMS Yeast Res. 2006, 6, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Kidd, S.E.; Hagen, F.; Tscharke, R.L.; Huynh, M.; Bartlett, K.H.; Fyfe, M.; MacDougall, L.; Boekhout, T.; Kwon-Chung, K.J.; Meyer, W. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Columbia, Canada). Proc. Natl. Acad. Sci. USA 2004, 101, 17258–17263. [Google Scholar] [CrossRef] [Green Version]
- Byrnes, E.J., 3rd; Li, W.; Lewit, Y.; Ma, H.; Voelz, K.; Ren, P.; Carter, D.A.; Chaturvedi, V.; Bildfell, R.J.; May, R.C.; et al. Emergence and pathogenicity of highly virulent Cryptococcus gattii genotypes in the northwest United States. PLoS Pathog. 2010, 6, e1000850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrnes, E.J., 3rd; Li, W.; Ren, P.; Lewit, Y.; Voelz, K.; Fraser, J.A.; Dietrich, F.S.; May, R.C.; Chaturvedi, S.; Chaturvedi, V.; et al. A diverse population of Cryptococcus gattii molecular type VGIII in southern Californian HIV/AIDS patients. PLoS Pathog. 2011, 7, e1002205. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Diaz, J.H. The Disease Ecology, Epidemiology, Clinical Manifestations, and Management of Emerging Cryptococcus gattii Complex Infections. Wilderness Environ. Med. 2020, 31, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogliati, M. Global warming impact on the expansion of fundamental niche of Cryptococcus gattii VGI in Europe. Environ. Microbiol. Rep. 2021, 13, 375–383. [Google Scholar] [CrossRef]
- Sánchez Paredes, E.; Aguilera León, Y.P.; Alvarado-Ramírez, E.V.; Castañón Olivares, L.R.; Martínez Aguilar, M.L. Climate Change and Global Distribution of Cryptococcosis. In The Impact of Climate Change on Fungal Diseases; Frías-De-León, M.G., Brunner-Mendoza, C., Reyes-Montes, M.d.R., Duarte-Escalante, E., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 181–201. [Google Scholar]
- Hagen, F.; Ceresini, P.C.; Polacheck, I.; Ma, H.; van Nieuwerburgh, F.; Gabaldón, T.; Kagan, S.; Pursall, E.R.; Hoogveld, H.L.; van Iersel, L.J.J.; et al. Ancient Dispersal of the Human Fungal Pathogen Cryptococcus gattii from the Amazon Rainforest. PloS ONE 2013, 8, e71148. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Randhawa, H.S.; Prakash, A.; Meis, J.F. Environmental prevalence of Cryptococcus neoformans and Cryptococcus gattii in India: An update. Crit. Rev. Microbiol. 2012, 38, 1–16. [Google Scholar] [CrossRef]
- Springer, D.J.; Chaturvedi, V. Projecting global occurrence of Cryptococcus gattii. Emerg. Infect. Dis. 2010, 16, 14–20. [Google Scholar] [CrossRef]
- Mak, S.; Vélez, N.; Castañeda, E.; Escandón, P. The Fungus among Us: Cryptococcus neoformans and Cryptococcus gattii Ecological Modeling for Colombia. J. Fungi 2015, 1, 332–344. [Google Scholar] [CrossRef] [Green Version]
- Kidd, S.E.; Chow, Y.; Mak, S.; Bach, P.J.; Chen, H.; Hingston, A.O.; Kronstad, J.W.; Bartlett, K.H. Characterization of environmental sources of the human and animal pathogen Cryptococcus gattii in British Columbia, Canada, and the Pacific Northwest of the United States. Appl. Environ. Microbiol. 2007, 73, 1433–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrnes, E.J., 3rd; Bildfell, R.J.; Frank, S.A.; Mitchell, T.G.; Marr, K.A.; Heitman, J. Molecular evidence that the range of the Vancouver Island outbreak of Cryptococcus gattii infection has expanded into the Pacific Northwest in the United States. J. Infect. Dis. 2009, 199, 1081–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, L.M.N.; Maguire, J.A.; Doyle, P.; Fyfe, M.; Roscoe, D.L. Cryptococcus neoformans infections at Vancouver Hospital and Health Sciences Centre (1997–2002): Epidemiology, microbiology and histopathology. J. Med. Microbiol. 2004, 53 Pt 9, 935–940. [Google Scholar] [CrossRef] [Green Version]
- Mak, S.; Klinkenberg, B.; Bartlett, K.; Fyfe, M. Ecological niche modeling of Cryptococcus gattii in British Columbia, Canada. Environ. Health Perspect. 2010, 118, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Springer, D.J.; Billmyre, R.B.; Filler, E.E.; Voelz, K.; Pursall, R.; Mieczkowski, P.A.; Larsen, R.A.; Dietrich, F.S.; May, R.C.; Filler, S.G.; et al. Cryptococcus gattii VGIII isolates causing infections in HIV/AIDS patients in Southern California: Identification of the local environmental source as arboreal. PLoS Pathog. 2014, 10, e1004285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Randhawa, H.S.; Boekhout, T.; Hagen, F.; Klaassen, C.H.; Meis, J.F. Temperate climate niche for Cryptococcus gattii in Northern Europe. Emerg. Infect. Dis. 2012, 18, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, M.; D’Amicis, R.; Zani, A.; Montagna, M.T.; Caggiano, G.; De Giglio, O.; Balbino, S.; De Donno, A.; Serio, F.; Susever, S.; et al. Environmental distribution of Cryptococcus neoformans and C. gattii around the Mediterranean basin. FEMS Yeast Res. 2016, 16, fow045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loperena-Alvarez, Y.; Ren, P.; Li, X.; Bopp, D.J.; Ruiz, A.; Chaturvedi, V.; Rios-Velazquez, C. Genotypic characterization of environmental isolates of Cryptococcus gattii from Puerto Rico. Mycopathologia 2010, 170, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.S.; Milhausen, S.M.; Manthey, M.K. 3-Hydroxyanthranilate in Cryptococcus neoformans: A secreted reductant that does not enable wood rot. Med. Mycol. 2003, 41, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Idnurm, A.; Giles, S.S.; Perfect, J.R.; Heitman, J. Peroxisome function regulates growth on glucose in the basidiomycete fungus Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Aisen, P.; Casadevall, A. Melanin, melanin “ghosts,” and melanin composition in Cryptococcus neoformans. Infect. Immun. 1996, 64, 2420–2424. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Prados-Rosales, R.; Itin, B.; Casadevall, A.; Stark, R.E. Solid-state NMR Reveals the Carbon-based Molecular Architecture of Cryptococcus neoformans Fungal Eumelanins in the Cell Wall. J. Biol. Chem. 2015, 290, 13779–13790. [Google Scholar] [CrossRef] [Green Version]
- Williamson, P.R.; Wakamatsu, K.; Ito, S. Melanin biosynthesis in Cryptococcus neoformans. J. Bacteriol. 1998, 180, 1570–1572. [Google Scholar] [CrossRef] [Green Version]
- Schwager, H.; Masselter, T.; Speck, T.; Neinhuis, C. Functional morphology and biomechanics of branch-stem junctions in columnar cacti. Proc. Biol. Sci. 2013, 280, 20132244. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, O.; Casadevall, A. Experimental modulation of capsule size in Cryptococcus neoformans. Biol. Proced. Online 2004, 6, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Littman, M.L. Capsule synthesis by Cryptococcus neoformans. Trans. N. Y. Acad. Sci. 1958, 20, 623–648. [Google Scholar] [CrossRef]
- Trevijano-Contador, N.; Rossi, S.A.; Alves, E.; Landín-Ferreiroa, S.; Zaragoza, O. Capsule Enlargement in Cryptococcus neoformans Is Dependent on Mitochondrial Activity. Front. Microbiol. 2017, 8, 1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Rocha Filho, C.A.A.; Karla Amorim, P.; de Albuquerque Lima, T.; da Silva, P.M.; Celine de Moura, M.; Barroso Coelho, L.C.B.; Benedeta Zingali, R.; Viana Pontual, E.; Henrique Napoleão, T.; Guedes Paiva, P.M. PgTI, the First Bioactive Protein Isolated from the Cactus Pilosocereus gounellei, is a Trypsin Inhibitor with Antimicrobial Activity. Adv. Res. 2019, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Casadevall, A. Impact of melanin on microbial virulence and clinical resistance to antimicrobial compounds. Antimicrob. Agents Chemother. 2006, 50, 3519–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mednick, A.J.; Nosanchuk, J.D.; Casadevall, A. Melanization of Cryptococcus neoformans affects lung inflammatory responses during cryptococcal infection. Infect. Immun. 2005, 73, 2012–2019. [Google Scholar] [CrossRef] [Green Version]
- Chrissian, C.; Camacho, E.; Fu, M.S.; Prados-Rosales, R.; Chatterjee, S.; Cordero, R.J.B.; Lodge, J.K.; Casadevall, A.; Stark, R.E. Melanin deposition in two Cryptococcus species depends on cell-wall composition and flexibility. J. Biol. Chem. 2020, 295, 1815–1828. [Google Scholar] [CrossRef] [PubMed]
- Ngamskulrungroj, P.; Price, J.; Sorrell, T.; Perfect, J.R.; Meyer, W. Cryptococcus gattii virulence composite: Candidate genes revealed by microarray analysis of high and less virulent Vancouver island outbreak strains. PLoS ONE 2011, 6, e16076. [Google Scholar] [CrossRef] [Green Version]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [Green Version]
- da Silveira Agostini-Costa, T. Bioactive compounds and health benefits of Pereskioideae and Cactoideae: A review. Food Chem. 2020, 327, 126961. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Irizarry, P.; Sánchez, B.; Loperena-Álvarez, Y. Cryptococcus gattii Can Use the Cactus Pilosocereus spp. to Grow and Develop a Capsule and Produce Melanin In Vitro. Microbiol. Res. 2023, 14, 798-809. https://doi.org/10.3390/microbiolres14020056
Ramos-Irizarry P, Sánchez B, Loperena-Álvarez Y. Cryptococcus gattii Can Use the Cactus Pilosocereus spp. to Grow and Develop a Capsule and Produce Melanin In Vitro. Microbiology Research. 2023; 14(2):798-809. https://doi.org/10.3390/microbiolres14020056
Chicago/Turabian StyleRamos-Irizarry, Paola, Bárbara Sánchez, and Yaliz Loperena-Álvarez. 2023. "Cryptococcus gattii Can Use the Cactus Pilosocereus spp. to Grow and Develop a Capsule and Produce Melanin In Vitro" Microbiology Research 14, no. 2: 798-809. https://doi.org/10.3390/microbiolres14020056